










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.4 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i4.16
ARTICLE
"Male effect" and "temporary weaning" in synchronization of post-partum ovarian activity in Pelibuey ewes
J.A. Hernández-MarínI, V; C. Cortez-RomeroII; C.A. Herrera-CorredorIII; P. Pérez-HernándezIV; A. Pro-MartínezV; J. Gallegos-SánchezV, #
IDepartamento de Veterinaria y Zootecnia, División de Ciencias de la Vida, Campus Irapuato-Salamanca, Universidad de Guanajuato, Irapuato, Guanajuato, México, C.P. 36824
IICampus San Luis Potosí, Colegio de Postgraduados, Salinas de Hidalgo, San Luis Potosí, México, C.P. 78622
IIIFacultad de Agronomía, Universidad Autónoma de San Luis Potosí, Soledad de Graciano Sánchez, San Luis Potosí, México, Apartado Postal 32
IVCampus Veracruz, Colegio de Postgraduados, Carretera Xalapa-Veracruz km 88.5, Veracruz, México. C.P. 91690
VGanadería, Campus Montecillo, Colegio de Postgraduados, Montecillo, Estado de México, México, C.P. 56230
ABSTRACT
To evaluate the response of the "male effect" and "temporary weaning" on the synchronization of postpartum ovarian activity in Pelibuey ewes, an experiment was carried out using 78 Pelibuey ewes with suckling lambs. The ewes were separated from their offspring for 48 hours and randomly assigned to one of four treatments derived from the arrangement of two factors, "male effect" and "temporary weaning", each at two levels. Treatments were: T1 (n = 20) control ewes, without "male effect" and without "temporary weaning"); T2 (n = 19) ewes without "male effect" and with "temporary weaning"; T3 (n = 20) ewes with "male effect" and without "temporary weaning"; and T4 (n = 19), ewes with "male effect" and with "temporary weaning". The response to oestrus, return to oestrus, gestation rate and lambing rate were analysed using logistic regression. The onset of oestrus was analysed using survival curves. No significant differences were found for lambing rate and prolificacy among treatments. "Temporary weaning" (T2) and "male effect" (T3) did not influence the response to oestrus, rate and duration of return to oestrus, or gestation rate and fertility, and was similar to the control group (T1). The interaction of "male effect" with "temporary weaning" (T4) increased the response to oestrus, reduced the rate and duration of return to oestrus, and the gestation rate, but increased fertility. Synchronizing post-partum ovarian activity with "male effect" and "temporary weaning" reduces the onset of oestrus and the rate of return to oestrus, but increases the response to oestrus and fecundity in Pelibuey ewes.
Keywords: Biostimulation, progestogens, prolificacy, prostaglandins, suckling
Introduction
Currently, reproductive management protocols are based on the application of exogenous hormones such as progestogens (P4), which simulate the action of the corpus luteum (CL) and prostaglandins (PGF2a), which lyse CL to induce a follicular phase followed by ovulation (Abecia et al., 2012). Socio-sexual methods, such as the use of the "male effect" also produce physiological responses that increase follicular development, stimulate the secretion of estradiol (Meilán & Ungerfeld, 2014), increase the secretion of luteinizing hormone (LH) and induce ovulation (Hernández-Marín et al., 2016). To reduce the onset of oestrus without affecting lamb development, "temporary weaning" (Rodríguez et al., 1986) can be applied, or vegetable fats can be included in supplements to feed sheep, which improve the energy balance and bioavailability of fatty acids for the synthesis of cholesterol, eicosanoids and steroid hormones (Torres-Zapata et al., 2016).
The simplest artificial method for inducing oestral behaviour and ovulation in sheep is to use a controlled internal drug release (CIDR®) device, because it imitates the presence of the CL in a natural oestrus cycle, alters the release of LH, and maintains the quality of ovulation, animal welfare and public health. In hair sheep synchronized with CIDR, control of suckling for short periods of the day (30 min) and exposure to male sheep favoured the increase and concentration in the number of ovulations (Castillo-Maldonado et al., 2013) and conception before 35 days post-partum (Oldham et al., 2011).
Reproductive management with PGF2a is a good alternative, because this hormone is metabolized faster in the liver and does not accumulate in tissues (Davis et al., 1980). Its administration in two doses is recommended to synchronize oestrus in cycling sheep, and its response varies according to the technique of insemination, the dose and the intervals between doses (Fierro et al., 2013). It is necessary to generate protocols of short duration, with fewer doses and more effective release methods (Abecia et al., 2011) combined with natural methods to provide for healthier and safer animal production. It is unknown whether a "male effect" or "temporary weaning" is sufficient to replace the second dose of PGF2a during the luteal phase in sheep (Ungerfeld, 2011). Therefore, the objective of the present study was to evaluate the "male effect" and "temporary weaning" and their effects on the synchronization of ovarian activity at 45 days post-partum with two doses of PGF2a in Pelibuey ewes.
Materials and Methods
The study was conducted in the Laboratorio de Reproducción de Ovinos y Caprinos (LaROCa) (Laboratory of Ovine and Caprine Reproduction) at Colegio de Postgraduados, Campus Montecillo, Texcoco, State of Mexico, Mexico (located at 19°29' N, 98°53' W). The facility is 2250 metres above sea level, with a climate classified as Cb(wo)(w)(i')g. This corresponds to a temperate sub-humid climate with a summer rainfall and an average annual temperature of 636.5 mm and 15.2 °C, respectively (García, 2004).
During the study, 78 Pelibuey ewes were used, and had an average body of 56.6 ± 6.8 kg. Each ewe consumed 2.5 kg/day of a diet consisting of milled oat hay (700 g/kg) mixed with a commercial concentrate (300 g/kg) (Borrega Plus®, Alimentos Tepexpan, México) with 15% crude protein and 2.9 Mcal metabolizable energy/kg, mineral salts and free access to water. This study was carried out in accordance with the standards for use and care of research animals at Colegio de Postgraduados, Campus Montecillo, Mexico, and according to Normas Oficiales Mexicana (Mexican Standards) NOM-024-ZOO-1995 and NOM-033-ZOO-1995 (SAGARPA, 2015).
Seven weeks before the biostimulation of the ewes, they were kept at a minimum distance of 500 m from the pen containing rams (Walkden-Brown et al., 1993) to avoid visual, auditory and olfactory contact between them. This method was designed to increase the stimulus at the time that the "male effect" was realized. Five adult unneutered and sexually experienced Pelibuey males were used to provide the "male effect". The rams were trained for collection of semen with an artificial vagina, on which macro/micro seminal tests were performed. A ram fit with an apron was introduced to the ewe pens for two periods of 30 min/day (08:00 and 15:00) from day 30 to day 45 post-partum. During the "male effect" the exposed ram was replaced by a different one to prevent the females from getting used to a particular male. The rams were used for biostimulation and oestrus detection in ewes.
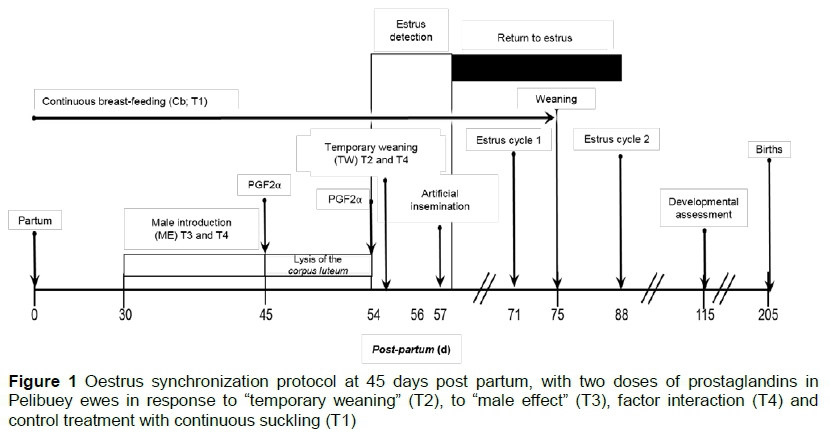
The Pelibuey ewes were assigned to treatments, following a completely randomized design in a 2 x 2 factorial arrangement. The combination of factor levels generated four treatments: T1 (n = 20, control sheep, without stimulus from the "male effect" and without "temporary weaning" after the second dose of PGF2a; 54 days post-partum); T2 (n = 20, sheep without stimulus from the "male effect" and with "temporary weaning" for 48 hours after the second dose of PGF2a (54 days post-partum); T3 (n = 19, sheep stimulated daily by the "male effect" for 15 days and without "temporary weaning" after the second dose of PGF2a, 54 days post-partum); and T4 (n = 19; sheep stimulated daily by the "male effect" for 15 days and with "temporary weaning" for 48 hours after the second dose of PGF2a, 54 days post-partum) (Figure 1). PGF2a was applied when a sheep ovulated, thus lysing the CL.
All ewes were given two applications of 1 mL synthetic prostaglandin (250 mg sodium cloprostenol, Celosil®, MSD Animal Health, Mexico) intramuscularly at 45 and 54 days post-partum. After the second dose, they were exposed for 30 min to a Pelibuey ram that was provided with an apron (preventing copulation), to detect oestrus every 4 hours to 72 hours. Ewes in oestrus were moved to a different pen to perform artificial insemination. All Pelibuey ewes in the present study suckling their lambs continuously from lambing until weaning (75 days). At 54 days post-partum, the lambs were temporarily weaned, when ewes and their lambs were separated and housed in different pens for 48 hours to induce a pre-ovulatory peak of LH (Rodríguez et al., 1986).
The synchronized ewes were inseminated using intrauterine laparoscopy and fresh semen (Cortez-Romero et al., 2011). They were kept without solid or liquid diets for 24 hours prior to artificial insemination to avoid accidental puncturing of the rumen or urinary bladder. Two systematized swivelling stretchers were used to hold and immobilize the ewes in a dorsal decubitus position and tilt them at an angle of 45°, so that the viscera could move in a cranial direction (McKelvey et al., 1985). The abdominal region anterior to the udder was then shaved and disinfected with an iodine solution.
The number of ewes that returned to oestrus was detected by introducing an adult Pelibuey ram with an apron for 60 min/day (09:00 and 16:00) for 34 days after insemination (approximate duration of two normal oestrus cycles). The number of pregnant ewes was recorded using an abdominal transducer of 7.5 MHz integrated with real-time ultrasonography equipment (Medison SonoAce Pico, USA) at 60 days after artificial insemination.
Statistical Analysis Systems® software (SAS, 2012) was used to analyse the data. The statistical model was:
Yijk = μ + A, + Bj + (AB)ij + Eijk
where: Yijk = variable response in repetition k, level j of B, level i of A
μ = general average
Ai = effect of factor A at level i
Bj = effect of factor B at level j
(AB)ij = effect of the AB interaction at level i, j
Eijk= random error
The response variables for oestrus, return to oestrus, gestation rate and lambing rate were analysed with a logistic regression model using the PROC LOGISTIC procedure, and evaluated with the Wald test. For the initiation of oestrus, the Shapiro & Wilk (1965) test was performed to observe univariate normality and justify the analysis with the Kaplan Meier survival curves and the Cox regression method (Salinas, 2008), and the log-rank test was used, using the LIFETEST procedure. Analyses of variance of prolificacy and fecundity were performed with a completely randomized design using a 2 x 2 factorial arrangement.
Results
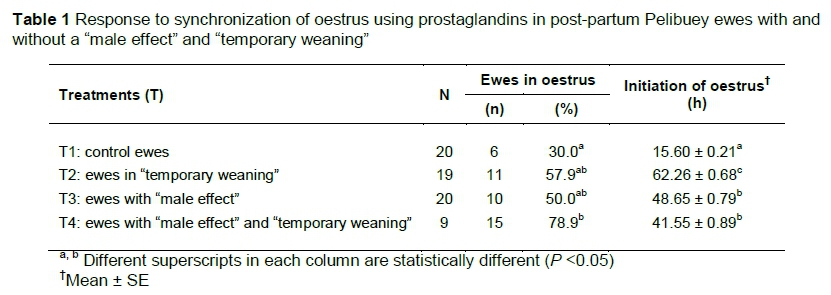
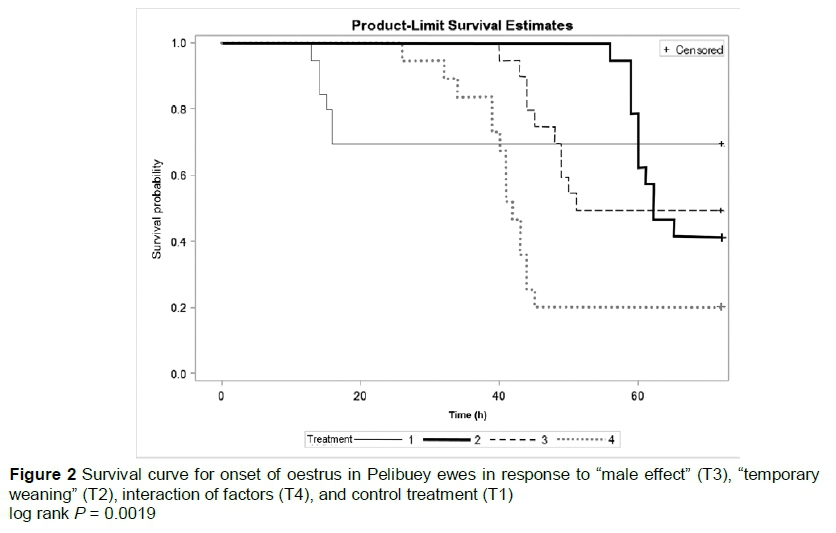
The "male effect" and its interaction with "temporary weaning" influenced the response to oestrus significantly (P <0.05) with two doses of prostaglandins in post-partum Pelibuey ewes (Table 1). The onset of oestrus differed (P <0.05) among treatments (Figure 2). "Temporary weaning" did not favour (P <0.05) the onset of oestrus, but this was improved (P <0.05) by interaction of the "male effect" with "temporary weaning" (Table 1).
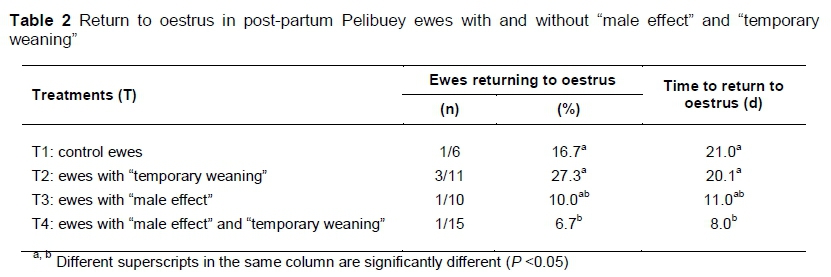
The "male effect" and "temporary weaning" did not influence (P >0.05) the percentage return to oestrus, and the effect was similar (p >0.05) to that observed for the control ewes. Ewes that were subjected to a "male effect" and "temporary weaning" showed a reduced (P <0.05) percentage returning to oestrus and duration of "return to oestrus" (Table 2).
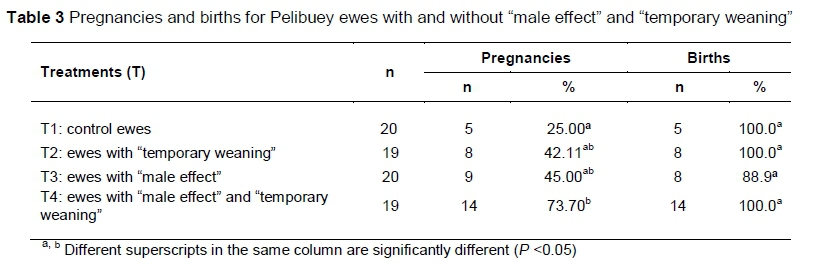
The interaction of the male effect with "temporary weaning" significantly influenced (P <0.05) gestation rate. Yet, lambing rates were similar (P >0.05) in post-partum Pelibuey ewes in response to the synchronization protocol with PGF2a (Table 3).
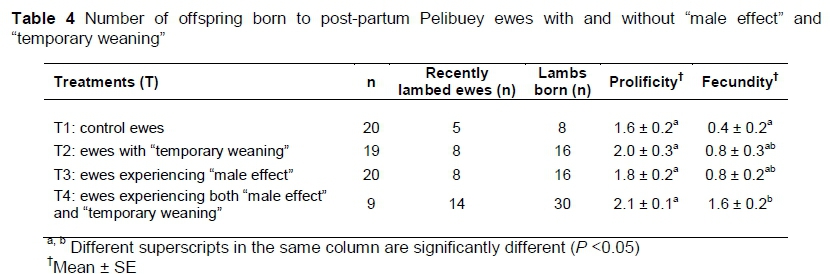
The "male effect" and "temporary weaning", and their interaction, did not significantly influence (P >0.05) the prolificacy of post-partum Pelibuey ewes. This effect was similar (P >0.05) to that observed in the control ewes (Table 4). The "male effect" and "temporary weaning" did not significantly increase (P >0.05) the fecundity of post-partum Pelibuey ewes. This result was statistically similar (P >0.05) to that observed for the control sheep. However, ewes that experienced the interaction of the "male effect" and "temporary weaning" significantly increased (P <0.05) their fecundity (Table 4).
Discussion
In the induction of luteolysis, the CL is eliminated and the follicular phase is induced with ovulation (Abecia et al., 2012). The results obtained in response to oestrus with the two applications of PGF2a, with intervals of 10 days between doses were similar by 84% and 88% to results obtained by Das et al. (1999), but differed by 42.6% for oestrus as reported by Olivera-Muzante et al. (2013). The presentation of oestrus in Pelibuey ewes in T4 in the present study is attributed to the difference between doses and application intervals of PGF2a, because the reduction of double doses with 9 to 10 days of difference between applications of some PGF2a analogs can induce luteolysis and reduce costs per treatment (Contreras-Solís et al., 2009). Camacho-Ronquillo et al. (2008) reported 35.7% oestrus in Pelibuey ewes without restriction of suckling and synchronized with PGF2a at 30 and 40 days post-partum, which is similar to the results in the present study from ewes in T1, but differs by 43.2% from the response to oestrus by ewes in T4.
Separating the lambs for 72 hours increased ovulation, reduced the onset of oestrus in ewes, and did not affect the growth or development of the offspring (Rodríguez et al., 1986). Additionally, the proportion of ewes in oestrus increased when rams were introduced at the time of applying the second dose of PGF2a (Ungerfeld, 2011). Therefore, the interaction of the male effect (before applying the first dose of PGF2a) with temporary weaning (after the second dose of PGF2a) was reflected in the response to oestrus synchronization in Pelibuey ewes in T4 in the present study.
The presence of lambs with suckling ewes induces oestrus during the post-partum period (Arteaga et al., 2007). This result differs, depending on the synchronization protocol and physiological state of the ewes. In protocols using PGF2a, oestrus was reported to occur at 36 ± 2.3 hours (Letelier et al., 2011) and between 38.6 ± 0.5 and 51.6 ± 2.4 hours (Fierro et al., 2013). These results are higher than those obtained for ewes in T1, and different from those for ewes in T2, T3 and T4 in the present study. The introduction of the male influences the onset of oestrus, following the drug applied to synchronize oestrus. This was similar to introducing vasectomized rams after applying the second dose of PGF2a to advance the onset of oestrus (Ungerfeld, 2011.
In the present study, T4 influenced the percentage of ewes returning to oestrus in synchrony with PGF2a, and consequently the rate of return to oestrus was reduced. This result may be because of interaction. Yet, the ewes returned to oestrus earlier, possibly as a consequence of reduced progesterone (P4), because the oviduct and uterine environment were not adequate for maintaining embryonic development (Olivera-Muzante et al., 2013). This is attributed to the fact that during synchronization of oestrus, ewes are sensitive to environmental factors that influence the reproductive processes, such as thermal stress (Naqvi et al., 2004), in which low concentrations of P4 are related to delays in embryonic development, resulting in conception failure or changes in weight during the post-partum period (Castillo-Maldonado et al., 2013), in which ewes with continuous suckling use their body reserves of energy, protein and minerals for milk production. Thus, it is necessary to include vegetable fats as energy supplements and ensure the bioavailability of fatty acids for the synthesis of cholesterol, eicosanoids and steroid hormones (Torres-Zapata et al., 2016).
The differences in the concentration of P4 are attributed to the lack of atresia of dominant follicles, thus reducing the formation of a normal CL, or a CL of short duration is generated. The lack of P4 is associated with the syndrome of corpus luteum of short duration in which ovulation is induced during anoestrus in ewes (Rodríguez-Iglesias et al., 2013), which is owing to anticipated luteolysis, which is activated when the CL is sensitive to PGF2a. In sheep, this occurs three to four days after ovulation (Rubianes et al., 2003).
In synchronization protocols with PGF2a, Olivera-Muzante et al. (2013) reported 84.7% and 65.3% return to oestrus, which was reflected as lower pregnancy rates. The insemination technique influences the percentage of pregnancy, such as 62% of pregnant sheep synchronized with two doses of PGF2a and inseminated by laparoscopy (Fierro et al., 2011), which is greater than 42.6% of pregnant sheep inseminated at a fixed time (Olivera-Muzante et al., 2013). These results differ from the percentage of pregnancies obtained with synchronization using PGF2a in the present study. The proportion of pregnant sheep that responded to synchronization and did not return to oestrus was reflected in fertilization. Values less than 45% of gestation synchronized with two doses of PGF2a in the present stud. Perhaps these were the result of some failure in ovulation, lack of fertilization, the development of an abnormal CL that could not maintain embryonic development or by embryonic death before or after maternal recognition of pregnancy (Fierro et al., 2013). The application of the second dose of PGF2a during the luteal middle phase causes an asynchrony between follicular waves that determine the presence of follicles in different stages (growth, permanence or regression of follicles) and promotes the long life of the ovulatory follicle, or ovulation of newly emerged follicles (Barret et al., 2002). Therefore, it is normal to obtain a low reproductive response after insemination at a fixed time with PGF2a, because the proportion of ovulatory follicles from the second follicular wave results in less synchronized ovulation, compared with the reduction of the probability of conception when inseminating 48 hours after the second dose of PGF2a (Olivera-Muzante et al., 2011).
It is possible that ewes do not maintain gestation after foetal death owing to endocrine failure during the establishment of the foetus, or that the amount of P4 is not enough to maintain it (Bari et al., 2003). This effect was observed in Pelibuey sheep in T1, T2 and T3 in the present study, which is related to the management factors associated with artificial insemination that modify fertility. In reproductive planning, the intervals between births, reproductive season, age of the mother, heat stress and nutritional status or breed are negative influences. Dixon et al. (2007) reported a 19.9% loss of late embryos, and estimated that 21.2% were lost from day 25 after trans-cervical post-insemination with frozen semen. Therefore, the insemination protocol influenced the response of the Pelibuey ewes to synchronization with two doses of PGF2a in the present study owing to the number of services and the time of insemination, because they differed between artificial insemination and natural breeding (Kumar & Naqvi, 2014).
The three main variables that contribute to prolificacy are ovulatory rate, embryonic and foetal survival (Martin et al., 2004). The GnRH analogs in synchronization with PGF2a increase prolificacy in ewes (1.7 offspring) (Martemucci & D'Alessandro, 2011) or do not affect it, but the fertility increases when applying the GnRH 24 hours after the second dose of PGF2a. Castillo-Maldonado et al. (2013) reported 1.8 ± 0.2 offspring from prolificacy and 1.42 offspring from fecundity, which differs from the results obtained in the Pelibuey ewes synchronized with two doses of PGF2a in the present study. Fierro et al. (2011) reported prolificacy of 1.3 offspring and a fecundity of 0.8 offspring, which corroborates results obtained in the present study. The results obtained for fecundity in the present study are attributed to the "male effect", the type of insemination and the synchronization protocol, because after introducing the male, the pre-ovulatory discharge of LH occurred from 3 to 30 hours and ovulation occurred between 24 to 60 hours. However, ovulation induced by the "male effect" was not preceded by signs of oestrus and was characterized by short oestrus cycles (less than seven days) and low secretion of luteal P4. That is, CL of short duration and silent oestrus are observed, which then re-establish normal oestrus cycles and fertility (Oldham & Martin, 1978).
Conclusions
Synchronizing post-partum ovarian activity with the "male effect" and "temporary weaning" reduces the onset of oestrus and the rate of return to oestrus, but increases the response to oestrus and fecundity in Pelibuey ewes. Therefore, compared with other studies, this reproductive management could be recommended to improve the fertility of Pelibuey ewes synchronized with two doses of prostaglandins at 45 days post-partum.
Acknowledgements
The authors wish to thank Colegio de Postgraduados for the support from Fideicomiso No. 167304/2010-2011 to LPI-5 Biotecnología Microbiana Vegetal y Animal (Plant and Animal Microbial Biotechnology), the Laboratorio de Reproducción de Ovinos y Caprinos (Laboratory for Sheep and Goat Reproduction) for supporting the realization of the reproductive management protocols, and Humberto Vaquera Huerta for helping with the statistical analyses.
Authors' Contributions
JAHM carried out the research work and wrote the manuscript, based on the JAHM's PhD thesis, under the supervision of JGS, CCR, CAHR, PPH and APM. Data were collected by JAHM, and analysis of data and interpretation of results was led by JGS with assistance from PPH, CCR, APM and CAHR. Critical review and final approval of the version to be published were done by JGS, JAHM and PPH.
Conflict of Interest Declaration
The authors certify that they have no affiliations with any organisation or entity with any financial or non-financial interest in the subject matter or materials discussed in this manuscript.
References
Abecia, J.A., Forcada, F. & González-Bulnes, A., 2011. Pharmaceutical control of reproduction in sheep and goats. Vet. Clin. Food Anim. 27, 67-79. [ Links ]
Abecia, J.A., Forcada, F. & González-Bulnes, A., 2012. Hormonal control of reproduction in small ruminants. Anim. Reprod. Sci. 130, 173-179. [ Links ]
Arteaga, C.M.L., Martínez-Gómez, M., Guevara-Guzmán, R. & Hudson, R., 2007. Comunicación química en mamíferos domésticos. Vet. Méx. 38, 105-123. [ Links ]
Bari, F., Khalid, M., Haresign, W., Murray, A. & Merrel, B., 2003. Factors affecting the survival of sheep embryos after transfer within a MOET program. Theriogenol. 59, 1265-1275. [ Links ]
Barret, D.M.W., Bartlewski, P.M., Cook, S.J. & Rawlings, N.C., 2002. Ultrasound and endocrine evaluation of the ovarian response to PGF2a given at different stages of the luteal phase in ewes. Theriogenol. 58, 1409-1424. [ Links ]
Camacho-Ronquillo, J.C., Rodríguez-Castillo, J.C., Hernández-Hernández, J.E., Pro-Martínez, A., Becerril-Pérez, C.M. & Gallegos-Sánchez, J., 2008. Características reproductivas de ovejas Pelibuey sincronizadas e inducidas a la pubertad. Arch. Lat. Prod. Anim. 16, 18-24. [ Links ]
Castillo-Maldonado, P.P., Vaquera-Huerta, H., Tarango-Arambula, L., Pérez-Hernández, P., Herrera-Corredor, A. & Gallegos-Sánchez, J., 2013. Restablecimiento de la actividad reproductiva posparto en ovejas de pelo. Arch. Zoot. 62(239), 419-428. [ Links ]
Contreras-Solís, I., Vásquez, B., Díaz, T., Letelier, C., López-Sebastian, A. & González-Bulnes, A., 2009. Ovarian and endocrine responses in tropical sheep treated with reduced doses of cloprostenol. Anim. Rep. Sci. 114, 384-392. [ Links ]
Cortez-Romero, C., Herrera-Corredor, C.A., Gallegos-Sánchez, J. & Salazar-Ortíz, J., 2011. Manual sobre fisiología de la reproducción, inseminación artificial y ultrasonografía en ovinos. ISBN: 978-607-715-024-4. Colegio de Postgraduados. Fundación Grupo Produce A.C., México, D.F. 87 pp. (in Spanish). [ Links ]
Das, G.K., Naqvi, S.M.K., Gulyani, R., Joshi, A. & Mittal, J.P., 1999. Effect of two protocols of PGF2a treatment for synchronization of estrus in a tropical sheep. Theriogenol. 51, 283. [ Links ]
Davis, A.J., Fleet, I.R., Harrison, F.A. & Walker, F.M.M., 1980. Pulmonary metabolism of prostaglandin F2a in the conscious non-pregnant ewe and sow. J. Physiol. 301, 86. [ Links ]
Dixon, A.B., Knights, M., Winkler, J.L., Marsh, D.J., Pate, J.L., Wilson, M.E., Dailey, R.A., Seidel, G. & Inskeep, E.K., 2007. Patterns of late embryonic and fetal mortality and association with several factors in sheep. J. Anim. Sci. 85, 1274-1284. [ Links ]
Fierro, S., Gil, J., Viñoles, C. & Olivera-Muzante, J., 2013. The use of prostaglandins in controlling estrous cycle of the ewe: A review. Theriogenol. 79, 399-408. [ Links ]
Fierro, S., Olivera-Muzante, J., Gil, J. & Viñoles, C., 2011. Effects of prostaglandin administration on ovarian folicular dynamics, conception, prolificacy, and fecundity in sheep. Theriogenol. 76, 630-639. [ Links ]
García, E., 2004. Modifications to the System Köppen climate classification. 5th edition. Book series 6. Institute of Geography, Universidad Nacional Autónoma de México. p. 51. (in Spanish). [ Links ]
Hernández-Marín, J.A., Pro-Martínez, A., Cortez-Romero, C., Pérez-Hernández, P., Herrera-Corredor, C.A. & Gallegos- Sánchez, J., 2016. Inducción de la ovulación con efecto macho y un reconstituyente energético en ovejas Pelibuey prebúberes. Agrociencia 50: 811-823. [ Links ]
Kumar, D. & Naqvi, S.M.K., 2014. Effect of time and depth of insemination on fertility of Bharat Merino sheep inseminated trans-cervical with frozen-thawed semen. J. Anim. Sci. Technol. 56, 8. [ Links ]
Letelier, C.A., Contreras-Solis, I., García-Fernández, R.A., Sánchez, M.A., García-Palencia, P., Sánchez, B., Ariznavarreta, C., Tresguerres, J.A.F., Flores, J.M. & González-Bulnes, A., 2011. Effects of oestrus induction with progestagens or prostaglandin analogues on ovarian and pituitary function in sheep. Anim. Reprod. Sci. 126, 61-69. [ Links ]
Martemucci, G. & A.G. D'Alessandro., 2011. Synchronization of oestrus and ovulation by short time combined FGA, PGF2, GnRH, eCG treatments for natural service or AI fixed-time. Anim. Reprod. Sci. 123, 32-39. [ Links ]
Martin, G.B., Milton, J.T.B., Davidson, R.H., Banchero, G.E., Lindsay, D.R. & Blache, D., 2004. Natural methods of increasing reproductive efficiency in sheep and goats. Anim. Reprod. Sci. 82-83, 231-46. [ Links ]
McKelvey, W.A.C., Robinson, J.J. & Aitken, R.P., 1985. The evaluation of a laparoscopic insemination technique in ewes. Theriogenol. 24, Suppl. 5, 519-535. [ Links ]
Meilán, J. & Ungerfeld, R., 2014. Does introduction of rams during the late luteal phase promote the estrus response in cyclic ewes? Short communication. Small Rum. Res. 120, 116-120. [ Links ]
Naqvi, S.M.K., Maurya, V.P., Gulyani, R., Joshi, A. & Mittal, J.P., 2004. The effect of thermal stress on superovulatory response and embryo production in Bharat Merino ewes. Small Rum. Res. 55, 57-63. [ Links ]
Oldham, C.M. & Martin, G.B., 1978. Stimulation of seasonally anovular ewes by rams. II. Premature regression of ram- induced corpora lutea. Anim. Reprod. Sci. 1, 291-295. [ Links ]
Oldham, C.M., Thompson, A.N., Ferguson, M.B., Gordon, D.J., Kearney, G.A. & Paganoni, B.L., 2011. The birth weight and survival of Merino lambs can be predicted from the profile of live-weight change of their mothers during pregnancy. Anim. Prod. Sci. 51, 776-783. [ Links ]
Olivera-Muzante, J., Gil, J., Viñoles, C. & Fierro, S, 2013. Reproductive outcome with GnRH inclusion at 24 or 36 h following a prostaglandin F2-based protocol for timed AI in ewes. Anim. Reprod. Sci. 138, 175-179. [ Links ]
Olivera-Muzante, J., Gil, J., Fierro, S., Menchaca, A. & Rubianes, E., 2011. Alternatives to improve a prostaglandin- based protocol for timed artificial insemination in sheep. Theriogenol. 76, 1501-1507. [ Links ]
Rodríguez, O.L., Heredia, M., Quintal, F. & Carrillo, L., 1986. Manejo de la lactación para incrementar la eficiencia reproductiva en ovejas Pelibuey. I. Presencia del cordero en destetes temporales. Téc. Pec. Mex. 51, 104-110. [ Links ]
Rodríguez-Iglesias, R.M., Ciccioli, N.H., Ferrería, J., Pevsner, D.A., Rosas, C.A., Rodríguez, M.M. & Pedrueza, J.R., 2013. Short-lived corpora lutea syndrome in anoestrous ewes following 17ß-oestradiol or MAP treatments applied before an allogenic sexual stimulation with rams and oestrous ewes. Anim. Reprod. Sci. 136, 268-279. [ Links ]
Rubianes, E., Menchaca, A. & Carbajal, B., 2003. Response of the 1-5 day-aged ovine corpus luteum to prostaglandin F2a. Anim. Reprod. Sci. 78, 47-55. [ Links ]
Salinas, F.M., 2008. Modelos de Regresión VI. Análisis de supervivencia. Ciencia y Trabajo. 28, 75-78. [ Links ]
SAGARPA. Diario Oficial de la Federación. Norma Oficial Mexicana 024 y 033-ZOO-1995. Accessed June 2015. URL: http://sagarpa.gob.mx/normateca/normateca2/SENASICA%20NORM%2028.pdf and http://dof.gob.mx/nota_detalle.php?codigo=4883147&fecha=16/10/1995 [ Links ]
SAS Institute Inc., 2012. Statistical Analysis System. SAS user's guide statistics, version 9.4 (TS1MO). SAS Institute Cary, N.C., USA. [ Links ]
Shapiro, S.S. & Wilk, M.B., 1965. An analysis of variance test for normality. Biometrika. 52, 591-611. [ Links ]
Torres-Zapata, S., Luna-Palomera, C., Aguilar-Cabrales, J.A., Peralta-Torres, J.A., Aké-López, J.R., Sánchez-Dávila, F. & Abad-Zavaleta, J., 2016. Ovulatory response and embryo quality in Katahdin ewes supplemented with palm oil. S. Afri. J. Anim. Sci. 46, Suppl. 3, 261-368. [ Links ]
Ungerfeld, R., 2011. Combination of the ram effect with PGF2a estrus synchronization treatments in ewes during the breeding season. Anim. Reprod. Sci. 124, 65-68. [ Links ]
Walkden-Brown, S.W., Restall, B.J. & Henniawati. 1993. The male effect in the Australian Cashmere goat. 3. Enhancement with buck nutrition and use of oestrus females. Anim. Reprod. Sci. 32, 69-84. [ Links ]
Received 17 December 2017
Accepted 5 July 2018
First published online 7 September 2018
# Corresponding author: gallegos@colpos.mx


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}