










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.4 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i4.14
ARTICLE
Digestible threonine and its effects on growth performance, gut morphology and carcass characteristics in broiler Japanese quails (Coturnix coturnix japónica)
M.F. RasheedI; M.A. RashidI, #; SaimaI; A. MahmudII; M.S. YousafIII; M.I. MalikI
IDepartment of Animal Nutrition, University of Veterinary and Animal Sciences, Faculty of Animal Production and Technology, 54000 Lahore, Pakistan
IIDepartment of Poultry Production, University of Veterinary and Animal Sciences, Faculty of Animal Production and Technology, 54000 Lahore, Pakistan
IIIDepartment of Physiology, University of Veterinary and Animal Sciences, Lahore-54000, Pakistan
ABSTRACT
Threonine is the third limiting amino acid in corn-soy-based poultry diets and has an important role in body and intestinal development of chicken. Although work on threonine (Thr) in chicken is well established, information about the effects of digestible threonine (dThr) on quail performance is limited. Therefore, the current study was designed to determine the impact of increasing levels of dThr on growth performance, feed conversion ratio (FCR), gut health and carcass characteristics in meat-type quails. A total of 324 mixed-sex day-old quail were randomly allocated to three treatments with six replicates per treatment and 18 birds per replicate. There were three dietary treatments, namely control (C) with recommended levels of dThr according to the Brazilian Tables guidelines for Japanese quails: 10% dThr (C diet supplemented with 10% more dThr) and 20%dThr (C diet supplemented with 20% more dThr). At day 35 of the experiment, three birds from each pen were slaughtered. A duodenal sample was collected and preserved to evaluate gut health. The carcass characteristics were determined from the slaughtered birds. Total feed intake and average daily feed intake were higher in the C treatment than in 10% dThr. Final bodyweight (BW), weight gain and average daily gain (ADG) increased linearly. The birds fed diets supplemented with 20% extra dThr had the highest final weight and bodyweight gain (BWG) compared with those birds that were fed on C and 10% dThr supplemented diets. Feed conversion ratio was improved in 10% dThr compared with the C birds. Villus height (VH) was similar among treatments. The highest crypt depth (CD) was observed in C, followed by 10% dThr and 20% dThr. The birds fed 20% dThr had higher VH:CD than 10% dThr. The birds in the C treatment had lowest VH:CD. Carcass weights with and without giblets were higher in the 20% dThr than in the 10% dThr and C treatments. Breast mass yield (BMY) was greatest in 20% dThr compared with C and 10% dThr. It may be concluded that supplementation of Thr higher than the requirements referred to in Brazilian Tables improves growth performance and gut health of meat-type quail.
Keywords: Crypt depth, mucin, performance, quail, villus height
Introduction
Threonine is a 2-amino-3-hydroxy butanoic acid with a molecular mass of 119.2 g/mol (Ayasan, 2004). For low protein diets, it is the third limiting amino acid in broilers (Kidd et al., 1999). Threonine is known to help to maintain gut barrier integrity and intestinal mucin synthesis (Bertolo et al., 1998). It is integrated directly into mucin, which is a major glycoprotein for the protection of intestinal linings from injury (Schaart et al., 2005). A deficiency of threonine (Thr) leads to a decrease of mucin production in the small intestine in rats (Faure et al., 2005). The layer of mucus is an essential part of the non-immune gut barrier. However, during the inflammation process, Thr accessibility may become restricted for mucin production, which impairs the gut protection system. Increasing dietary Thr inclusion may stimulate the synthesis of mucin and gut microbiota to favour mucosal healing and protect the intestine (Faure et al., 2006; Wang et al., 2009).
To date, several other investigators have evaluated dietary Thr requirements in broilers (Rosa et al., 2001; Rezaeipour et al., 2012; Rezaeipour & Gazani, 2014). The function of Thr is to maintain the digestive tract (Van der Schoor et al., 2002) and improve productivity (Kidd & Kerr, 1997; Kidd et al., 2003). It plays many roles, namely as a feather protein, a precursor of serine and glycine, a component of glycoprotein involved in gastrointestinal tract (GIT) mucin production (Lemme, 2001) and stimulate antibody production (Wang et al., 2006). Threonine, as, L isomer, is supplemented in the diets of commercial poultry and pigs to meet the nutritional requirements of amino acids for rapidly growing animals. Balancing essential amino acids enables animals to utilize feed more efficiently. To date, several studies have assessed whether dietary supplementation of Thr could improve the performance of pigs, layers, broilers and rabbits. Threonine requirements have been calculated for broilers from 0 to 8 weeks old (Kidd et al., 1997; Dozier et al., 2001), but the literature on Thr requirements for Japanese quail is limited. Japanese quail are raised mainly for meat and eggs, and are regarded as research animals (NRC, 1994). Their feeding requirements are lower (20-25 g/day) than those of chickens (120-130 g/day) (Ani et al., 2009). Typically, quail are marketed at 140-180 g at 5-8 weeks and reach peak egg production at the age of 5-8 weeks (Garwood & Diehl, 1987). Broiler quail Thr requirements are established and published in National Research Council (1994) and Brazilian Tables for Poultry and Swine (2011). Their requirements of digestible Thr are 1.02% and 0.79%, respectively.
Although work on Thr in chicken is well established, information about the effects of dietary L-Thr on quail performance is limited. The authors' aim was to evaluate dThr supplementation in quail diet using the Brazilian Tables (2011).
Materials and Methods
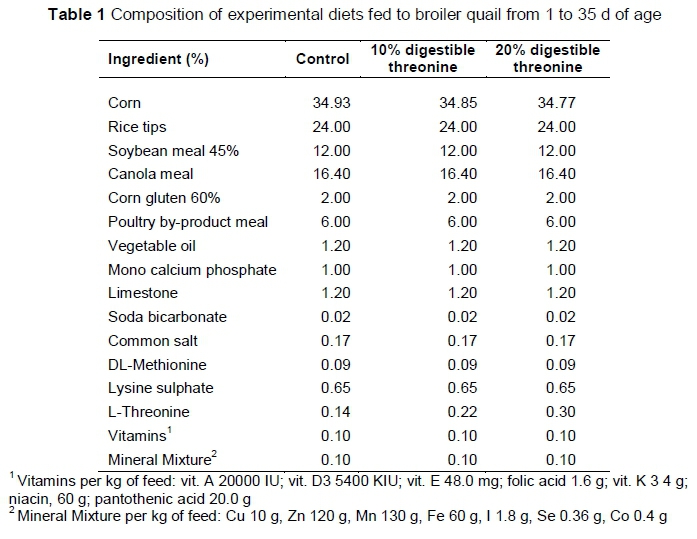
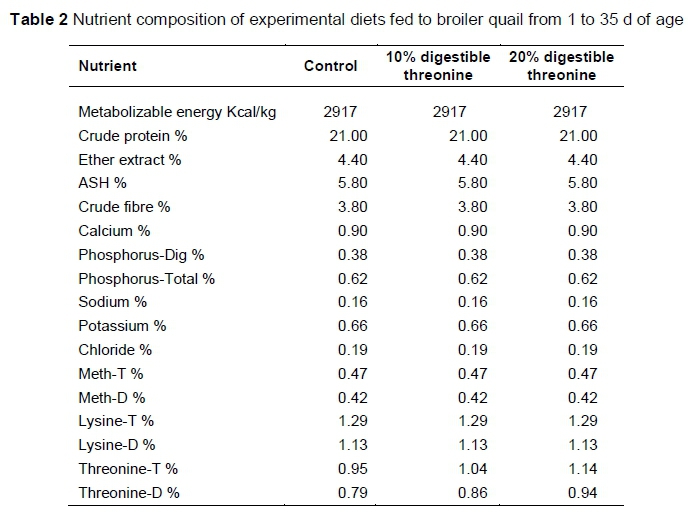
The experiment was performed at a commercial quail farm. Birds were reared in a mechanically ventilated quail battery system. Before the start of experiment, the shed was disinfected with a 35 mL solution of formalin, containing 40% formaldehyde and 10 KMnO4 g/ft3 according to farm protocols. A total of 324 day-old chicks were selected from the farm's own hatchery and randomly assigned to three dietary treatments with 108 chicks per treatment. Every treatment was further divided into six replicates with 18 chicks per replicate. The dietary treatments consisted of i) C, the control diet with the standard digestible threonine (dThr) level according to the Brazilian Tables for Poultry and Swine (2011); ii) 10% dThr, with 10% higher digestible threonine (dThr) than control; and iii) 20% dThr, with 20% higher dThr than control. The diets were formulated according to the Brazilian Tables for Poultry and Swine (2011). Diet formulation and nutrient composition are presented in Tables 1 and 2, respectively.
Chicks were given free access to experimental diets and fresh water was provided in each pen. Brooding temperature for the first week was kept at 95 °F. Every week the temperature was reduced by 5 °F until it reached room temperature (75 °F). Whole weight and bird mortality per replicate were recorded at the end of every week. A calculated amount of feed was offered to all birds in each replicate for the whole week. After the completion of each week, feed refusals were weighed to calculate the weekly feed intake of each replicate. Mortality weight was used to adjust the feed intake of that replicate. The birds in each replicate were weighed at day one and after that on a weekly basis. The data of the feed eaten by birds and gain in bodyweight, corrected for mortality, were used to calculate feed conversion ratio (FCR) as feed consumed by bird (g) divided by weight attained by bird (g).
At the end of the fifth week, three birds from each replicate were selected, weighed and humanely slaughtered for organ weights and morphometric measurements. Duodenal samples for morphometric study were collected from slaughtered birds as described by Gopinger et al. (2014). A section 2 cm long was collected from the middle of the duodenum, cleaned with normal saline (0.9% NaCl) and preserved in a volumetric plastic container with 10% formalin solution. Preserved intestinal sections were fixed in 10% formalin solution. Fixed tissue sections were dehydrated in ascending concentrations of alcohol (50%, 70%, 95%, absolute I, and absolute II) for cleaning. Dehydrated tissue samples were cleared in pure xylene. After clearing, the tissue samples were impregnated with paraffin wax I and paraffin wax II. The paraffin-impregnated tissue samples were divided into 5-|jm thick sections with microtome and mounted on clean glass slides (covered with a thin layer of egg albumen), after slight warming at 58 °C in a hot water bath. The slides were dipped in pure xylene I and pure xylene II for 5 minutes each. The slides were then dipped in descending concentrations of alcohol (absolute, 90%, 80% and 70%) for 5 minutes each. Then the slides were dipped three times in distilled water. The slides were placed in haematoxylin stain for 2 to 3 minutes. The slides were washed under tap water for 1 to 3 minutes, dipped three times in distilled water and ammonia water (two dips) and stained with eosin for 1 to 2 minutes. After staining the samples were washed under tap water (1 to 2 minutes) and dehydrated in ascending concentrations of alcohol (70%, 80%, 90%, and absolute) for 3 minutes each. The samples were placed in xylol I and xylol II for 3 minutes each. A drop of DPX (distyrene plasticizer xylene) was placed on the slides and a cover slip was applied. The tissue slides were examined for VH and CD with a compound microscope (Olympus CX31, Olympus USA) equipped with a digital imaging system (Olympus DP20, Olympus USA).
Carcass evaluation was carried out. The slaughtered birds were cut into parts for carcass evaluation according to Ojewola et al. (2001). Breast yield and carcass with and without giblets were weighed with a digital balance. Liver, heart, gizzards and non-edible organs were weighed. The gastrointestinal tract (GIT) with and without ingesta were weighed and the weights of the relative organs were calculated.
The data for total feed intake, average daily feed intake, final weight, weight gain, average daily gain (ADG), FCR, villus height (VH), crypt depth (CD), VH:CD and carcass characteristics were analysed with one-way ANOVA in a completely randomized design. Means were compared with Tukey's multiple comparison test. All the data were analysed with SPSS software (SPSS version 21.0 Inc., Chicago, IL, USA).
Results
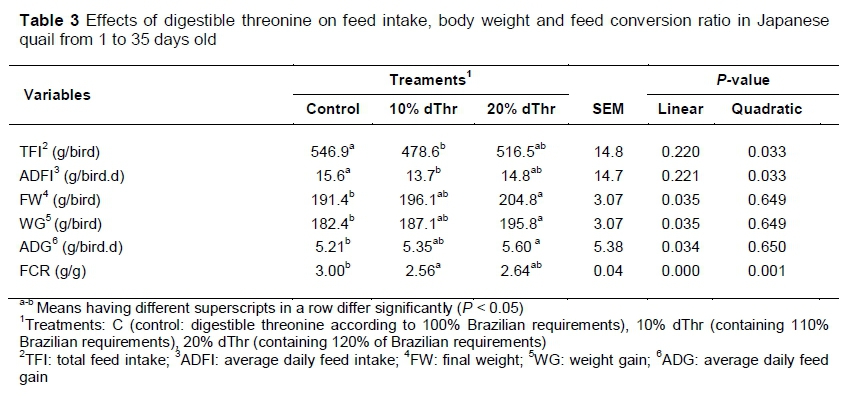
Means of total feed intake, average daily feed intake, weight gain, final weight, ADG and FCR are presented in Table 3.
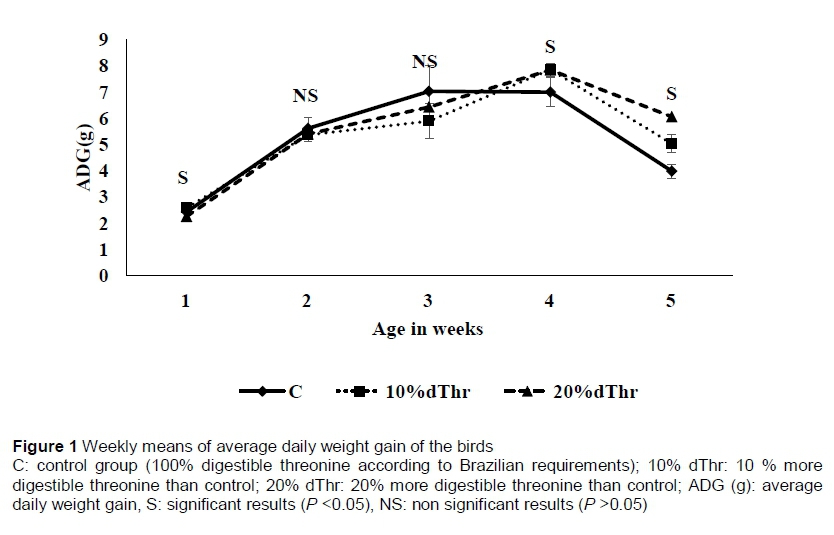
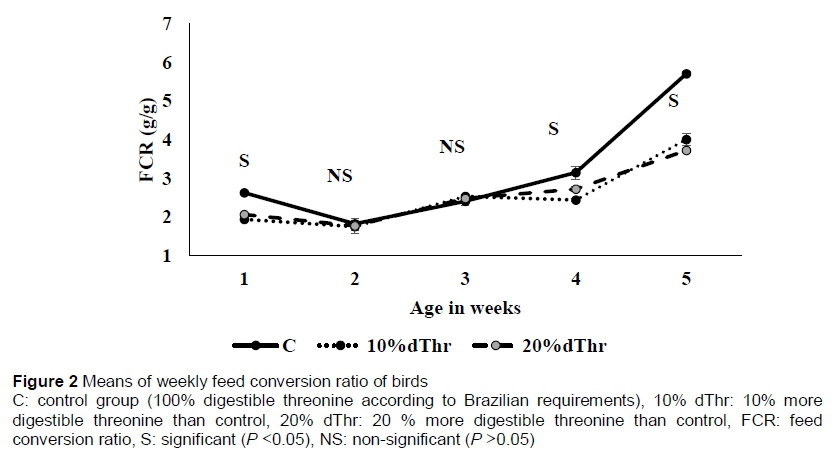
The total feed intake and average daily feed intake were significantly higher (quadratic) (P <0.05) in C than in 10% dThr. Final body weight (BW), weight gain and ADG increased linearly (P <0.05) with level of dThr in the diets. The birds fed diets supplemented with 20% dThr had the highest final weight and body weight gain (BWG) compared with those birds fed on C and 10% dThr, respectively. A treatment x week interaction was observed (P <0.05) for ADG in weeks 1, 4, and 5 (Figure 1). In week 1, ADG was significantly higher (P <0.05) in the 10% dThr quails than in the C and 20% dThr birds. During week 4, however, the growth rate was higher in the 10% dThr and 20% dThr birds than the C treatment. Interestingly, birds fed 20% dThr showed higher ADG than those in the 10% dThr and C treatments. The results showed clearly that FCR was improved (P <0.05) in the dTH10% compared with the C birds. A treatment x week interaction was observed in weeks 1, 4, and 5 (Figure 2).
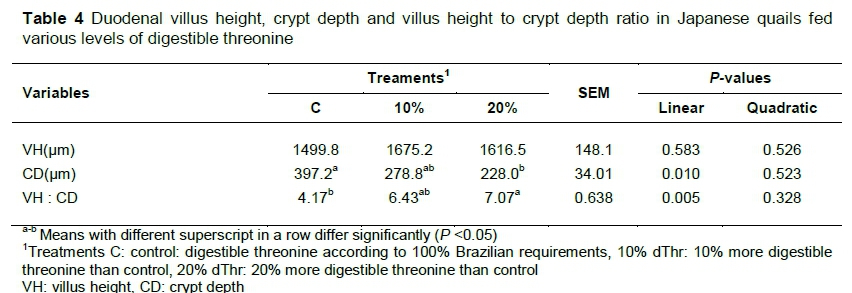
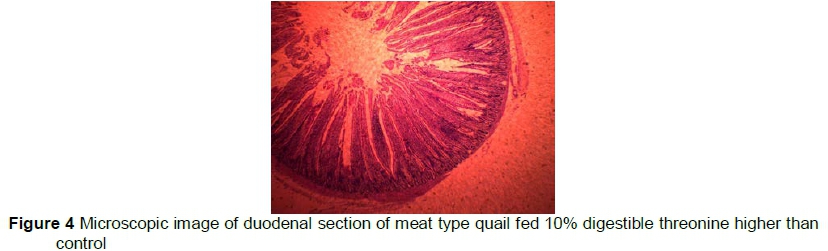
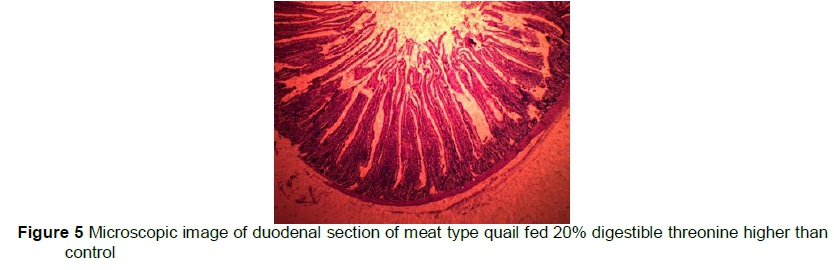
Means of VH, CD, and VH:CD ratio are presented in Table 4. VH was similar (P >0.05) among treatments. C had numerically less VH compared with those fed 10% and 20% extra dThr. CD increased (P <0.05) linearly in C compared with those fed the 20% dThr supplemented diet. The birds fed the 20% dThr had the highest (P<0.05) VH:CD, followed by 10% dThr and C.
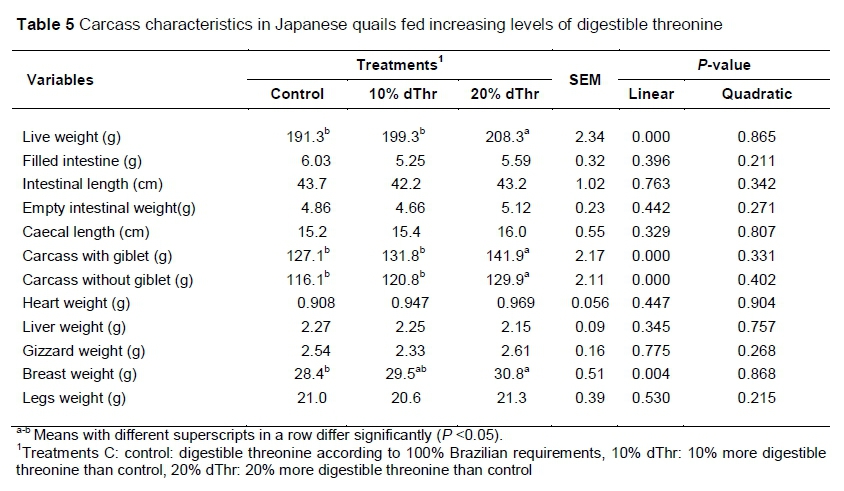
The results of the evaluation of carcass characteristics are shown in Table 5. Increasing levels of dThr in quail had no effects (P >0.05) on filled intestine, empty intestine, intestinal length, caecal length and leg weight. Carcass weight with and without giblets was higher (P <0.05) in 20% dThr compared with 10% dThr and C treatments. Similarly, heart, gizzard and liver weights were not influenced (P >0.05) by increasing levels of dThr. The birds fed on 20% dThr had heavier BMY (P <0.05) than the C and 10% dThr diets.
Discussion
It was hypothesized that the growth performance of quails could be improved with supplementation of Thr above the recommended levels. Gut health and carcass characteristics were studied as descriptive variables.
The results of the evaluation of growth performance of the present study showed that average daily and total feed intake were highest in the C treatment. In the C treatment, total and average daily feed intake were higher by 14.2% and 5.89% compared with the 10% dThr and 20% dThr diets, respectively. Final BW, weight gain and ADG increased linearly with increasing levels of dThr in the diets. The current results are in line with those of Lehmann et al. (1997), who supplemented Thr (0.82-0.88-0.94-1.00-1.06-1.12%) and stated that weight gain rose with increasing threonine in the diets of growing turkey toms. The data of the current experiment showed that 21.9% lower FCR was observed in C compared with 10% dThr. Similarly, Canogullari et al. (2009) stated that increasing Thr levels in laying quails improved FCR. In contrast to present study, Baylan et al. (2006) stated that increased dThr from 0.81 % to 1.06 % did not affect FCR in quails during the 1-35 day period. Poor FCR in C was due to poor utilization of feed compared with those fed 10% dThr and 20% dThr. Better FCR in 10% dThr and 20% dThr may be because threonine improved nutrient digestion owing to higher amylase secretion in the digestive tract (Block et al., 1966). Improved FCR in 10% dThr may be associated with improved nitrogen retention. Similarly, Dozier et al. (2001) stated that nitrogen retention was higher in broilers fed 0.74% of total Thr compared with 0.52% Thr in the diet.
Work by Block et al. (1966) showed that Thr constitutes 11% of amylase protein contents, suggesting the importance of dThr in diets of growing birds, and consequent improvement in growth rate. Other investigators suggested that Thr serves as a precursor of lysine and serine and plays a vital role in body proteins (Ojano-Diranin & Waldroup, 2002). Similarly, Good (2013) conducted an experiment to find out the interaction of Actigen and graded levels of Thr (0.56%, 0.64%, 0.80%, 0.96%, and 1.04%). The author stated that bird fed diets containing the recommended level of Thr (0.56%) had the lowest BWG, feed intake, and poor FCR compared with those fed various levels of Thr (0.64, 0.80, 0.96, and 1.04%). Ospina-Rojaset et al. (2013) reported improved FCR in broilers fed diets containing 10% (0.77%) more Thr than Ross's recommendations (0.70%), which is in line with the current findings in broiler quails. Estalkhzir et al. (2013) stated that 110% and 115% levels of Thr resulted in improved BWG, FCR, and lowered feed intake. Mazraeh et al. (2013) reported improved BWG and FCR in broiler chicken. However, feed intake was not similar among treatments, indicating the impact of Thr on nutrient utilization in meat-type birds.
In this experiment, higher growth rates, lower feed intake and better FCR in 10% dThr and 20% dThr compared with C may be attributed to the stabilization and maintenance of gut barriers (Bertolo et al., 1998; Schaart et al., 2005). One possible explanation for improved growth performance and FCR could be that Thr is a major component of mucin, a glycoprotein, which serves as a barrier and protects intestinal linings from injury. Mucin also serves as a protecting material from acidic chyme and pathogenic microbes (Horn et al., 2009). As a glycoprotein, mucin filters nutrients inside the gut lumen and helps in digestion and absorption (Smirnov et al., 2006).
Intestinal health and fitness of gut for digestion and absorption can be determined by measuring the VH and CD (Awad et al., 2009). In the small intestines, the duodenum is the principal site of absorption and digestion of nutrients. Duodenal morphometry was therefore used to indicate the impact of dietary treatments on gut health. An in vivo study in piglets showed that between 80% and 90% of dietary Thr is used by the intestine, most of which is incorporated into mucosal proteins (Schaart et al., 2005). The published data regarding Thr supplemental effects in quail are insufficient. The VH in meat type quails was similar and not influenced by the increasing level of dThr. The results in the present study are similar to those of Chen et al. (2016), who reported that supplementation of Thr (0.70% and 0.77%) did not improve VH in broilers from day 1 to day 35 of their experiment. Similarly, Eftekhari et al. (2015) reported that increasing levels of Thr (100%, 110%, 120% and 130%) did not influence VH in broilers. Good (2013) supplemented various levels of threonine (0.56%, 0.64%, 0.80%, 0.96% and 1.04%) and observed that VH was lower in birds fed 0.56% of threonine, and associated this decrease in VH to a deficiency of Thr in the diet. In broilers, those receiving ample amounts of threonine in the diet had better VH compared with those fed a Thr-deficient diet (Zaefarian et al., 2008). Contrary to the current findings, a study evaluating the supplementation of higher Thr to Lys ratio resulted in improved gut health in broilers. The authors supplemented the Thr deficient diet with recommended or 25% greater Thr levels and reported that supplementation of higher levels of Thr resulted in improved VH in broilers (Valizade et al., 2014).
The depth of crypt gives an estimate of the number of cells in intestinal villi. A larger CD therefore indicates a greater turnover rate of enterocytes, which require more protein and energy and vice versa (Abdullah et al., 2010). The current results showed that decreases in CD were observed in the 10% dThr and 20% dThr by 29.8% and 42.6 %, respectively, compared with C. These results are in line with those of Abbasi et al. (2014), whose supplementation of 10% more Thr than the recommended level resulted in a 6% increase in VH, an 8.3% decrease in CD and a 12% increase in the VH to CD ratio. Intestinal mass and viability are maintained by dietary proteins, which serve as a source of energy for normal intestinal functions. The small intestine uses 30% to 50% Thr, along with other amino acids directly and does not spare them for other tissues, except the intestine. Lower CD in Thr-supplemented birds may be associated with a rapid turnover of intestinal tissues through an ample supply of dThr. The current results are in line with those of Valizade et al. (2014), who found that higher levels of Thr resulted in improved CD and VH:CD ratio in broilers.
Contrary to the current findings, Chen et al. (2016) reported that supplementation of dThr did not influence the CD and VH:CD ratio in broilers. Eftekhari et al. (2015) reported that various levels of Thr (100%, 110%, 120% and 130%) did not influence CD and VH:CD ratio in broilers. Similarly, Good (2013) reported that supplementation of Thr at various levels (0.56%, 0.64%, 0.80%, 0.96% and 1.04%) did not influence CD and the VH:CD ratio in the jejunum of broilers at seven days old. The higher dietary fibre might have caused sloughing of intestinal mucosa (Mushtaq et al., 2009), which in turn increases the amino acid requirements for the synthesis of new cells (Parsons et al., 1983). Choct (2009) reported that fast growing broilers allocate 12% of newly synthesized protein to the GIT. The lower CDs in the current experiment indicate a slower tissue turnover. Thus, a lower nutrient supply is required to support it (Boka et al., 2014). Threonine plays a significant role in the development of the intestines of chickens because it is involved in the synthesis of mucin (Faure et al., 2005).
Carcasses with and without giblets were higher in 10% dThr and 20% dThr by 3.70% and 11.64%, respectively, compared with the C treatment quails. Breast meat yield is the most desired part in the carcass of birds. In addition to lysine and methionine, Thr plays a key role in the development of breast muscles in birds. A suboptimal supply of dietary Thr may decrease breast meat growth breast meat yield by 3.8% and 8.45%, respectively, less than C. The current results are in accordance with the Estalkhzir et al. (2013). The authors reported 11.48% improved carcass weight, and 29.85% greater breast meat yield in broiler chickens. However, in quails, the response was low compared with broilers, which may be associated with genetically higher growth and better FCR potential in commercial meat type broilers. Thus, Baylan et al. (2006) stated that supplemental L-threonine did not affect carcass yield, breast meat, and thigh percentage in quails. For broilers, the requirement of threonine for carcass gain differs, depending on age, strain, sex and crude protein content of feed, dose and types of ingredients (Barkley & Wallis, 2001). Similarly, several researchers reported that Thr supplementation increased the breast meat yield (Mack et al., 1999; Dozier et al., 2000, 2001; Ciftci & Ceylan, 2004; Jahanian, 2010), which is in line with the current findings. In the current experiment supplementation of dThr had no effect on thigh weight, which corroborates the findings of Kerr et al. (1999) that supplementation of dThr did not influence the thigh yield.
Conclusions
In conclusion, the supplementation of dThr above the recommendations of Brazilian Tables for Poultry and Swine improved growth performance, FCR, gut health, and breast meat yield of broiler-type Japanese quails.
Authors' Contributions
M.F. Rasheed: Data collection, analysis and write up; M.A. Rashid: Experiment planning, execution of experiment, lab analysis and write up; Saima: Lab analysis; A. Mahmud:, Statistical analysis; M.S. Yousaf: Lab analysis and write up; M.I. Malik: Lab analysis and write up.
Conflict of Interest Declaration
The authors declare no conflict of interest.
References
Abbasi, M., Mahdavi, A., Samie, A. & Jahanian, R., 2014. Effects of different levels of dietary crude protein and threonine on performance, humoral immune responses and intestinal morphology of broiler chicks. Rev. Bras. Cienc. Avic. 16, 35-44. [ Links ]
Abdullah, A.Y., Mahmoud, K.Z., Nusairat, B.M. & Qudsieh, R.I., 2010. Small intestinal histology, production parameters, and meat quality as influenced by dietary supplementation of garlic (Allium sativum) in broiler chicks. Ital. J. Anim. Sci. 9, 80. [ Links ]
Ani, A., Okeke, G. & Emeh, M., 2009. Response of growing Japanese quail (Cortunix Cortunix japónica) chicks to diets containing different energy and protein levels. In: Diversifying Nigerian economy animal production options. Proceedings of the 34th Annual Conference of the Nigerian Society for Animal Production 15, 328-331. [ Links ]
Awad, W., Ghareeb, K., Abdel-Raheem, S. & Böhm, J., 2009. Effects of dietary inclusion of probiotic and synbiotic on growth performance, organ weights, and intestinal histomorphology of broiler chickens. Poult. Sci. 88, 49-56. [ Links ]
Ayasan, T., 2004. Investigation of threonine requirements of broilers. PhD thesis, Agricultural Faculty, Animal Science, University of Cukurova, Adana. [ Links ]
Barkley, G. & Wallis, I., 2001. Threonine requirements of broiler chickens: Why do published values differ? Br. Poult. Sci. 42, 610-615. [ Links ]
Baylan, M., Canogullari, S., Ayasan, T. & Sahin, A., 2006. Dietary threonine supplementation for improving growth performance and edible carcass parts in Japanese quail, Coturnix coturnix Japónica. Inter. J. Poult. Sci. 5, 635-638. [ Links ]
Bertolo, R.F., Chen, C.Z., Law, G., Pencharz, P.B. & Ball, R.O., 1998. Threonine requirement of neonatal piglets receiving total parenteral nutrition is considerably lower than that of piglets receiving an identical diet intragastrically. Br. J. Nutr. 128, 1752-1759. [ Links ]
Block, R.J., Weiss, K.W. & Cornett D.B., 1966. The amino acid composition of proteins. In: R.J. Block & D. Bolling (eds). The Amino Acid Composition of Proteins and Foods. Charles C. Thomas, Springfield, IL. pp. 149-295 [ Links ]
Boka, J., Mahdavi, A., Samie, A. & Jahanian, R., 2014. Effect of different levels of black cumin (Nigella sativa L.) on performance, intestinal Escherichia coli colonization and jejunal morphology in laying hens. J. Anim. Physio Anim. Nut. 98(2), 373-383. [ Links ]
Brazilian Tables for Poultry and Swine 2011. Composition of feedstuffs and nutritional requirements 2011. 3rd edition. Brazil: UFV Viçosa. [ Links ]
Canogullari, S., Baylan, M. & Ayasan, T., 2009. Threonine requirement of laying Japanese quails. J. Anim. Vet. Adv. 8(8), 1539-1541. [ Links ]
Chen, Y.P., Cheng, Y.F., Li, X.H., Yang, W.L., Wen, C., Zhuang, S. & Zhou, Y.M., 2016. Effects of threonine supplementation on the growth performance, immunity, oxidative status, intestinal integrity, and barrier function of broilers at the early age. Poult. Sci. 96, 405-413. [ Links ]
Choct, M., 2009. Managing gut health through nutrition. Br. Poult. Sci. 50, 9-15 [ Links ]
Ciftci, I. & Ceylan, N., 2004. Effects of dietary threonine and crude protein on growth performance, carcase and meat composition of broiler chickens. Br. Poult. Sci. 45, 280-289. [ Links ]
Dozier, W., Moran, E. & Kidd, M., 2000. Threonine requirement of broiler males from 42 to 56 days in a summer environment. J. Appl. Poult. Res. 9, 496-500. [ Links ]
Dozier, W., Moran, E. & Kidd, M., 2001. Comparisons of male and female broiler responses to dietary threonine from 42 to 56 days of age. J. Appl. Poult. Res. 10, 53-59. [ Links ]
Eftekhari, A., Rezaeipour, V. & Abdullahpour, R., 2015. Effects of acidified drinking water on performance, carcass, immune response, jejunum morphology, and microbiota activity of broiler chickens fed diets containing graded levels of threonine. Livest. Sci. 180, 158-163. [ Links ]
Estalkhzir, F.M., Khojasteh, S. & Jafari, M., 2013. The effect of different levels of threonine on performance and carcass characteristics of broiler chickens. J. Nov. Appl. Sci. 2, 382-386. [ Links ]
Faure, M., Mettraux, C., Moennoz, D., Godin, J.P., Vuichoud, J., Rochat, F., Breuillé, D., Obled, C. & Corthésy-Theulaz, I., 2006. Specific amino acids increase mucin synthesis and microbiota in dextran sulfate sodium treated rats. J. Nutr. 136, 1558-1564. [ Links ]
Faure, M., Moënnoz, D., Montigon, F., Mettraux, C., Breuillé, D. & Ballèvre, O., 2005. Dietary threonine restriction specifically reduces intestinal mucin synthesis in rats. J. Nutr. 135, 486-491. [ Links ]
Garwood, V. & Diehl, K., 1987. Body volume and density of live Coturnix quail and associated genetic relationships. Poult. Sci. 66, 1264-1271. [ Links ]
Good, L., 2013., The effects of actigen® and threonine supplementation on growth parameters, immune function, and intestinal health in monogastrics. MS thesis, College of Agriculture, University of Kentucky, Lexington, Kentucky. [ Links ]
Gopinger, E., Xavier, E., Elias, M., Catalan, A., Castro, M., Nunes, A. & Roll, V., 2014. The effect of different dietary levels of canola meal on growth performance, nutrient digestibility, and gut morphology of broiler chickens. Poult. Sci. 93, 1130-1136. [ Links ]
Horn, N.L., Donkin, S.S., Applegate, T.J. & Adeola, O., 2009. Intestinal mucin dynamics: Response of broiler chicks and White Pekin ducklings to dietary threonine. Poult. Sci. 88, 1906-1914. [ Links ]
Jahanian, R., 2010. Effects of dietary threonine on performance and immunocompetence of starting broiler chicks. In 2nd International Veterinary Poultry Congress, Tehran, Iran. pp. 20-21. [ Links ]
Kerr, B. & Kidd, M., 1999. Amino acid supplementation of low-protein broiler diets: 1. Glutamic acid and indispensable amino acid supplementation. J. App. Poult. Res. 8(3), 298-309. [ Links ]
Kidd, M. & Kerr, B., 1997. Threonine responses in commercial broilers at 30 to 42 days. J. Appl. Poult. Res. 6(4), 362-367 [ Links ]
Kidd, M., Kerr, B. & Anthony, N., 1997. Dietary interactions between lysine and threonine in broilers. Poult. Sci. 76, 608-614. [ Links ]
Kidd, M., Lerner, S., Allard, J., Rao, S. & Halley, J., 1999. Threonine needs of finishing broilers: growth, carcass, and economic responses. J. Appl. Poult. Res. 8, 160-169. [ Links ]
Kidd, M., Pote, L. & Keirs, R., 2003. Lack of interaction between dietary threonine and Eimeria acervulina in chicks. J. Appl. Poult. Res. 12, 124-129. [ Links ]
Lehmann, D., Pack, M. & Jeroch, H., 1997. Effects of dietary threonine in starting, growing, and finishing turkey toms. Poult. Sci.76, 696-702. [ Links ]
Lemme, A., 2001. Responses of broilers to dietary threonine: A survey of the international literature. Amino News 2, 1-6. [ Links ]
Mack, S., Bercovici, D., De Groote, G., Leclercq, B., Lippens, M., Pack, M., Schutte, J. & Van Cauwenberghe, S., 1999. Ideal amino acid profile and dietary lysine specification for broiler chickens of 20 to 40 days of age. Br. Poult. Sci. 40, 257-265. [ Links ]
Mazraeh, A.A., Shahryar, H.A., Nobar, R.S.D. & Gorbani, A., 2013. The effects of different levels of L-threonine on growth performance and carcass traits of broiler chickens during phase growth. Bull. Env. Pharmacol. Life. Sci. 2(11), 122-124. [ Links ]
Mushtaq, T., Sarwar, M., Ahmad, G., Mirza, M., Ahmad, T., Noreen, U., Mushtaq, M. & Kamran, Z., 2009. Influence of sunflower meal based diets supplemented with exogenous enzyme and digestible lysine on performance, digestibility and carcass response of broiler chickens. Anim. Feed Sci. Technol. 149, 275-286. [ Links ]
NRC., 1994. Nutrient Requirements of Poultry. 9th revised edition. National Academy Press, Washington, DC. [ Links ]
Ojewola, G.S., Abasiekong, S.F. & Nwachukwu, C.S., 2001. Methionine supplementation in the productive efficiency, carcass characteristics and economics of growing indigenous turkey. Nigerian J. Anim. Sci. 4(2). [ Links ]
Ojano-Dirain, C. & Waldroup, P., 2002. Week of age under ivioderate temperature stress. Int. J. Poult. Sci. 1, 16-21. [ Links ]
Ospina-Rojas, I., Murakami, A., Oliveira, C. & Guerra, A., 2013. Supplemental glycine and threonine effects on performance, intestinal mucosa development, and nutrient utilization of growing broiler chickens. Poult. Sci. 92(10), 2724-2731. [ Links ]
Parsons, C.M., Potter, L.M. & Brown Jr, R.D., 1983. Effects of dietary carbohydrate and of intestinal microflora on excretion of endogenous amino acids by poultry. Poult. Sci. 62, 483-9. [ Links ]
Rezaeipour, V. & Gazani, S., 2014. Effects of feed form and feed particle size with dietary L-threonine supplementation on performance, carcass characteristics and blood biochemical parameters of broiler chickens. J. Anim. Sci. Technol. 56(1), 20. [ Links ]
Rezaeipour, V., Fononi, H. & Irani, M., 2012. Effects of dietary L-threonine and Saccharomyces cerevisiae on performance, intestinal morphology and immune response of broiler chickens. S. Afr. J. Anim. Sci. 42, 266-273. [ Links ]
Rosa, A., Pesti, G., Edwards, H. & Bakalli, R., 2001. Threonine requirements of different broiler genotypes. Poult. Sci. 80, 1710-1717. [ Links ]
Schaart, M.W., Schierbeek, H., Van der Schoor, S.R., Stoll, B., Burrin, D.G., Reeds, P.J. & Van Goudoever, J.B., 2005. Threonine utilization is high in the intestine of piglets. J. Nutr. 135, 765-770. [ Links ]
Smirnov, A., Tako, E., Ferket, P.R. & Uni, Z., 2006. Mucin gene expression and mucin content in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates. Poult. Sci. 85, 669-673. [ Links ]
Valizade, M.R., Sadeghi, A.A., Chamani, M., Shawrang, P. & Feizi, F., 2014. The Effect of increasing dietary threonine to lysine ratio on carcass characteristics, mucin gene expression and morphological analysis of ileum of male broiler chickens challenged with Salmonella. Int. J. Biosci. 5(11), 138-146. [ Links ]
Van der Schoor, S.R., Reeds, P.J., Stoll, B., Henry, J.F., Rosenberger, J.R., Burrin, D.G. & Van Goudoever, J.B., 2002. The high metabolic cost of a functional gut. Gastroenterology. 123, 1931-1940. [ Links ]
Wang, X., Qiao, S. Liu, M. & Ma, Y., 2006. Effects of graded levels of true ileal digestible threonine on performance, serum parameters and immune function of 10-25 kg pigs. Anim. Feed Sci. Technol. 129, 264-278. [ Links ]
Wang, W., Qiao, S. & Li, D., 2009. Amino acids and gut function. Amino acids 37, 105-110. [ Links ]
Zaefarian, F., Zaghari, M. & Shivazad, M., 2008. The threonine requirements and its effects on growth performance and gut morphology of broiler chicken fed different levels of protein. Int. J. Poult. Sci. 7, 1207-1215. [ Links ]
Received 8 March 2018
Accepted 12 July 2018
First published online 5 September 2018
# Corresponding author: drafzal@uvas.edu.pk


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}