










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.4 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i4.4
ARTICLES
Effects of air temperature on physiology and productive performance of pigs during growing and finishing phases
R.F. OliveiraI; R.H.R. MoreiraI; M.L.T. AbreuII; M.P. GionbelliII; A.O. TeixeiraIII; V.S. CantarelliII, #
IAnimal Science Department, Universidade Estadual Paulista, SP, 14.884-900, Brazil
IIAnimal Science Department, Universidade Federal de Lavras, Lavras, MG, 37.200-000, Brazil
IIIAnimal Science Department, Universidade Federal São João de Rei, MG, 36.307-352, Brazil
ABSTRACT
Thirty-six castrated male pigs were used to determine the influence of thermal environment and reduction of consumption on performance and carcass composition. Animals were housed in two climate chambers. In one, animals were in thermal comfort (TN) (22 °C), and in the other, pigs were under heat stress (HS) (34 °C). Animals were distributed in a randomized block design, making three treatments (TN, HS and animals in thermal comfort with food consumption paired with that observed in HS (PFTN)), with six replicates and two animals per experimental unit. Data were obtained on performance and carcass composition. The weight gains of HS and PFTN animals were reduced by 40.5% and 34.7%, respectively, reflecting a reduction of 13.2% in the final weight of PFTN animals. Triiodothyronine concentration was not affected by heat, but there was an increase in lymphocyte numbers in PFTN animals. The HS and PFTN animals showed lower hot carcass weight. However, there were no effects on hot carcass yield and relative weights of heart, lung and spleen. Heat stress compromised performance. The negative effects of high temperature on pigs include reduction in feed intake and changes in physiology.
Keywords: Environment, heat stress, pair feed, pig growth, pig production
Introduction
In recent years, there has been a global increase in environmental temperatures. This has affected pig production since, in heat stress, there is a need for adjustment to maintain homeothermia. To survive in hot environments, the pig makes physiological adjustments (Kiefer et al., 2010; Soerensen & Pedersen, 2015), resulting in alterations in blood parameters (Dong et al., 2012; Pearce et al., 2013; Dantas et al., 2014; Haoaet al., 2014). Associated with this, there is greater thermogenesis owing to the high protein deposition in the carcass (White et al., 2008), which may compromise performance even more.
In a study conducted with growing pigs kept in heat (32 °C) for two weeks, Fernandez et al. (2015) verified changes in carbohydrate and lipid metabolism. In another study, also with growing animals kept in heat (30 °C), there were hormonal changes, with a significant reduction in circulating insulin (Mayorga et al., 2017). Similar results with finishing pigs were obtained by Haoa et al. (2014), who verified that the deleterious effects in performance were caused not only by the reduction in consumption, but also by enzymatic changes in the blood and in the muscles of the animals. The isolated results of the effect of the thermal environment were obtained with the adoption of the paired feeding technique. Thus, metabolic changes in pigs may not be due solely to the lower intake of nutrients in the warm environment. However, the responses were obtained in studies conducted for short periods (2 to 3 weeks). In view of these aspects, the objective of this work was to determine whether the worst performance of the pigs kept in heat is due only to the reduction in the nutrient intake or whether they suffer the consequences of physiological changes in this hostile environment during the growth and termination phases.
Material and Methods
The use of experimental animals was approved by CEUA (Ethics Committee of Animals Use) of the Universidade Federal de Lavras (UFLA) with protocol number 059/14.
Thirty-six commercial hybrid barrows with high genetic potential were used. Their mean initial weight was 30.5 ± 0.6 kg and the mean age, 77 ± 2 days. Animals were housed in two climate chambers in which temperature, humidity and ventilation could be controlled. Each chamber had 12 experimental pens with concrete floors, 2.3 x 1.5 m, equipped with semi-automatic feeders and nipple drinkers. In one of the chambers, animals were in thermal comfort (22 °C) and in the other pigs were in heat stress (34 °C). The three treatments consisted of HS: animals in heat stress (34°C); TN: animals in thermal comfort (22 °C); and PFTN: animals in thermal comfort (22 °C) with food consumption paired with that observed in HS. Twelve animals per treatment were distributed in six replicates, an experimental unit being composed of two animals in the pen. Thus, one of the chambers housed 12 pigs (comfort) plus 12 of the PFTN treatment, and the other chamber 12 pigs of the HS treatment. Pigs were distributed in randomized blocks, creating three treatments with six replications. The initial weight of animals was used as a criterion for forming the blocks.
Pigs were weighed and distributed to their experimental units simultaneously. However, to enable pairing, data collection in PFTN animals was started 24 hours after the experiment began, so that all pigs remained 71 days in the experiment (43 days in the growing phase and 28 days in the finishing phase). The feed intake of HS was calculated and the same amount was provided to PFTN animals the next day. This procedure was repeated every day during the experiment. Consequently, every day the diet provided, leftovers and waste were measured in all experimental units.
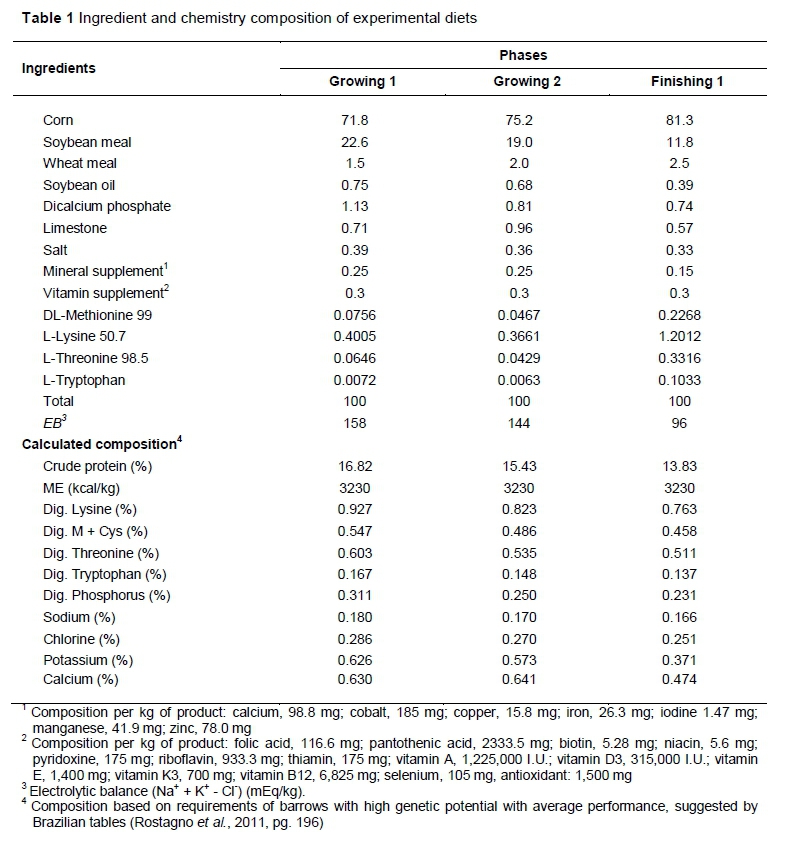
Experimental diets (Table 1) were iso-energetic and formulated based on corn and soybean meal, supplemented with vitamins and minerals to meet the minimum requirements suggested by the Brazilian tables (Rostagno et al., 2011). The authors applied the recommendations for barrows with high genetic potential with average performance of 30-50 kg (growing 1), 50-70 kg (growing 2) and 70-100 kg (finishing).
The environmental conditions of the climatic chambers were monitored throughout the trial period by digital thermohygrometer (Model7666.02.0.00, Cotronic Technology Ltd, Incoterms, China), with an accuracy of 1 °C ± 5%. Relative humidity (RH) with internal and external sensors coupled to the black globe. The equipment was maintained at various points of the experimental units at half the animals' body height and the collected data were used to calculate the globe temperature and humidity index (BGHI).
At the end of each phase (growing and finishing), after weighing, blood was sampled from animals of each experimental unit via jugular vein puncture. Animals were randomly chosen and in the next phase collection was repeated on the same animal. After standing for about 90 minutes to decant, blood samples were centrifuged at 3000 rpm for 10 minutes to remove the serum, which was stored at -20 °C. After this process, serum levels of total triiodothyronine hormone were obtained (total T3) from kits using chemiluminescence in AXES equipment. An aliquot of this material was used to measure neutrophil (N) and lymphocyte (L) concentrations to calculate the N/L ratio and for blood chemistry analyses to determine glucose, urea and creatinine levels, which helped to characterize the heat stress status.
The supplied feed, leftovers and waste were considered to determine the average daily feed intake (ADFI). Animals were weighed at the beginning and end of each phase to establish the average daily gain (ADG). Feed conversion was obtained from the ratio between ADFI and ADG. Digestible lysine consumption (DLC) was obtained from the ADFI considering the lysine concentration in the diet. Based on DLC, the authors determined the lysine use efficiency for weight gain (LUEG), which was calculated by considering the weight gain obtained per gram of digestible lysine intake during the experiment. After the finishing phase, backfat thickness (BFT), loin depth (LD) and ribeye area (REA) of pigs were measured. For this, the Aloka ultrasound equipment (SSD-500 model) and a linear transducer of 3.5 MHz (UST 5011 model) with accuracy of 92.1%, were used according to Souza's (2011) methodology. The measurements were taken at 6.5 cm from the midline on the left side of the animal around the last rib (P2 point).The final weights of the piglets in the growth, consumption and feed conversion phases were used as covariates to analyse the performance of piglets in the finishing phase in these same parameters.
At the end of the experimental period, animals were subjected to 12 hours fasting, and then weighed. All animals were moved to a commercial slaughterhouse and slaughtered by electrical stunning and bleeding. Subsequently, the carcass was scalded, dehaired, cleaned and opened for evisceration. At the end of the slaughter line, carcasses were weighed to determine the hot carcass weight (HCW) and hot carcass yield (HCY). Carcasses were divided along the middle and weighed. For carcass traits, HCY, length (CL), hot HCW, and the pH at 45 minutes (pH45min) were evaluated. The organs (heart, lungs, liver, kidneys, and spleen) were washed and hung for 15 minutes in the shade to drain water and blood for subsequently weighing.
The data obtained from parameters, blood analyses, performance and carcass traits were tested to verify normality by the Lilliefors test. Then, the data were submitted to analysis of variance by F test at 5% probability, using SAS (SAS System, Cary, NC, 2000). Once significance had been verified, the authors used the Tukey test at 5% probability. The final weight of the growing phase was used as a covariate for the finishing phase.
Results
Values of air temperature (air T), relative humidity (RH) and globe temperature and humidity index (BGHI) of acclimatized rooms are presented in Table 2.
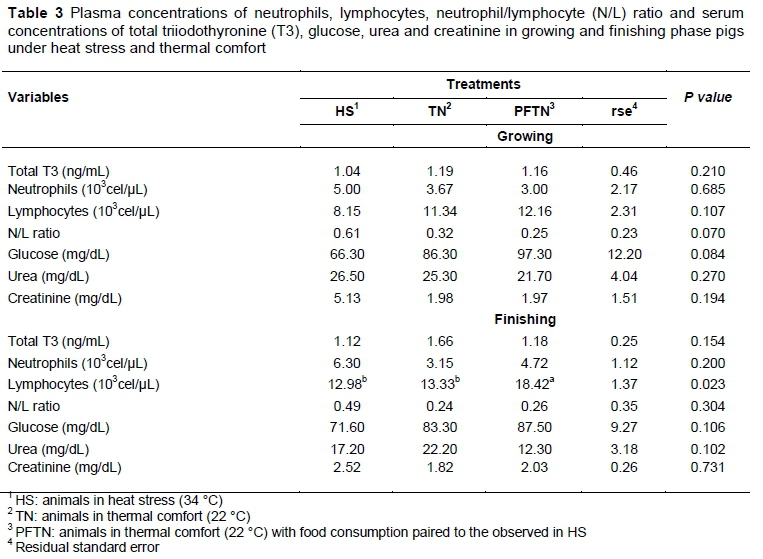
Table 3 shows the plasma concentrations of neutrophils, lymphocytes, neutrophil/lymphocyte (N/L) ratios, and serum concentrations of total T3, glucose, urea and creatinine in pigs during the growing and finishing phases.
During the growing phase, there were no effects (P >0.050) of treatments on plasma concentrations of neutrophils, lymphocytes, N/L ratio and serum total T3, glucose, urea and creatinine. In the finishing phase, there were no effects (P >0.050) of treatments on concentrations of total T3, neutrophils and glucose, urea and creatinine. However, there was an increase (P =0.023) in lymphocyte concentration of PFTN animals.
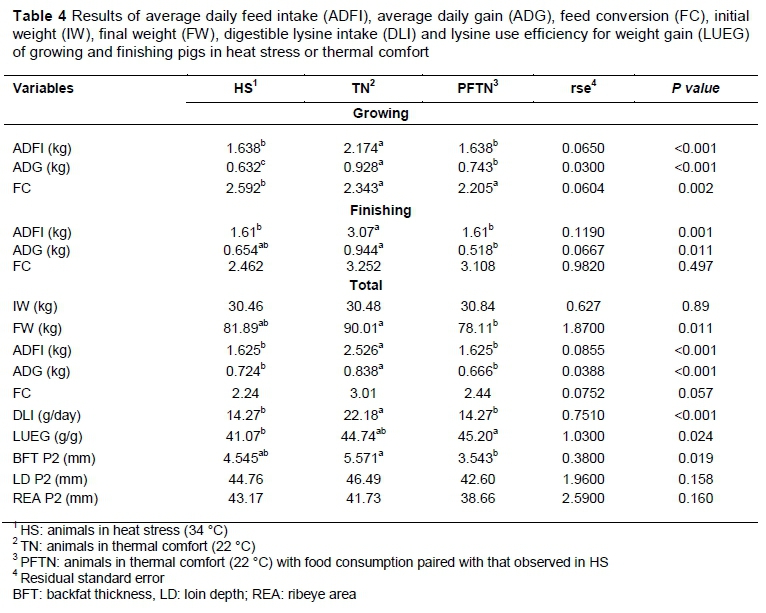
Performance values in the growing and finishing phases and in the total period of the experiment are presented in Table 4.
It was observed that heat stress of 34 °C promoted reduction (P <0.050) of ADG and ADFI in the growing and finishing phases. In the growing phase, animals under heat stress from HS treatment showed a reduction in ADFI of 24.7% compared with TN animals. In the total period of the experiment, dietary restriction negatively influenced (P =0.011) the final weight of pigs, with a reduction of 13.2% compared with TN. In this period, HS and PFTN pigs had lower ADFI (P <0.001), decreasing by 35.6% compared with TN. Similar behaviour was observed in ADG. For this trait, HS and PFTN pigs showed reductions (P <0.001) of 40.5% and 34.7%, respectively, compared with TN.
Digestible lysine intake (DLI) had a similar response to ADFI. The HS and PFTN pigs showed lower DLI (P <0.001) than TN animals in the total period. The feed intake of HS was calculated and the same amount was provided to PFTN animals the next day. This procedure was repeated every day during the experiment. On the other hand, PFTN animals had higher LUEG (P =0.024) than animals under heat stress, being 10% higher compared with HS animals. Food restriction promoted a reduction in backfat thickness (BFT) (P =0.019), but did not influence LD (P =0.158) and REA (P =0.160).
Animals under thermal comfort (TN) had higher BFT compared with PFTN. It is possible that the lower energy consumption promoted by ADFI restriction of PFTN animals, is responsible for BFT decrease.
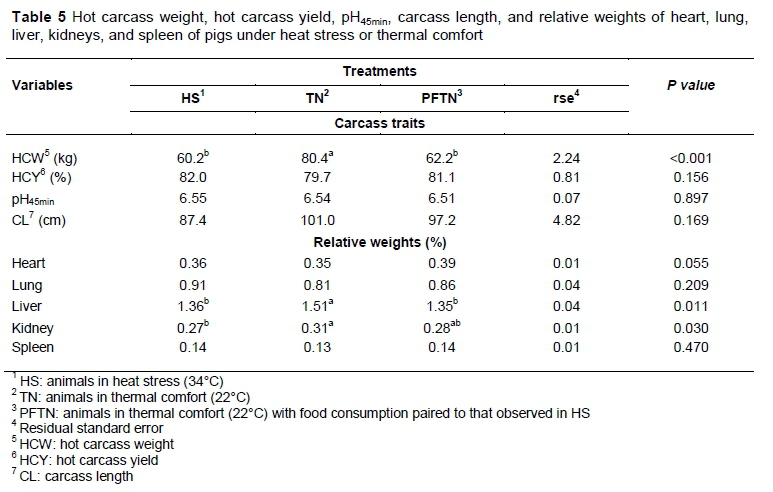
Carcass traits and relative weights of the organs (heart, lung, liver, kidney and spleen) are presented in Table 5. Heat stress or food restriction promoted lower (P <0.01) HCW. However, there were no treatment effects (P >0.05) on HCY, initial pH and CL, or on relative weight of heart, lung and spleen. However, TN animals had greater relative weight of liver (P =0.011), compared with other treatments, and of kidney (P =0.030), compared with HS.
Discussion
Despite the intensity of heat stress, there was no reduction in total T3 concentrations in growing and finishing pigs. The results of this study do not agree with frequent reports in the literature by various authors that confirmed the existence of an environmental influence on serum and plasma T3 (Oliveira & Donzele, 1999; Patience et al., 2005). Batista et al. (2011) also reported that the reduction in the blood concentration of T3 in the heat is justified because the thyroid hormones are involved in the metabolic activity of the animals and, consequently, in the production of body heat. There is no biological explanation for this, since in the heat the concentration of T3 was reduced in the blood circulation.
Although the haematological parameters of growing and finishing pigs were not affected by heat, the restriction of feed intake of PFTN animals caused lymphocytosis. This increase in lymphocytes was not enough to influence the N/L ratio. Possibly, it was caused by increased adrenaline secretion stimulated by the autonomic nervous system (Marzzoco & Torres, 2007), because of the restriction of the food supply. Adrenaline promotes lymphocyte mobilization into the general circulation, increasing the total count, and causing physiological lymphocytosis (González & Silva, 2008).
The animals kept in the heat did not present variations in glycaemia. The current results differ from those reported by Dantas et al. (2014), who observed lower values of glucose in animals maintained in heat.
Although heat stress did not influence urea concentration, only TN treatment reached the reference values proposed by Kaneko et al. (2008), in that PFTN animals in growing phase and animals from HS and PFTN in the finishing phase showed lower values. These results are similar to those observed by Freitag et al. (2014), who evaluated the effect of crude protein reduction on blood parameters of pigs in the initial growing phase under heat stress (28.8 °C) and comfort temperatures (18.1 °C), in which no differences were observed in plasma concentrations of urea. In addition, creatinine values were considered normal for growing and terminating animals (Unger et al., 2007; Kaneko et al., 2008). These results differ from those obtained by Freitag et al. (2014), who observed an increase in creatinine concentration in animals subjected to heat stress.
The lowest ADG observed in HS animals may be explained by ADFI reduction, in that it was lower compared with TN. That is, the gain followed consumption. Pigs exposed to temperatures above the upper critical (27 °C) level reduce food consumption (and therefore nutrients), attempting to decrease the metabolic rate to reduce body temperature, which causes performance reduction. Another factor would be energy expenditure, aimed at heat dissipation, resulting in an increase in days to reach slaughter weight. These results are consistent with studies from Manno et al. (2006) and Kiefer et al. (2009; 2010), who showed that the performance of pigs in the growing phase decreased under heat stress.
Similarly, the ADFI of PFTN animals in the growing and finishing phases resulted in food restrictions of 24.7% and 47.6%, respectively, compared with TN animals. Despite similar consumption, the ADGs of HS and PFTN pigs were different in the growing phase, in which PFTN animals presented a higher performance. This can be only partly explained by the intensity of heat stress on animals, as according to Collin et al. (2001), the negative effect of high temperatures on pig ADG is not explained solely by feed intake reduction. These results were similar to those obtained by Manno et al. (2006) and Morales et al. (2014), in which pairfeeding of pigs under thermal comfort and others under heat stress during the growing phase showed that the effect of high temperature would not be restricted to reduction in consumption. Additionally, Quiniou et al. (2000), who studied the effect of environment, observed that room temperature was related to animal feed intake and body weight.
Animals subjected to a high temperature environment (HS) presented feed conversion differences compared with those in a comfort environment during the growing phase, regardless of the feeding programme (TN or PFTN). The proportional variation of ADFI and ADG of these treatments may explain the current results, which differ from those obtained by Manno et al. (2006), who observed that during the growing phase, animals exposed to high temperature showed better feed efficiency (2.08) than pigs under thermal comfort, fed ad libitum (2.22) or paired (2.23).
Renaudeau et al. (2011), in a study with growing pigs, observed lower feed conversion values in animals under heat stress. On the other hand, Rodrigues et al. (2012) observed no differences for feed conversion in the finishing phase of pigs in thermoneutral or high temperature environments. These results could be associated with lower efficiency of pigs at high temperature in using the energy for growing (Renaudeau et al., 2008) in that part of the nutrients are diverted to heat dissipation.
A similar variation was observed between DLI and feed intake. However, HS animals were less efficient than PFTN, indicating a possible effect of environment temperature on protein metabolism, in that greater gluconeogenesis intensity for energy is observed in pigs under heat stress conditions (Pearce et al., 2013). Moreover, lysine, which is the first limiting amino acid, is ketogenic (Liao et al., 2015).
The intense heat stress associated with increased energy expenditure for maintenance, owing to the increased heat loss, may have compromised the LUEG in animals subjected to HS.
These results differ from those reported by Collin et al. (2001), in which they observed that pigs under heat stress are metabolically more efficient in synthesis processes, reducing the protein turnover. Manno et al. (2006) observed better LUEG in animals subjected to heat stress.
The HS and PFTN animals had lighter carcasses. These results are in agreement with those obtained by Manno et al. (2006) and Kiefer et al. (2009; 2010), who observed lighter carcass weight in pigs under heat stress. Manno et al. (2006) observed that growing pigs in thermal comfort with paired feeding with pigs under heat stress had lighter carcasses compared with animals under heat stress and thermal comfort fed ad libitum.
Although animals kept in TN presented heavier HCW, there was no effect of treatments on HCY. This was probably because the relative weights of viscera, which were unaffected by treatments, with the exception of liver and kidney weight. The reduction in relative weights of viscera and organs caused by high temperature was expected in this study, owing to the high expenditure rate of energy relative to animal size, which would have a substantial impact on animal maintenance requirements (Renaudeau et al., 2013). These results were similar to those obtained by Ferreira et al. (2007) and Orlando et al. (2007), which, in a study with growing pigs under heat environment, observed no significant effect of CP levels on the relative weight of the evaluated organs. Rinaldo et al. (2000) who studied the effect of a tropical climate on pig performance during the initial stages of growing, observed that are reduction of liver and kidneys is most associated with a reduction of voluntary consumption and the generation of internal heat, which are similar results to those observed in this study.
These results are in agreement with those obtained by Manno et al. (2006), who observed carcass composition changes, depending on the feeding restriction of pigs subjected to thermal comfort, which resulted in reduced daily deposition of protein and fat in their carcasses compared with animals fed ad libitum. In contrast, Pearce et al. (2013), in a study with growing pigs, observed higher BFT in pig carcasses subjected to heat stress and ad libitum feeding compared with animals under thermal comfort receiving ad libitum and paired feeding with consumption of animals under heat stress.
Conclusion
The heat stress of 34 °C changed the physiology of growing and finishing pigs, adversely affecting their performance. Deleterious effects of heat stress in pigs are not based only on feed intake reduction.
Acknowledgments
This study was supported by the Federal University of Lavras - Minas Gerais / Brazil. Animal Nutri provided the animals and ingredients used in the experimental ration.
Authors' Contributions
RFO, MLTA, MPG, AOT, VSC and RAF conceived, designed, and analysed the experiment; RFO and RHRM supervised animal management; RFO carried out laboratory analysis. All authors drafted, read, and approved the final manuscript.
Conflict of interest declaration
The authors of this article state that there is no conflict of interest between the other authors and with any person or institution outside of the publication.
References
Batista, R.M., Oliveira, R.F.M., Donzele, J.L., Oliveira, W.P., Lima, A.L. & Abreu, M.L.T., 2011. Lisina digestível para suínos machos castrados de alta deposição de carne submetidos a estresse por calor dos 30 aos 60 kg. R. Bras. Zootec. 40, 1925-1932. [ Links ]
Collin, A., VanMilgan, J., Dubois, S. & Noblet, J., 2001. Effect of high temperature and feeding level on energy utilization in piglets. J. Anim. Sci. 79, 1849-1857. [ Links ]
Dantas, W.M.F., Ribeiro Filho, J.D., Guimaráes, J.D. & Guimaráes, S.E.F., 2014. Perfil metabólico e ponderal de leitões submetidos a dietas com diferentes teores de fósforo em ambientes térmicos distintos. Pesq. Agropec. Bras. 49, 901-910. [ Links ]
Dong, H., Zhong, Y.G., Liu, F.H., Yang, K., Yu, J. & Xu, J.Q., 2012. Regulating effects and mechanisms of Chinese medicine decoction on growth and gut hormone expression in heat stressed pigs. Livest. Sci.143, 77-84. [ Links ]
Fernandez, M.V.S., Johnson, J.S., Abuajamieh, M., Stoakes, S.K., Seibert, J.T., Cox, L.& Rhoads, R.P., 2015. Effects of heat stress on carbohydrate and lipid metabolism in growing pigs. Physiol. Rep. 3, 12315-12332. [ Links ]
Ferreira, R.A., Oliveira, R.F.M., Donzele, J.L. & Saraiva, E.P., 2007. Redução da proteína bruta e suplementação de aminoácidos para suínos machos castrados dos 30 aos 60 kg mantidos em ambiente de alta temperatura. R. Bras. Zootec. 36, 818-824. [ Links ]
Freitag, D.C., Klosowsket, E.S., Pozza, P.C., Oliveira, A.C., Tsutsumi, C.Y., Nunes, R.V. & Sangali, C.P., 2014. Redução da proteína bruta em rações sobre os balancos metabólicos de suínos mantidos em diferentes condicöes térmicas. Semina: Ciencias Biológicas e da Saúde. Londrina, 35, 61-70. [ Links ]
González, F.H.D. & Silva, S.C., 2008. Patología clínica veterinária: texto introdutório. Porto Alegre: Universidade Federal do Rio Grande do Sul. [ Links ]
Haoa, Y., Fenga, Y., Yanga, P., Fenga, J., Linb, H. & Gua, X., 2014. Nutritional and physiological responses of finishing pigs exposed to a permanent heat exposure during three weeks. Arch. Anim. Nutr. 68, 296-308. [ Links ]
Kaneko, J.J., Harvey, J.W. & Bruss, M., 2008. Clinical biochemistry of domestic animals. 5th ed. Academic Press, San Diego. [ Links ]
Kiefer, C., Meignen, B.C.G., Sanches, J.F. & Carrijo, A.S., 2009. Resposta de suínos em crescimento mantidos em diferentes temperaturas. Arch. Zootec. 58, 55-64. [ Links ]
Kiefer, C., Moura, M.S., Silva, E.A., Santos, A.P., Silva, C.M., Luz, M.F. & Nantes, C.L., 2010. Respostas de suínos em terminacáo mantidos em diferentes ambientes térmicos. Rev. Bras. Saúde Prod. Anim. 11, 496-504. [ Links ]
Liao, S.F., Wang, T. & Regmi, N., 2015. Lysine nutrition in swine and the related monogastric animals: Muscle protein biosynthesis and beyond. Springer Plus 4, 147. [ Links ]
Manno, M.C., Oliveira, R.F.M., Donzele, J.L., Oliveira, W.P., Vaz, R.G.M.V., Silva, B.A.N., Saraiva, E.P. & Lima, K.R.S. 2006. Efeitos da temperatura ambiente sobre o desempenho de suínos dos 30 aos 60 kg. Rev. Bras. Zootec. 35, 471-477. [ Links ]
Marzzoco, A. & Torres, B.B., 2007. Bioquímica básica. 3rd edition. Guanabara Koogan, Rio de Janeiro. [ Links ]
Mayorga, E.J., Kvidera, S.K., Horst, E.A., Al-Qaisi, M.A., Dickson, M.J., Seibert, J.T., Lei, S., Rambo, Z.J., Wilson, M.E. & Baumgard, L.H., 2017. Effects of zinc amino acid complex on biomarkers of gut integrity and metabolism during heat stress and a following recovery period in growing pigs. J. Anim. Sci. 95(Suppl2), 158-158. Doi:10.2527/asasmw.2017.325. [ Links ]
Morales, A., Grageola, F., García, H., Arce, N., Araiza, B., Yáñez, J. & Cervantes, M., 2014. Performance, serum amino acid concentrations and expression of selected genes in pair-fed growing pigs exposed to high ambient temperatures. J. Anim. Physiol. nim. Nutr. 98, 928-935. [ Links ]
Oliveira, R.F.M. & Donzele, J.L., 1999. Effect of environmental temperature on performance and on physiological and hormonal parameters of gilts fed at different levels of digestible energy. Anim. Feed. Sci. Technol. 81, 319-331. [ Links ]
Orlando, U.A.D., Oliveira, R.F.M., Donzele, J.L. & Silva, F.C.O., 2007. Níveis de proteína bruta e suplementação de aminoácidos em rações para leitoas dos 30 aos 60 kg mantidas em ambiente de alta temperatura. Rev. Bras. Zootec. 36, 1573-1578. [ Links ]
Patience, J.F., Umboh, J.F., Chaplin, R.K. & Nyachoti, C.M., 2005. Nutritional and physiological responses of growing pigs exposed to a diurnal pattern of heat stress. Livest. Prod. Sci.96, 205-214. [ Links ]
Pearce, S.C., Gabler, N.K., Ross, J.W., Escobar, J., Patience, J.F., Rhoads, R.P. & Baumgard, L.H., 2013. The effects of heat stress and plane of nutrition on metabolism in growing pigs. J. Anim. Sci. 91, 2108-2118. [ Links ]
Quiniou, N., Dubois, S. & Noblet, J., 2000. Voluntary feed intake and feeding behaviour of group-housed growing pigs are affected by ambient temperature and body weight. Livest. Prod. Sci. 63, 245-253. [ Links ]
Renaudeau, D. Gourdine, J.L. & St-Pierre, N.R., 2011. A meta-analysis of the effect of high ambient temperature on growing-finishing pigs. J. Anim. Sci. 89, 2220-2230. [ Links ]
Renaudeau, D. Kerdoncuff, M. Anais, C. & Gourdine, J.L., 2008. Effect of temperature level on thermal acclimation in Large White growing pigs. Animal 1619-1626. [ Links ]
Renaudeau, D., Frances, G., Dubois, S., Gilbert, H. & Noblet, J., 2013. Effect of thermal heat stress on energy utilization in two lines of pigs divergently selected for residual feed intake. J. Anim. Sci. 91, 1162-1175. [ Links ]
Rinaldo, D., Dividich, J. & Noblet, J., 2000. Adverse effects of tropical climate on voluntary feed intake and performance of growing pigs. Livest. Prod. Sci. 66, 223-234. [ Links ]
Rodrigues, N.E.B., Fialho, E.T., Zangeronimo, M.G., Cantarelli, V.S., Rodrigues, P.B., Rodrigues Filho, M., Gomide, E.M. & Betarelli, R.P., 2012. Reduction in the protein level and addition of oil in diets for finishing pigs under different temperatures. Rev. Bras. Zootec. 41, 1878-1883. [ Links ]
Rostagno, H.S., Albino, L.F.T., Donzele, J.L., Gomes, P.Z., Oliveira, R.F., Lopes, D.C., Ferreira, A.S., Barreto, S.L.T. & Euclides, R.F., 2011. Tabelas brasileiras para aves e suínos: composição de alimentos e exigencias nutricionais. 3rd edition. Vicosa,UFV, Departamento de Zootecnia. [ Links ]
Soerensen, D. & Pedersen, L., 2015. Infrared skin temperature measurements for monitoring health in pigs: A review. Acta Vet. Scand. 57, 5-16. [ Links ]
Souza, G.H.C., 2011.Níveis de ractopamina em dietas para suínos em terminacáo. Dissertation, Universidade Federal dos Vales do Jequitinhonha e Mucuri, Diamantina. [ Links ]
Unger, V., Grosse-Siestrup, C., Fehrenberg, C., Fischer, A., Meissler, M. & Groneberg, D.A., 2007. Reference values and physiological characterization of a specific isolated pig kidney perfusion model. J. Occup. Med. Toxicol. 2, 1-13. [ Links ]
White, H.M., Richert, B.T., Schinckel, A.P., Burgess, J.R., Donkin, S.S. & Latour, M.A., 2008. Effects of temperature stress on growth performance and bacon quality in grow-finish pigs housed at two densities. J. Anim. Sci. 86, 1789-1798. [ Links ]
Received 7 August 2017
Accepted 15 January 2018
First published online 7 July 2018
# Corresponding author: rony@dzo.ufla.br


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}