










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.3 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i3.13
ARTICLES
In vitro ruminal fermentation and fatty acid production by various oil seeds
P. KubelkováI, #; D. JalčII; F. JančíkI; P. HomolkaI
IDepartment of Nutrition and Feeding of Farm Animals, Institute of Animal Science, av. Přátelství 815,104 00 Prague-Uhříněves, Czech Republic
IIInstitute of Animal Physiology, Slovak Academy of Sciences, av. Soltésovej 4-6, 040 01 Kosice, Slovak Republic
ABSTRACT
Rumen simulating techniques (Rusitec) were used to determine the impact of diets containing milled oilseeds on the fermentation parameters and amount of fatty acids (FA) in the effluent. High-forage diets containing no oilseeds (control diet (CD)) or 10% oilseed meal from rapeseed (RS), sunflower seed (SS), or flaxseed (FS) were used on a dry matter (DM) basis. No differences in DM digestibility were observed between the diets. Inclusion of SS and FS significantly reduced the pH values of the ruminal fluid, and a significant decline in the ammonia nitrogen (NH3-N) (mg/d) production in effluent was observed in the vessels with SS. Generally, oilseeds in these diets significantly reduced the amount of total fermentation gases (L/d); however, only a tendency toward methane (CH4, %) decrease was detected. The addition of oilseeds also significantly diminished the amount of total volatile fatty acids (VFA) produced (mmol/d). Significant reductions in the amounts of saturated FA in the vessels with RS and FS were observed compared with the CD and a significantly higher amount of monounsaturated fatty acids (MUFA) was noted in the vessels with RS. An increased amount of polyunsaturated fatty acids (PUFA), compared with the CD, was statistically significant only in the vessels with FS.
Keywords: Flaxseed, high-forage diet, methane, rapeseed, sunflower seed
Introduction
Fat supplements such as oilseeds are included in the diets of ruminants to increase energy density, improve nutrient utilization, enhance milk and meat yields, and modify fatty acid (FA) composition (Soder et al., 2013). Dietary fats can modify the ruminal microbial population responsible for cellulose digestion (Getachew et al., 2001), reducing the rumen degradation of fibre and organic matter (Machmüller et al., 2006). Nevertheless, some experiments showed no effect (Beauchemin et al., 2007) and even positive responses (Sinclair et al., 2005) after the addition of oils to the diets. Controversial results may be due not only to the type and amount of oil, but also to the basal diet composition.
The addition of unsaturated fat sources decreased the concentration of ammonia nitrogen (NH3-N) in the rumen (Egan et al., 1986) and enhanced the effectiveness of microbial nitrogen synthesis (Oldick & Firkins, 2000). The addition of fat to the ruminant rations appears to be an efficient and easy way of reducing methane (CH4) production (Boadi et al., 2004). Fats with high amounts of long-chain unsaturated fatty acids, such as sunflower and rapeseed oils, depressed the emissions of CH4 by approximately 22% of the total energy received by cows fed diets with high contents of forage (McGinn et al., 2004; Beauchemin & McGinn, 2006). However, it has been reported that fats do not have any specific effect on CH4 release, and that the evident decrease reflects a fall in the digestibility of the ration nutrients (Beauchemin et al., 2007).
Based on these findings, the authors hypothesized that the addition of milled oilseeds to the high-forage diet is not necessarily associated with detrimental effects on ruminal fermentation. The objectives of the present study were therefore to determine the impact of feeding high-forage diets containing milled oilseeds on the fermentation parameters and amount of FAs in effluents in an in vitro experiment.
Materials and Methods
The incubation was performed using Rumen simulating techniques (Rusitec) with a unit consisting of four vessels with a nominal volume of 850 ml each (Czerkawski & Breckenridge, 1977). The control diet (CD) included 70% cut meadow hay (from 10 to 12 mm long) and 30% barley meal (ground through a 1 mm screen sieve) on a dry matter (DM) basis. Experimental diets included 70% cut meadow hay, 20% barley meal and 10% oilseed meal - rapeseeds (RS), sunflower seeds (SS) or flax seeds (FS) on a DM basis. Seeds were ground through a 1-mm screen sieve. The flow through fermenters was maintained by continuous infusion of McDougall's solution of artificial saliva supplemented with microelements (mg/L): zinc sulfate (ZnSO), 1.92; magnesium sulfate (MnSO4), 1.02; cobalt sulfate (CoSO4), 0.06; at pH 8.4; (McDougall, 1948) at a rate of 610 mL/d (dilution rate of 3.0%/h; Dohme et al., 2000). With the dietary crude protein (CP) contents ranging from 90.0 g/kg for the CD to 111.0 g/kg (on DM basis) for the diets containing oilseeds, 283.2 (CD), 85.0 (RS diet), 160.5 (SS diet), and 122.8 (FS diet) mg of urea per 1 L of McDougall's solution of artificial saliva was added to balance the CP contents in the respective vessels to 12%.
The duration of the experimental period was 12 days, with the first six days being used for equilibration and the last six days for sample collection. Ruminal fluid used as inoculum was obtained from two ruminally fistulated Slovak merino sheep (mean bodyweight 42.1 + 2.0 kg) fed on a diet consisting of 70% meadow hay and 30% barley meal. Sheep were housed in individual pens with free access to water and feed. The experimental protocol was approved by the Ethical Committee of The Institute of Animal Physiology Slovak Academy of Sciences and State Veterinary and Food Office (Ro-2762/05-221/3). Rumen content (solid and liquid) was collected through a ruminal cannula one hour before the morning feeding and transferred to the laboratory under anaerobic conditions at 39 °C. Each reaction vessel was filled with 450 ml rumen fluid and 400 ml artificial saliva. Squeezed particulate rumen contents (100 g) were weighed into a nylon bag (pore size 100 μπι), which was placed inside the feed container in each vessel as a donor of bacteria during the first 24 hours. Each vessel contained two bags of the same substrate (introduced on two consecutive days), which remained for 48 hours in the fermenters. The fermenters were purged with nitrogen (N2) after manipulation to maintain anaerobic conditions, and the temperature in vessels was maintained at 39 °C. During sampling (days 7-12), the effluent collection flasks were cooled to 2 °C in a refrigerated water bath to inhibit microbial growth and fermentation.
Fluid was sampled daily from each vessel before the bags were replaced and the pH (Inolab Level 1, WTW, Weilheim, Germany) was measured immediately. (The pH values on days 7-12 were used for statistical analysis). The gas produced was collected daily on days 7-12 in new rubber bags and analysed for total gas volume and CH4 concentration. The liquid effluents in the flasks were also collected daily, and on days 7-12 were analysed for volatile fatty acids (VFA), FA and NH3-N production. The disappearance of DM after 48 hours incubation was determined from the weight loss after oven drying at 55 °C for 48 hours, and the residues were then analysed for neutral detergent fiber (NDF) (Van Soest et al., 1991) in the presence of sodium sulphite without α-amylase treatment and the results presented as ash free. The acid detergent fiber (ADF) (AOAC Official Method 973.18, AOAC, 2005), CP (Kjeldahl method), ether extract (EE), and ash (AOAC, 2005) analyses were done on the diets and in the residues to allow for the determination of nutrient digestibility. The VFA concentrations in the effluents were determined by gas chromatography (Cottyn & Boucque, 1968) in a Perkin-Elmer 8500 gas chromatograph using crotonic acid as an internal standard. The NH3-N concentrations were measured by the microdiffusion method (Conway, 1962), gas volumes were determined with a flow gasometer, and CH4 was detected in a Perkin-Elmer 8500 gas chromatograph as described by Czerkawski & Clapperton (1984).
The lipids from the diets and effluents were extracted and methylated according to the method of Folch et al. (1957). The fatty acid methyl ester profiles were determined by gas chromatography on a 6890N chromatograph (Agilent Technologies) with a G1315A autosampler. A 60 x 0.25-mm x 0.25-μΓΠ i.d. fused silica capillary column (DB-23, Agilent Technologies) was used. The N2 was applied as the carrier gas at 0.8 ml/min and as the makeup gas at 30 ml/min. The temperatures of the injector and the flame ionization detector were 230 °C and 260 °C, respectively. A timed-temperature program was used with the following settings: i) an initial oven temperature of 120 °C with a hold for 6 min; ii) an increase of 15 °C/min to 170 °C; iii) an increase of 3 °C/min to 210 °C, followed by a hold for 13.5 min; and iv) an increase of 40 °C/min to 230 °C, with a final hold for 7 min. The total run time was 44 min. The split ratio was 1:1 for the effluents and 1:40 for the feeds, with 1 μ! injected. The total run flow rates of air and hydrogen were 300 ml/min and 30 ml/min, respectively. Data were extracted with a GC ChemStation B.01.01.
The data were analysed statistically using the general linear model (GLM) of SAS Institute, Inc. (2000) for a completely random design with the following model:
Y = μ + T + e,j,
Where: Y is the dependent variable
μ is the overall mean,
T is the fixed effect of the dietary treatments (i = 1-4)
ej is the residual effect
When differences among the vessels were significant at the 0.05 level, Tukey's post hoc test was used to compare the means among treatments. The normality of values was evaluated with the Shapiro-Wilk test (SAS Institute, 2000). The results in the tables are presented as the means and the standard errors of means (SEM).
Results and Discussion
The chemical composition of the components and the diets are shown in Table 1. The oilseeds contained from 182 g/kg to 225 g/kg (on a DM basis) CP compared with 118 g/kg in barley meal. As expected, the fat contents were higher for the oilseed diets than for CD.
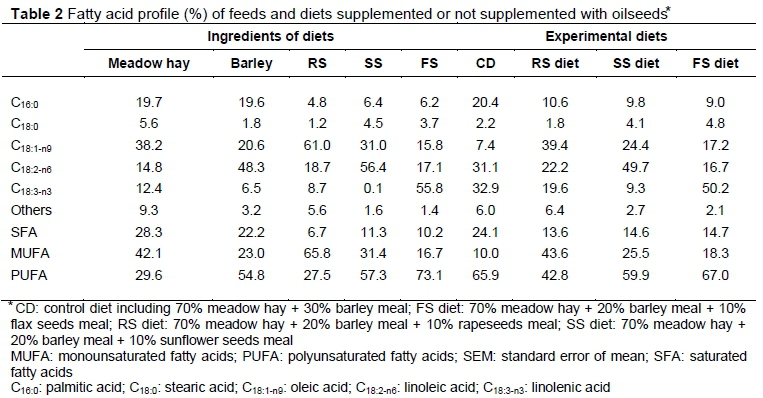
Table 2 shows the FA profile of the oilseeds and experimental diets. The FS diet contained the highest amount of α-linolenic acid (C18:3; 55.8%) among the oilseeds. The content of linoleic acid (C18:2) was highest in SS and reached 56.4% of total FA. The RS diet supplied the highest content of oleic acid (c18:1; 61% of measured FA). Overall, the concentrations of palmitic acid (C16:0) and stearic acid (C18:0) were low in all oilseeds and ranged from 4.8 to 6.4% for C16:0 and from 1.2 to 4.5% for C18:0.
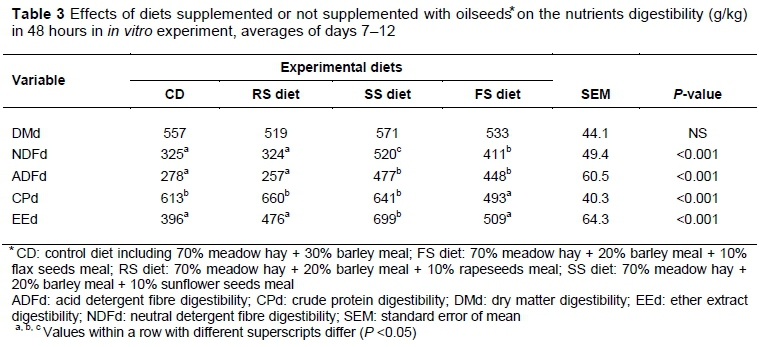
No differences (P >0.05) in dry matter digestibility (DMd) were observed between the diets in the present study (Table 3). Similarly, no differences in DMd were observed in diets with RS in experiments by Beauchemin et al. (2009) and Zhang et al. (2007). However, a decrease in DMd was observed in diets for lactating cows that contained FS or SS (Beauchemin et al., 2009) while an increase in DMd was reported in an experiment by Zhang et al. (2007), in which lactating ewes were fed a diet containing FS. The digestibility of neutral detergent fiber (NDFd; Table 3) was enhanced by the addition of SS and FS to the diet, compared with CD (P <0.001) diets. Inclusion of RS, relative to CD, had no effect on NDFd. The higher digestibility of acid detergent fibre (ADFd) was observed in the SS and FS diets compared with the CD (P <0.001) and RS diets (P <0.001). No differences were found between the CD and RS or between the SS and FS diets. In comparison with CD, feeding FS decreased the digestibility of crude protein (CPd). The low CPd of FS in the rumen of sheep was also reported by Zagorakis et al. (2015). According to Gonthier et al. (2004), FS supplementation increased post-ruminal digestibility. Additionally, FS had high fibre fraction digestibility (Gonthier et al., 2004; Zagorakis et al., 2015). The addition of oilseeds to the diet enhanced the digestibility of fat (EEd) compared with CD (P <0.001). Because of the higher amount of EE in oilseeds, the higher EEd in experimental diets was expected. In the present study, the authors did not confirm the negative effect of the addition of oilseeds to the high forage diets on the digestibility of nutrients. According to Moran (2005), dietary lipid content of about 5% should not cause harmful imbalance to the rumen environment and is not likely to affect digestibility negatively.
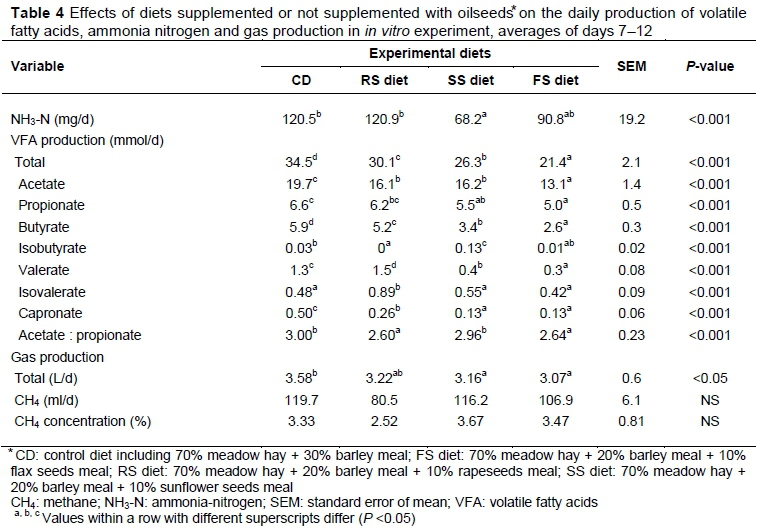
The addition of SS to the diet lowered the production of NH3-N (mg/d) in the effluent, compared with CD and RS diets (P <0.001; Table 4). The production of volatile fatty acids (VFA) (mmol/d) was lowered by the addition of oilseeds to the diet (p <0.001) (Table 4). The lowest production of VFA was observed in vessels with FS diet in comparison with other diets (P < 0.001). The FS added to the diet also decreased the production of acetate and butyrate, compared with other oilseed diets and CD (P <0.001). Additionally, the lowest acetate : propionate ratio was observed in vessels with RS and FS diets, compared with CD and SS diets (P <0.001). The FS affected the rumen fermentation pattern in a manner similar to that shown by Sutton et al. (1983) in a study with sheep, with a lower VFA concentration and smaller acetate : propionate ratio. This would indicate disturbed ruminal fibre degradation in the rumen (Jenkins, 1987), but the higher digestibility of fibre fractions was observed in our study. Simultaneously, the production of methane was not decreased (see below), thus the authors assume no reduction of microbial population and utilization of fiber fermentation products for creation of microbial biomass (Demeyer, 1991). Acetate : propionate ratio of 3.04.1 was reported in vivo and 2.0-4.1 in vitro in an experiment by Brown et al. (2002). High acetate : propionate ratio is an indication of proportionally higher digestible NDF in the feeds.
Generally, the addition of oilseeds to the high-forage diet reduced the amount of total fermentation gases (L/d) compared with CD, particularly for SS (P <0.05) and FS (P <0.01; Table 4). A positive correlation between gas and VFA production was detected by Getachew et al. (2004). Fermentative gas is produced mainly when feedstuffs are fermented to acetate and butyrate. However, although a decline in the production of fermentation gases was observed, the authors detected only a tendency for CH4 (%) decrease (P =0.06). Martin et al. (2008) fed various forms of flaxseeds (5.7% added fat) to dairy cows and reported a 10% reduction in CH4 (g/kg of dry matter intake (DMI)) for unprocessed seeds and a 49% reduction for the crude oil of the seeds. McGinn et al. (2004) added 5% sunflower oil to a forage-based diet for beef cattle and reduced CH4 (g/kg of DMI) by 17%. Beauchemin et al. (2007) added sunflower oil and whole sunflower seeds (3.4% added fat) to a forage-based diet fed to beef cattle and observed a 15% reduction in CH4 production. However, no changes in CH4 production were observed by Johnson et al. (2002), who added up to 5.6 % fat from a mixture of RS and whole cottonseeds to the diet for dairy cows. Similar results were obtained by Woodward et al. (2006), who fed dairy cows on pasture with a mixture of FS and fish oils. The literature indicates that supplemental fats could reduce CH4 emissions, but in many cases CH4-suppressing effects were influenced by the types of forage (maize silage versus hay) (Chung et al., 2011), and the amount of added fat (% in diet) and its treatment (Martin et al., 2008).
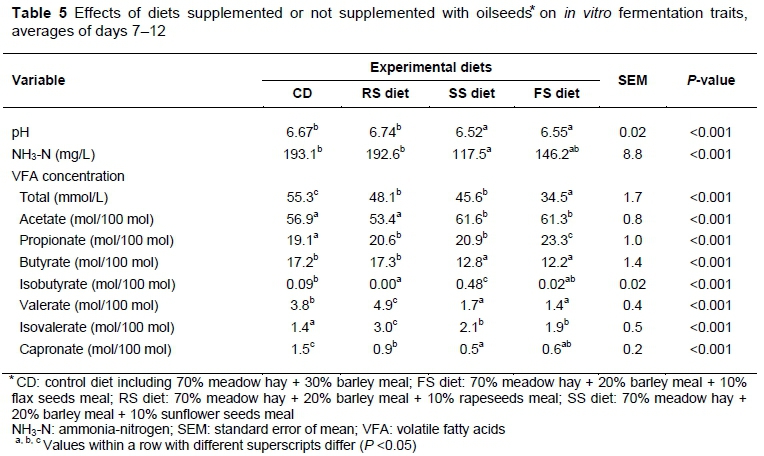
The inclusion of oilseeds reduced the pH values of the ruminal fluid in the vessels of SS and FS diets compared with those in the CD (P <0.01 and P <0.001, respectively) and RS diets (P <0.001; Table 5). No differences in pH were observed between the CD and RS diets or between the SS and FS diets. Despite these variations, the mean pH values remained within the physiological range (Krause & Oetzel, 2006). Martínez et al. (2010) compared pH values in rumen of sheep and Rusitec for a high-forage diet. The authors observed that ruminal pH before feeding and mean values of pH over the 12 hours post feeding were similar in both systems (6.69 and 6.45 in sheep and 6.57 and 6.32 in Rusitec). Following these results, the authors can conclude that oilseeds added to the high-forage diets should not have detrimental effects on pH values in the rumen of sheep. A decline in the NH3-N concentration (mg/L) in the vessel with SS, particularly in comparison with the CD and RS diets (P <0.001) (Table 5) was observed in the current study. According to Shingfield et al. (2008), adding SS oil to the diets of lactating cows tends to reduce the NH3-N concentration in the rumen. The NH3-N content in the liquid effluent from the vessels varied among diets from 45 to 207 mg/L in Boguhn et al.'s (2006) experiment, but the ammonia concentrations were greater than the 100 mg/L reported by Van Soest (1994) as the optimal concentration for efficient amino acid synthesis and microbial growth. Brito et al. (2006) suggested that a ruminal NH3-N concentration below 85 mg/L could depress microbial N synthesis in the rumen. In the present study, the mean NH3-N concentration ranged from 117 to 193 mg/L and indicates that NH3-N availability did not impair microbial growth. The addition of oilseeds diminished the amount of total VFA concentration (mmol/L) (Table 5). The lowest VFA concentration was detected in vessels with FS (P <0.001), while SS and RS also lowered the concentration of VFA, compared with CD (P <0.001 and P <0.01, respectively). Leupp et al. (2006) noted no changes in total VFA concentration on the addition of rapeseeds to the diets of steers fed with switchgrass hay. Compared with the CD and RS diets, the use of SS and FS enhanced the molar proportion of acetate and reduced the molar proportion of butyrate. All oilseeds, however, enhanced the molar proportion of propionate in effluent, compared with CD. Compared with other studies, the molar proportions of individual VFA, determined in Rusitec in the current study, were within the range determined by Martinez et al. (2010) for ruminants.
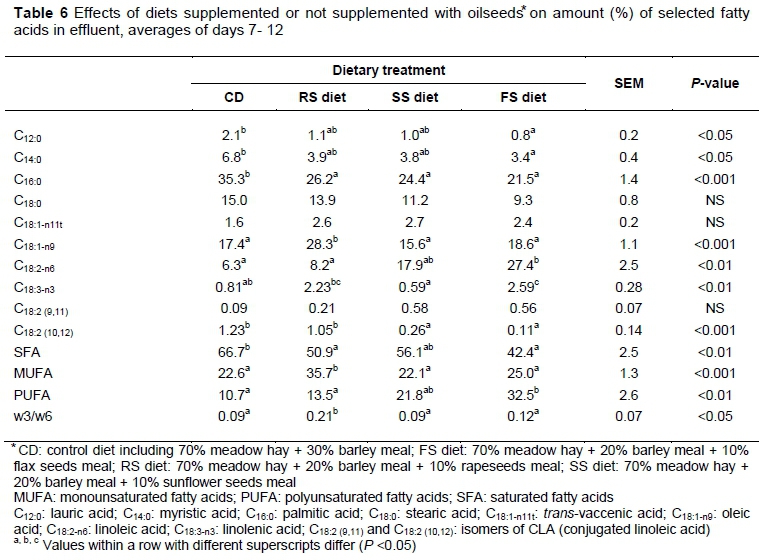
The addition of oilseeds to the diets affected the compositions of the fatty acids in the effluents (Table 6). The authors observed a decrease in the amount of saturated fatty acids (SFA) in the vessels with RS (P <0.05) and FS (P <0.001) compared with CD, but a higher amount of monounsaturated fatty acids (MUFA) was noted in the vessel with RS compared with the CD (P <0.001), SS (P <0.01), and FS diets (P <0.01). An increased amount of polyunsaturated fatty acids (PUFA) was detected only in the vessel with FS compared with CD (P <0.01) and RS (P <0.05). The higher ratio values (w3/w6) were observed in vessels with RS. The quantity of lauric acid (C12:0) was reduced only in FS compared with CD (P <0.05), and myristic acid (C14:0) was decreased by the addition of SS and FS compared with CD (P <0.05). The amount of stearic acid (C18:0), the end product of the biohydrogenation of unsaturated FA, was not influenced by the addition of oilseeds to the diets. The highest amount of oleic acid (C18:1) was found in the vessel with RS (P <0.001), which reflects the higher amount of this FA in this oilseed. The vessel with FS contained a higher amount of linoleic acid (C18:2|n-6) compared with the CD (P <0.01) and RS diets (P <0.05) and a higher amount of linolenic acid (C18:3n-3) compared with the CD (P <0.05) and SS diets (P <0.01). Loor et al. (2002) have shown that C18:1, C18:2, n-6and C18:3, n-3from dietary rapeseed and soybean oils are converted primarily to C18:0 and trans-C18:1 isomers in ruminal fluid as a result of PUFA biohydrogenation by rumen microbes. The amounts of trans-vaccenic acid (C18:1|n-11t) and c-9, t-11 CLA were not influenced by the addition of oilseeds; only the amount of t-10, c-12 CLA was decreased by the addition of SS and FS (P < 0.001) compared with the CD and RS diet. The amounts of C18:1, n-11tand c-9, t-11 CLA were not influenced in the ruminal fluids of fistulated cows fed FS in experiments by Cörtes et al. (2010) and the amount of c-9, t-11 CLA in the chymus was not influenced by the addition of sunflower oil to the diets of steers (Sackmann et al., 2003). Beaulieu et al. (2002) and Duckett et al. (2002) detected a higher amount of t-10, c-12 CLA after the addition of oil to diets with high portions of concentrates.
Conclusions
In the present study, up to 4.8% total fat did not affect the digestibility of nutrients negatively; the mean pH values remained within the physiological range. Unfortunately, only a tendency to CH4 decrease was detected. The values of FAs in effluent in vessels were influenced by the composition of diets. The addition of oilseeds, mainly RS and FS, decreased the amount of saturated FA and enhanced (only numerically, except vessels with FS) the amount of polyunsaturated FA in effluent. The SS and FS in diets enhanced the digestibility of fibre fractions; despite of this, the production of VFAs decreased. These data, which were obtained from an in vitro experiment, indicated that adding oilseeds to the high-forage diet did not have negative effects on the rumen microbial population.
Acknowledgments
This work was supported by the Ministry of Agriculture of the Czech Republic under grant MZERO0715 and the Scientific Grant Agency of the Ministry of Education of the Slovak Republic and the Slovak Academy of Sciences under grant VEGA 2/0009/14.
Authors' Contributions
PK and PH designed the study while its implementation, sample collection and data analysis were done by PK and DJ. PK and FJ participated in results, statistics and interpretation. PK wrote the draft manuscript, while DJ, FJ and PH edited it.
Conflict of Interest Declaration
The authors declare that they have no conflicts of interests with regard to this work.
References
AOAC (Association of Analytical Chemists), 2005. Official methods of analysis. 18th edition. AOAC, Gaithersburg, USA. [ Links ]
Beauchemin, K.A. & McGinn, S.M., 2006. Methane emissions from beef cattle: Effects of fumaric acid, essential oil, and canola oil. J. Anim. Sci. 84, 1489-1496. [ Links ]
Beauchemin, K.A., McGinn, S.M. & Petit, H.V., 2007. Methane abatement strategies for cattle: Lipid supplementation of diets. Can. J. Anim. Sci. 87, 431-440. [ Links ]
Beauchemin, K.A., McGinn, S.M., Benchaar, C. & Holtshausen, L., 2009. Crushed sunflower, flax, or canola seeds in lactating dairy cow diets: Effects on methane production, rumen fermentation, and milk production. J. Dairy Sci. 92, 2118-2127. [ Links ]
Beaulieu, A.D., Drackley, J.K. & Merchen, N.R., 2002. Concentrations of conjugated linoleic acid (cis-9 trans-11 octadecadienoic acid) are not increased in tissue lipids of cattle fed a high concentrate diet supplemented with soybean oil. J. Anim. Sci. 80, 847-861. [ Links ]
Boadi, D., Benchaar, C., Chiquette, J. & Massé, D., 2004. Mitigation strategies to reduce enteric methane emissions from dairy cows: Update review. Can. J. Anim. Sci. 84, 319-335. [ Links ]
Boguhn, J., Kluth, H. & Rodehutscord, M., 2006. Effect of total mixed ration composition and efficiency of ruminal microbial crude protein synthesis in vitro. J. Dairy Sci. 89, 1580-1591. [ Links ]
Brito, A.F., Broderick, G.A. & Reynal, S.M., 2006. Effects of varying dietary ratios of alfalfa silage to corn silage on omasal flow and microbial protein synthesis in dairy cows. J. Dairy Sci. 89, 3939-3953. [ Links ]
Brown, V.E., Rymer, C., Agnew, R.E. & Givens, D.I., 2002. Relationship between in vitro gas production profiles of forages and in vivo rumen fermentation patterns in beef steers fed those forages. Anim. Feed Sci. Technol. 98, 13-24. [ Links ]
Chung Y.H., He, M.L., McGinn, S.M., McAllister, T.A. & Beauchemin, K.A., 2011. Linseed suppresses enteric methane emissions from cattle fed barely silage, but not from those fed grass hay. Anim. Feed Sci. Technol. 166-167, 321-329. [ Links ]
Conway E.J., 1962. Microdiffusion analysis and volumetric error. 5th edition. Crosby Lockwood, London, UK. 322 pp. [ Links ]
Cörtes, C., da Silva-Kazama, D.C., Kazama, R., Gagnon, N., Benchaar, C., Santos, G.T.D. & Zeoula, L.M., 2010. Milk composition, milk fatty acid profile, digestion, and ruminal fermentation in dairy cows fed whole flaxseed and calcium salts of flaxseed oil. J. Dairy Sci. 93, 3146-3157. [ Links ]
Cottyn, B.G. & Boucque, C.V., 1968. Rapid method for the gas chromatographic determination of volatile fatty acids in rumen fluid. J. Agric. Food Chem. 16, 105-107. [ Links ]
Czerkawski, J.W. & Breckenridge, G., 1977. Design and development of a long-term rumen simulation technique (Rusitec). Br. J. Nutr. 38, 371-384. [ Links ]
Czerkawski, J.W. & Clapperton, J.L., 1984. Fats as energy-yielding compounds in the ruminant diet. In: J. Wiseman (ed). Fats in animal nutrition. Butterworths, Boston, MA. pp. 249. [ Links ]
Demeyer, D.I., 1991. Quantitative aspects of microbial metabolism in the rumen and hindgut. In: Jouany, J.P., Ed., Rumen microbial metabolism and ruminant digestion. INRA, Paris, pp. 217-237. [ Links ]
Dohme S., Machmüller, A., Wasserfallen, A. & Kreuzer, W., 2000. Comparative efficiency of various fats rich in medium chain fatty acids to suppress ruminal methanogens as measured with Rusitec. Can. J. Anim. Sci. 80, 473-482. [ Links ]
Duckett, S.K., Andrae, J.G. & Owens, F.N., 2002. Effect of high oil corn or added corn oil on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finishing diets. J. Anim. Sci. 80, 3353-3360. [ Links ]
Egan, A.R., Boda, K. & Varady, J., 1986. Regulation of nitrogen metabolism and recycling. In: L.P. Milligan, W.L. Grovum, W.L. & A. Dobson (eds). Control of digestion and metabolism in ruminants. Prentice Hall, Englewood Cliffs, NJ. pp. 386-402. [ Links ]
Folch, J., Lees, M. & Sloane-Stanley, G.H., 1957. A simple method for the isolation and purification of total lipids from animal tissues. J. Biolog. Chem. 226, 497-509. [ Links ]
Getachew, G., DePeters, E.J., Robinson, P.H. & Taylor, S.J., 2001. In vitro rumen fermentation and gas production: influence of yellow grease, tallow, corn oil and their potassium soaps. Anim. Feed Sci. Technol. 93, 1-15. [ Links ]
Getachew, G., Robinson, P.H., DePeters, E.J. & Taylor, S.J., 2004. Relationships between chemical composition, dry matter degradation and in vitro gas production of several ruminant feeds. Anim. Feed Sci. Technol. 111, 57-71. [ Links ]
Gonthier, C., Mustafa, A.F., Berthiaume, R., Petit, H.V., Martineau, R. & Ouellet, D.R., 2004. Effects of feeding micronized and extruded flaxseed on ruminal fermentation and nutrient utilization by dairy cows. J. Dairy Sci, 87, 1854-1863. [ Links ]
Johnson, K.A., Kincaid, R.L., Westberg, H.H., Gaskins, C.T., Lamb, B.K. & Cronrath, J.D., 2002. The effect of oilseeds in diets of lactating cows on milk production and methane emissions. J. Dairy Sci. 85, 1509-1515. [ Links ]
Krause, K.M. & Oetzel, G.R., 2006. Understanding and preventing subacute ruminal acidosis in dairy herds: A review. Anim. Feed Sci. Technol. 126, 215-236. [ Links ]
Leupp, J.L., Lardy, G.P., Soto-Navarro, S.A., Bauer, M.L. & Caton, J.S., 2006. Effects of canola seed supplementation on intake, digestion, duodenal protein supply, and microbial efficiency in steers fed forage-based diets. J. Anim. Sci. 84, 499-507. [ Links ]
Loor, J.J., Bandara, A.B.P.A. & Herbein, J.H., 2002. Characterisation of 18:1 and 18:2 isomers produced during microbial biohydrogenation of unsaturated fatty acids from canola and soybean oil in the rumen of lactating cows. J. Anim. Physiol. Anim. Nutr. 86, 422-434. [ Links ]
Machmüller, A., 2006. Medium-chain fatty acids and their potential to reduce methanogenesis in domestic ruminants. Agric. Ecosys. Environ. 112, 107-114. [ Links ]
Martin, C., Rouel, J., Jouany, J.P., Doreau, M. & Chilliard, Y., 2008. Methane output and diet digestibility in response to feeding dairy cows crude linseed, extruded linseed, or linseed oil. J. Anim. Sci. 86, 2642-2650. [ Links ]
Martinez, M.E., Ranilla, M.J., Tejido, M.L., Ramos, S. & Carro, M.D., 2010. Comparison of fermentation of diets of variable composition and microbial populations in the rumen of sheep and Rusitec fermenters. I. Digestibility, fermentation parameters, and microbial growth. J. Dairy Sci. 93, 3684-3698. [ Links ]
McDougall, E.I., 1948. Studies of rumen saliva. 1. The composition and output of sheep's saliva. Biochem. J. 43, 99-109. [ Links ]
McGinn, S.M., Beauchemin, K.A., Coates, T. & Colombatto, D., 2004. Methane emissions from beef cattle: Effects of monensin, sunflower oil, enzymes, yeast, and furmaric acid. J. Anim. Sci. 82, 3346-3356. [ Links ]
Moran, J., 2005. How the rumen works. In: Moran, J., Ed., Tropical dairy farming. Landlinks Press, Collingwood, VIC, Australia. p. 41-49. [ Links ]
Oldick, B.S. & Firkins, J.L., 2000. Effects of degree of fat saturation on fiber digestion and microbial protein synthesis when diets are fed twelve times daily. J. Anim. Sci. 78, 2412-2420. [ Links ]
Sackmann, J.R., Duckett, S.K., Gillis, M.H., Realini, C.E., Parks, A.H. & Eggelston, R.B., 2003. Effects of forage and sunflower oil levels on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finished diets. J. Anim. Sci. 81, 3174-3181. [ Links ]
SAS (Statistical Analysis Systems), 2000. User's guide (version 8.1). SAS Institute Inc., Raleigh, North Carolina, USA. [ Links ],
Shingfield, K.J., Ahvenjärvi, S., Toivonen, V., Vanhatalo, A., Huhtanen, P. & Griinari, J.M., 2008. Effect of incremental levels of sunflower-seed oil in the diet on ruminal lipid metabolism in lactating cows. Br. J. Nutr. 99, 971-983. [ Links ]
Sinclair, L.A., Cooper, S.L., Chikunya, S., Wilkinson, R.G., Hallett, K.G., Enser, M. & Wood, J.D., 2005. Biohydrogenation of n-3 polyunsaturated fatty acids in the rumen and their effects on microbial metabolism and plasma fatty acid concentrations in sheep. Anim. Sci. 8, 239-248. [ Links ]
Soder, K.J., Brito, A.F. & Rubano, M.D., 2013. Short communication: Effect of oilseed supplementation of an herbage diet on ruminal fermentation in continuous culture. J. Dairy, Sci. 96, 2551-2556. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Van Soest, P.J., 1994. Nutritional ecology of the ruminant. 2nd edition. Cornell University Press, Ithaca, NY, USA. [ Links ]
Woodward, S L., Waghorn, G.C. & Thomson, N.A., 2006. Supplementing dairy cows with oils to improve performance and reduce methane - Does it work? Proc. N. Z. Soc. Anim. Prod. 66, 176-181. [ Links ]
Zagorakis, K., Liamadis, D., Milis, Ch., Dotas, V. & Dotas, D., 2015. Nutrient digestibility and in situ degradability of alternatives to soybean meal protein sources for sheep. Small Rumin. Res. 67, 307-311. [ Links ]
Zhang, R.H., Mustafa, A.F. & Zhao, X., 2007. Effects of feeding oilseeds on nutrient utilization by lactating ewes. Small Rumin. Res. 67, 307-311. [ Links ]
Received 25 April 2017
Accepted 23 January 2018
First published online 4 April 2018
# Corresponding author: kubelkova.petra@vuzv.cz


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}