










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.3 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i3.12
ARTICLES
Orelha de Elefante Mexicans (Opuntia stricta [Haw.] Haw.) spineless cactus as an option in crossbred dairy cattle diet
R. C. SilvaI; M. A. FerreiraII; J. C. V. OliveiraIII; D. C. SantosIII; M. A. S. GamaIV; J. C. C. ChagasII, #; J. G. InácioII; E. T. S. SilvaV; L. G. R. PereiraIV
IFederal Institution of Education, Science and Technology of Alagoas - 57460000 - Piranhas, AL - Brazil
IIFederal Rural University of Pernambuco - Department of Animal Science - 52171900 - Recife, PE - Brazil
IIIAgronomic Institute of Pernambuco - 56519360 - Arcoverde, PE - Brazil
IVEmpresa Brasileira de Pesquisa Agropecuária (EMBRAPA) - 36038-330 - Juiz de Fora, MG - Brazil
VFederal Rural University of Pernambuco - Garanhuns Academic Campus - 55292270 - Garanhuns, PE - Brazil
ABSTRACT
A new genotype of spineless cactus is being used in the diets of dairy cattle that are raised in semi-arid regions. However, little is known about its nutritional value. This study aimed to evaluate the effects of replacing Miúda (Nopalea cochenillifera Salm Dyck) with Orelha de Elefante Mexicana (Opuntia stricta [Haw.] Haw.) spineless cactus, on nutrient intake and digestibility, milk yield and composition, feeding behaviour, microbial protein synthesis, nitrogen balance, and ruminal and blood parameters of dairy cows. Ten Girolando cows, 500 ± 51.6 kg bodyweight, were distributed in a double Latin square design 5 x 5. The treatments consisted of replacement levels of Miúda (MIU) by Orelha de Elefante Mexicana (OEM) at 0, 25, 50, 75, and 100%. The intake and digestibility of dry matter (DM) (14.38-12.95 kg d-1, 716.3-658.9 g d-1), organic matter (OM) (13.01-11.43 kg d-1, 747.8-704.8 g d-1), crude protein (CP) (2.02-1.61 kg d-1, 863.8845.2 g d-1) and total digestible nutrients (TDN) (9.38-7.92 kg d-1) decreased linearly with the increase in replacement. Despite the decrease in intake and digestibility, the supply of nutrients was sufficient to maintain a milk yield of 12.5 kg d-1. The average daily weight gain decreased linearly with the increase in replacement, while protein microbial efficiency (g microbial CP kg-1 TDN intake; 91.24 to 127.44 g kg-1) increased linearly. Thus, OEM could replace 100% MIU in diets with 48% of spineless cactus, for crossbred lactating cows with 12.5 kg d-1 milk yield. Therefore, OEM is a viable new option for producing milk in smallholder livestock systems in semi-arid regions.
Keywords: Alternative forage, milk yield, ruminal parameter, semi-arid, smallholder livestock system
Introduction
In Brazil, the smallholder system produces 70% of food consumed in the country, with milk production represented by 58% (MDA, 2015). This livestock system characterizes the semi-arid regions of the Brazilian northeast, and one of its few viable economic activities is dairy production (Oliveira et al., 2016).
The spineless cactus is an essential forage for livestock in Brazilian semi-arid regions. It has adaptive agronomic characteristics, such as being tolerant to hydric stress, and being suitable as forage in diets for dairy goats, dairy cattle, and sheep (Mahouachi et al., 2012; Monteiro et al., 2014; Lins et al., 2016, respectively). It also has a high content of non-fibrous carbohydrates, and consequently significantly high energy content compared with other forage plants (Ben Salem, 2010; Siqueira et al., 2017). However, thousands of hectares have been decimated by infestations of cochineal carmine (Dactylopius sp.). This plague is expanding steadily through spineless cactus areas as a result of favourable conditions during the prolonged drought of 2012 in the Brazilian northeast.
During this time, the main spineless cactus genotype that was used was the Gigante (Opuntia ficus-indica (L.) Mill.), a plant that is highly susceptible to cochineal. Even for varieties that are resistant to such pests, most dairy farmers did not anticipate the effect of cochineal carmine. The exclusive use of Gigante (Opuntia ficus-indica (L.) Mill.) spineless cactus could be considered a monoculture, which brought disastrous social and environmental consequences.
Another genotype in spineless cactus that is widely used is Miúda (Nopalea cochenillifera), which is resistant to cochineal carmine and has well-known nutritional value (Batista et al., 2013; Ferreira et al., 2014). The best strategy to control such pests is to cultivate spineless cactus varieties that are resistant to cochineal carmine. This is also an environment-friendly practice. Thus, a resistant genotype is of great importance for smallholder farming because it dispenses with costly chemicals that pollute the environment, and contributes to production system sustainability.
Currently, the search for spineless cactus genotypes that are resistant to cochineal carmine has highlighted Orelha de Elefante Mexicana (Opuntia stricta [Haw.] Haw.) in particular because of its greater DM productivity than Miúda (Lopes et al., 2010). However, while the Orelha genotype is widespread among milk producers in all semiarid regions of Brazil, knowledge about its nutritional value is limited. Ferreira et al. (2012) observed DM intake decrease for crossbred cow fed Orelha compared with Miúda, and a consequential decrease in milk production. The authors associated this response with low Orelha palatability; hence, further investigation is required.
It was hypothesized that a ratio of Miúda and Orelha de Elefante Mexicana might maximize intake and digestion of nutrients and consequently dairy performance. This study aimed to evaluate the effects of replacing Miúda with Orelha de Elefante Mexicana spineless cactus on nutrient intake, digestibility, milk yield and composition, feeding behaviour, microbial protein synthesis, nitrogen balance, and ruminal and blood parameters of Girolando lactating cows.
Materials and Methods
All procedures were conducted in accordance with the guidelines set out by the Brazilian College of Animal Experimentation in the Code of Practice for the Care and Use of Animal for Experimental Purposes and were reviewed and approved by the Ethics Committee on Use of Animal for Research (CEUA) of the Federal Rural University of Pernambuco, under license number 010/2015.
The experiment was conducted at the experimental station in Arcoverde city, Pernambuco State (8° 25' 15" S, 37° 3' 41" W; and altitude 663 m), which has a semiarid climate (Bsh) according to the Köppen climate classification.
Ten multiparous Girolando cows (5/8 Holstein-Gyr), averaging (mean ± SD) 13.36 ± 2.27 kg milk d-1, 60 ± 1.66 days in milk, and 500 ± 51.6 kg of bodyweight were distributed in a double contemporary 5 x5 Latin square design.
The pre-adaptation of animals to management and facilities lasted 21 days. The first 14 days were for adaptation of the animals to the diets, followed by seven days for collecting data and samples. The experimental period lasted 105 days, divided into five periods of 21 days. The cows were housed in 24 m2 semi-covered individual pens equipped with drinking and feeding troughs.
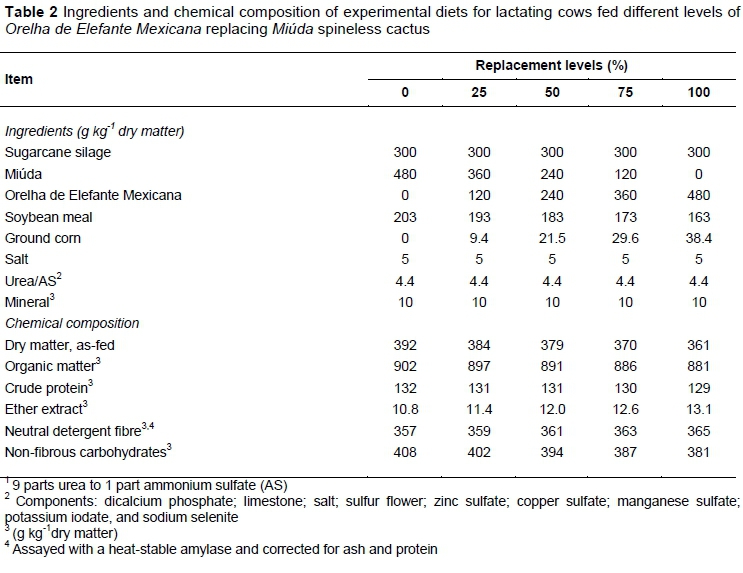
The experimental diets were formulated according to NRC (2001) regulations for cows producing 12 kg d-1 of milk with 4% of fat (Tables 1 and 2). The diets were isonitrogenous and formulated to meet protein requirements. The roughage/concentrate ratio was 78 : 22. The feed was provided ad libitum as total mixture divided into two meals at 7h00 and 16h00, allowing 12% leftovers. Feed leftovers were weighed daily prior to the morning meal to estimate the feed intake from the previous day and adjust the supply.
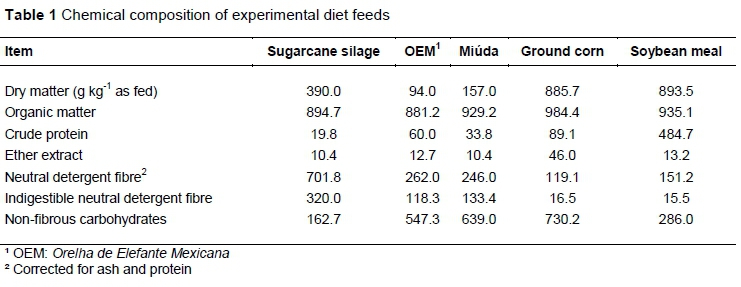
The chemical composition of the ingredients, ratios, and chemical composition of the experimental diets with various replacement proportions of Miúda spineless cactus (Nopalea cochenillifera Salm Dyck) for Orelha de Elefante Mexicana spineless cactus (Opuntia stricta [Haw.] Haw.), namely 0, 25, 50, 75, 100 % are shown in Tables 1 and 2.
The feed was mixed at the beginning of the experiment. One sample of each feed was collected during the mixing process. Throughout the collection period, weighing and samplings of sugarcane silage, spineless cactus, concentrated feed, and feed leftovers of each animal were made. At the end of each 21-day period, composite samples were prepared per animal. They were stored at -18 °C for subsequent chemical analyses.
At the end of the experiment, samples were thawed, pre-dried (55 °C for 72 h) in a forced-air circulation oven and ground in a Wiley knife mill, part with 1 mm porosity sieves and the rest with 2 mm sieves.
The bodyweight (BW) of each cow was recorded at the beginning and end of each experimental period after milking.
The feeding behaviour of all cows was evaluated from the tenth day of each experimental period (every 5 min for 24 h) according to the sampling method proposed by Martin & Baterson (2007). Feed and rumination efficiencies (kg DM h-1) were calculated by dividing the intake by total feeding time (feed efficiency) or rumination time (rumination efficiency).
To estimate the apparent digestibility coefficients, indigestible neutral detergent fibre (iNDF) obtained after 288 hours of ruminal incubation was used as an internal marker (Valente et al., 2015). Samples of feed, leftovers, and faeces were analysed to determine the concentrations of DM (method 934.01), mineral matter (MM) (method 942.05), crude protein (CP) (method 990.13), and ether extract (EE) (method 920.39) according to the AOAC (2005). The evaluations of concentrations of neutral detergent fibre (NDF) and ash corrections followed the protocols suggested by Mertens (2002). Neutral detergent insoluble nitrogen (NDIN) was analysed according to Licitra et al. (1996). Non-fibrous carbohydrates were calculated according to Hall (2000) and the intake of total digestible nutrients (CTDN) was calculated according to Sniffen et al. (1992).
Faecal samples were collected directly from the rectum of animals from the 15th to the 19th day of each experimental period at different times after feeding (6h00, 8h00, 10h00, 12h00, and 14h00) according to Ítavo et al. (2002). Then, the samples were frozen and stored at -18 °C for later pre-drying in a forced-air circulation oven (55 °C for 72 h) and grinding in Wiley knife mills, using a 1-mm sieve for part of the samples and a 2-mm sieve for the rest.
Milk samples were collected on the 18th and 19th days from the collectors of the milking machine in the morning and in the afternoon at 6h00 and 15h00 during each experimental period for the analysis of proteins, fat, lactose, total solids, and urea according to the methods proposed by ISO 9622/IDF141C (2013). An aliquot of milk was collected for allantoin analysis. Milk yield values were adjusted to 4% fat according to the NRC (2001).
Blood samples were collected on the 21st day four hours after morning feeding by puncturing the coccygeal vein. Samples were stored in two tubes, one containing anticoagulant (EDTA) and the other containing sodium fluoride. After collection, the blood samples were immediately centrifuged at 2,700 rpm for 20 min. The plasma samples were frozen at -18 °C for subsequent analysis of urea nitrogen, glucose, beta-hydroxybutyrate, and non-esterified fatty acids concentrations. Samples were analysed using commercial colorimetric system kits (Diagnostic Labtest® SA).
Samples of urine spot were obtained on the 21st day of each experimental period immediately after blood collection and four hours after the first daily supply of feed (Chizzotti et al., 2008). The urine was filtered. One 10 mL aliquot was immediately diluted in 40 mL of 0.036 N sulfuric acid and stored at -20 °C for later analysis of urea, allantoin (Chen & Gomes, 1992), and uric acid (George et al., 2006). Creatinine, urea and uric acid were analysed with commercial colorimetric system kits (Labtest Diagnóstica® SA).
The daily urinary excretion was estimated from a proposed excretion of creatinine of 24.05 mg kg-1 of bodyweight (Chizzotti et al., 2008). The microbial production was calculated according to Chen and Gomes (1992) considering an absorbed purine recovery of 0.85 and a contribution to the endogenous purine excretion as recommended by Gonzalez-Ronquillo et al. (2003).
Ruminal content samples were collected on the 21st day using a gavage according to the methodology proposed by Duffield et al. (2004). The collection was performed four hours after the first morning meal, using a flexible hose and a vacuum pump. Between collections, the hose was washed and lubricated with petroleum jelly for its first 30 cm. A sample of 200 mL rumen fluid was collected per animal. The rumen fluid was filtered with a gauze and the pH was measured with a portable potentiometer (Kasvi). Immediately after collection, the liquid was frozen for later analyses. The analyses of volatile fatty acids were performed using gas chromatography (FINNIGAN 9001, USA). Approximately 50 mL of ruminal fluid were acidified with 1 mL sulfuric acid 1 : 1 for subsequent ammoniacal nitrogen (N-NH3) analysis. The samples were stored at -15 °C. N-NH3 was analysed by Kjeldahl distillation according to the AOAC (2005).
The balance of nitrogenous compounds was obtained from the difference between the total nitrogen intake (Nint) and total nitrogen excreted in the faeces (N-faeces), urine (N-urine) and milk (N-milk). The calculation of nitrogen use efficiency was performed by dividing the average total N-milk (g d-1) by the average dietary intake of nitrogen (g d-1) according to methods proposed by Broderick (2003).
The data were submitted for analysis of variance and regression using the MIXED procedure of the statistical program SAS (version 9.4, SAS Institute Inc., Cary, NC, USA), adopting 5% as significance level for the type I error, according to the following model:

where: yijkl = observation ijkl
μ = general mean
Ti = treatment fixed effect i
Qj = square fixed effect j
Pk = period fixed effect k
(A/Q)y = animal l into square j random effect
T*Qij = interaction effect of treatment i and square j
εijkl= random error with mean 0 and variance σ
Results and Discussion
The MIU showed higher DM (+67 %), OM (+5.5 %), and NFC (+16 %) contents than OEM (Table 1). However, the diets had similar chemical compositions (Table 2). The DM, OM, CP, and TDN intake decreased linearly with the replacement of MIU for OEM (Table 3; P <0.05). The digestibility of DM, OM, and CP decreased linearly with the inclusion of OEM (Table 3; P >0.05). Nonetheless, the feeding behaviour was not altered by the function of replacement (Table 4; P >0.05).
The decrease in the intake of CP, NFC, and TDN observed at larger inclusions of OEM are associated with lower DM intake, as MIU was replaced. The OEM showed higher MM content (5%) than MIU, and consequently lower OM content (Table 1). Therefore, the decrease in OM intake at higher levels of OEM in the diets is associated with the lowest OM intake of such cultivar and a higher MM content.
The DM content of MIU (Table 1) was higher than in OEM (+67 %), and is an intrinsic characteristic of OEM that may promote higher volumes of natural matter (NM) ingested for greater levels of OEM diets, promoting a superior volume in gut compared with48 % levels MIU diet. In this study, the intake of NM was 58.48, 64.79, 70.66, 77.45, and 79.23 kg d-1 at 0, 25, 50, 75, and 100% replacement of MIU for OEM, respectively, and the dietary DM decreased from 24.59 to 16.34 with the replacement of MIU for OEM. This may have led to a lower intake of DM (Table 3) when using OEM due to ruminal distension, and providing a greater DM intake with MIU. The low DM content results in the intake of larger quantities of NM by the animal. According to Gebremariam et al. (2006), high amounts of spineless cactus may limit the DM intake because it contains high moisture content (Table 2). During this study, it was observed that animals that received experimental diets exclusively with MIU, or with high levels of MIU, showed a greater voracity at feed supply. The higher carbohydrate content contained in MIU could provide high palatability (Ferreira et al., 2012), which might possibly explain the high intake of DM and TDN in diets with higher levels of MIU.
According to the NRC (2001), the diets adequately met the nutritional requirements of cows (500 kg BW) producing 12 kg milk with 4% fat, and consuming an estimated 14.14, 1.74 and 7.6 kg d-1 DM, CP and TDN, respectively. For 44% spineless cactus present in diet, DM intake was highest (+2.55 kg) with 14.96 kg d-1 MIU and 12.11 kg d-1 OEM (Table 3). The CP intake was 2.18 vs. 1.90, the TC was 10.65 vs. 8.64, the NFC was 5.08 vs. 3.72, and the TDN was 9.51 vs. 7.25 kg d1, for MIU and OEM, respectively. The excess nutrients provided by greater inclusion of MIU diet may be diverted to other functions, such as weight gain (Sundrum, 2015). Data presented by Ferreira et al. (2012) corroborates with intakes presented in this study.
The higher DM intake and OM content of MIU compared with OEM provided a greater amount of fermentable OM in the rumen, particularly due to high NFC content (MIU: 64 % and OEM: 55 %). The digestibility of DM, OM and CP improved as the proportion of MIU in the diet increased. Thus, the linear decrease in dietary DM intake within creasing OEM levels resulted in a decrease in apparent digestibility of DM, OM and CP. The high content of NFC in spineless cactus is an important nutritional quality that acts as a determinant of the rapid degradation observed in spineless cactus, resulting in high energy release in the rumen (Ben Salem et al., 2002; Batista et al., 2003; Barros et al., 2017).
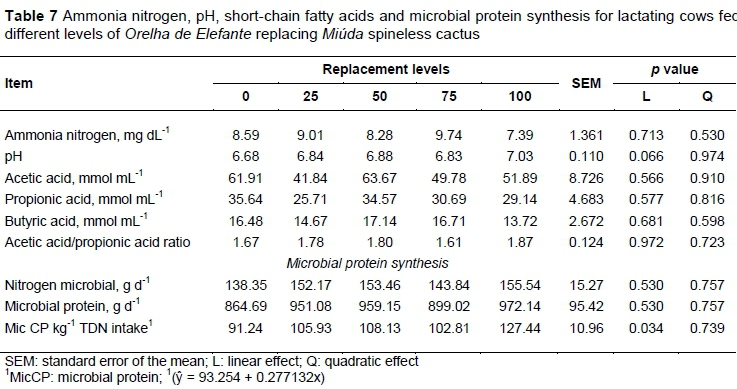
The results indicate that increasing levels of OEM in the diets was not a limiting factor for microbial protein synthesis, resulting in an output of 865-972 g microbial CP d-1 with the replacements. However, the levels provided greater microbial efficiency (g Mpse kg-1 TDN intake), from 91.24 to 127.44 g, in response to the lower intake of NDT with higher proportions of OEM. The availability of N and energy in the rumen is the main factor limiting microbial growth (Uddin et al., 2015). According to Broderick (2003), there is a complex interrelation between dietary protein and the amount of energy used by ruminants. With the inclusion of OEM, small amounts of corn (0, 0.94, 2.15, 2.96, and 3.84% in the DM of the diet) were added to balance the CP in the diets, contributing, consequently, to the improvement of the protein/energy ratio for diets. According to the results, there were no energy and protein limitations in the rumen.
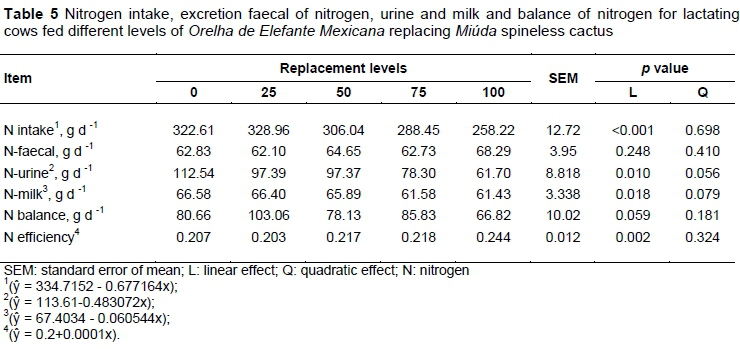
The N content in urine and milk decreased and N efficiency use increased linearly with the replacement of MIU for OEM (Table 5; P <0.05). The CP intake, and consequently N intake, is linked to the DM intake, which was lower according to greater levels of OEM in diets, making it clear that there was no diet sorting. Thus, the linear decrease in the intake of DM, CP, and N, resulting in a proportional decrease in N excretion through urine and milk.
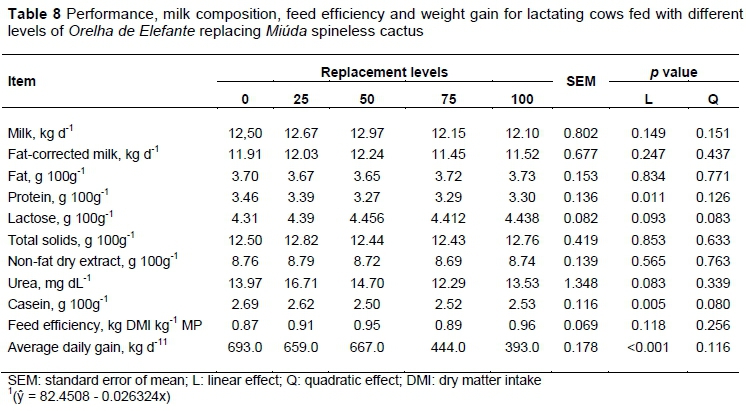
Related to the N efficiency pattern, a linear increase was expected after the cows had achieved production potential (12.5 kg/day of milk) and the excess of N consumed was diverted to other functions, as body weight gain (Table 8). However, to determine the N efficiency values were considered the N intake values related to N excretion in milk, and as a result was observed a decrease in N efficiency (Table 5).
Although a negative linear effect was observed on N intake, the positive N balance obtained indicates a retention of proteins in vivo. According to this, potential production was achieved and a portion of nutrients were metabolized for body deposition, as evidenced by the weight gain (Table 8)
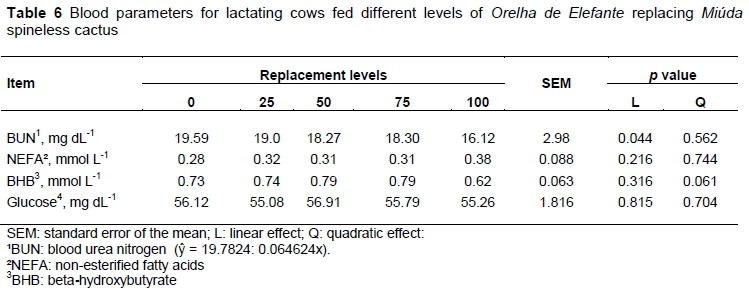
Non-esterified fatty acids (NEFA), glucose and beta-hydroxybutyrate (BHB) in the plasma did not vary among the diets (Table 6; P >0.05); otherwise, the blood urea nitrogen (BUN) decreased linearly according to replacement of MIU by OEM (Table 6; P <0.05). The behaviour observed for the concentration of BUN is in accordance with the intake of CP and N, which also showed a negative linear trend in the diets with increasing proportion of OEM. According to Oliveira et al. (2001), BUN concentrations above 19 mg dL-1 represent the limit for dietary N loss. In this study, the diet with MIU only had 19.59 mg dL-1 BUN and its concentration decreased linearly with increasing OEM content. In general, it did not exceed the recommended value, indicating there was no excess protein in the diet.
According to the NRC (2001), the reference value for microbial synthesis efficiency is 130 g microbial CP kg-1 TDN intake. However, under tropical conditions, according to Pina et al. (2010), the reference value is 120 g microbial CP kg-1 TDN intake. In this study, the reference value recommended by the authors corresponded to the experimental diet that presented the highest replacement level of MIU for OEM (Table 7).
Ammonia nitrogen (N-NH3) concentration in the rumen fluid, rumen pH, concentrations of acetic, propionic and butyric acids, and acetic acid : propionic acid ratio were not altered among the diets, nor microbial nitrogen (P >0.05). The replacement of MIU for OEM increased microbial protein synthesis efficiency linearly (Table 7; P <0.05). The average pH values of the ruminal liquid obtained in the replacement of MIU for OEM were close to those considered normal. According to Van Soest (1994), normal cellulolytic bacteria activity occurs at pH 6.2-7.2; and an optimal at pH 6.7 (Hackmann & Firkins, 2015), which is comparable with the average pH in this study (pH 6.8) (Table 7).
Milk yield, milk yield corrected to 4% of fat, fat, total solids, protein, lactose, casein, non-fat dry extract, urea, and feed efficiency (P >0.05) were not altered by replacement of MIU by OEM. The average daily weight gain decreased linearly with the replacements (Table 8) (P <0.05).
Considering the nutrients requirements balancing and the production potential of the animals no difference in milk yield was observed. However, the experimental diet with MIU promoted nutrients beyond the requirement necessary, according to a high DM and TDN intake. This fact could be important in situations where lactating cows are in negative energy balance; thus, contributing to the recovery of body condition through weight gain, as evidenced by the highest average daily gain for animals receiving higher proportions of MIU (Table 8).
Conclusion
It is possible to replace 100% Miúda with Orelha de Elefante Mexicana genotype in lactating crossbred cows diets based on 48% spineless cactus promoting 12.5 kg d-1 milk yield. The genotype tested is a new forage option for producing milk in smallholder livestock systems in semiarid regions, providing agricultural diversification.
Acknowledgements
The authors are grateful to the Brazilian Foundation, Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) by financial support, and the Agronomic Institute of Pernambuco (IPA).
Authors' Contributions
Conception, design of experiments and supervision of project by MAF, JCVO and DCS. RCS conducted the experiment and data analysis. All authors participated in interpretation of results and writing and integration of the manuscript. Critical revision and final approval of version to be published was also done by all authors.
Conflict of Interest Declaration
We wish to confirm that there are no known conflicts of interest associated with this publication and there has been no significant financial support for this work.
References
AOAC., 2005. Official methods of analysis. 18th ed. AOAC International, Gaithersburg, Maryland, USA. [ Links ]
Barros, L.J.A., Ferreira, M.A., Oliveira, J.C.V., Santos, D.C., Chagas, J.C.C., Alves, A.M.S.V., Silva, A.E.M. & Freitas, W.R., 2017. Replacement of Tifton hay by spineless cactus in Girolando post-weaned heifers' diets. Trop. Anim. Health Prod. 1-6. [ Links ]
Batista, A.M.V., Carvalho, F.F.R. & Rocha Filho, R.R., 2013. A palma forrageira na alimentação de ruminantes no semiárido brasileiro. In: Simpósio Brasileiro de Produção de Ruminantes, 2, 2013, Itapetinga. Anais... Itapetinga: Simpósio Brasileiro de Produção de Ruminantes. [ Links ]
Batista, A.M.V., Mustafa, A.F., Santos, G.R.A., de Carvalho, F.F.R., Dubeux Jr, J.C.B., Lira, M.A. & Barbosa, S.B.P., 2003. Chemical composition and ruminal dry matter and crude protein degradability of spineless cactus. J. Agron. Crop. Sci. 189, 123-126. [ Links ]
Ben Salem, H., 2010. Nutritional management to improve sheep and goat performances in semiarid regions. R. Bras. Zootec. 39, 337-347. [ Links ]
Ben Salem, H., Nefzaoui, A. & Ben Salem, L., 2002. Nitrogen supplementation improves the nutritive value of Opuntia ficus-indica F. inermis-based diets and sheep growth. Acta Hortic. 581, 317-321. [ Links ]
Broderick, G.A., 2003. Effects of varying dietary protein and energy levels on the production of lactating dairy cows. J. Dairy Sci. 86: 1370-138. [ Links ]
Chen, X.B. & Gomes, M.J., 1992. Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives - an overview of technical details. International Feed Research Unit. Rowett Research Institute. Aberdeen, UK. [ Links ]
Chizzotti, M.L., Valadares Filho, S.C. & Valadares, R.F.D. 2008. Determination of creatinine excretion and evaluation of spot urine sampling in Holstein cattle. Livest. Sci. 113, 218-225. [ Links ]
Duffield, T., Plaizier, J.C., Fairfield, A., Bagg, R., Vessie, G., Dick, P., Wilson, J., Aramini, J. & McBride, B., 2004. Comparison of techniques for measurement of rumen pH in lactating dairy cows. J. Dairy Sci. 87, 59-66. [ Links ]
Ferreira, M.A. & Urbano S.A., 2014. Novas Tecnologias para Alimentação de Bovinos Leiteiros na Seca. Rev. Científica. Prod. Ani. 15, 42-52. [ Links ]
Ferreira, M.A., Bispo, S.V., Rocha Filho, R.R., Urbano, S.A. & Costa, C.T.F., 2012. The use of cactus as forage for dairy cows in semi-arid regions of Brazil. P.1-22. In: Konvalina, P., (ed). Organic Farming and Food Production. InTech, South Bohemia, EU. [ Links ]
Gebremariam, T., Melaku, S. & Yami, A., 2006. Effect of different levels of cactus (Opuntia ficus-indica) inclusion on feed intake, digestibility and body weight gain in teff (Eragros tistef) straw-based feeding of sheep. Anim. Feed Sci. Technol. 131, 43-52. [ Links ]
George, S.K., Dipu, M.T., Mehra, U.R., Singh, P., Verma, A.K. & Ramgaokar, J.S., 2006. Improved HPLC method for the simultaneous determination of allantoin, uric acid and creatinine in cattle urine. J. Chromatogr. 832, 134-137. [ Links ]
González-Ronquillo, M., Balcells, J. & Guada, J.A., 2003. Purine derivative excretion in dairy cows: Endogenous excretion and the effect of exogenous nucleic acid supply. J. Dairy Sci. 86, 1282-1291. [ Links ]
Hackmann T.J. & Firkins J.L., 2015. Maximizing efficiency of rumen microbial protein production. Front. Microbiol. 6, 465. [ Links ]
Hall, M.B., 2000. Neutral detergent-soluble carbohydrates. Nutritional relevance and analysis. Gainesville: University of Florida, USA. [ Links ]
ISO 9622/IDF141C. 2013. Determination of milk fat, protein and lactose content: Guidance on the operation of mid- infrared instruments. Bruxelas, BEL. [ Links ]
Ítavo, L.C.V., Valadares Filho, S.C., Ferreira da Silva, F., Valadares, R.F.D., Paulino, M.F., Ítavo, C.C.B.F. & Moraes, E.H.B.K., 2002. Comparação de indicadores e metodologia de coleta para estimativas de produção fecal e fluxo de digesta em bovinos. R. Bras. Zootec. 31, 1833-1839. [ Links ]
Johnson, T.R. & Combs, D.K., 1991. Effects of prepartum diet, inert rumen bulk, and dietary polythyleneglicol on dry matter intake of lactating dairy cows. J. Dairy Sci. 74, 933-944. [ Links ]
Licitra, G., Hernandez, T.M. & Van Soest, P.J., 1996. Standardization of procedures for nitrogen fractionation of ruminant feeds. Anim. Feed Sci. Technol. 57, 347-358. [ Links ]
Lins, S.E.B., Pessoa, R.A.S., Ferreira, M.A., Campos, J.M.S., Silva, J.A.B.A., Silva, J.L., Santos, S.A. & Melo, T.T.B., 2016. Spineless cactus as a replacement for wheat bran in sugar cane-based diets for sheep: Intake, digestibility, and ruminal parameters. R. Bras. Zootec. 45, 26-31. [ Links ]
Lopes, E.B., Brito, C.H., Albuquerque, I.C. & Batista, J.L. 2010. Selection of cactus pear forage (Opuntia spp.) and (Nopalea spp.) genotypes resistant to the carmine cochineal (Dactylopius opuntia Cockerell, 1929) in the state of Paraíba, Brazil. Engenharia Ambiental - Espírito Santo do Pinhal- Pesquisa e Tecnologia 7, 204-215. [ Links ]
Mahouachi, M., Atti, N. &. Hajji, H., 2012. Use of spineless cactus (Opuntia ficus indica F. Inermis) for dairy goats and growing kids: Impacts on milk production, kid's growth, and meat quality. Sci. World J. 2012, 321567ID. [ Links ]
Martin, P. & Bateson, P., 2007. Measuring behaviour: An introductory guide. 3rd ed. Cambridge, Cambridge, UK. [ Links ]
McDermott, J.J., Staal, S.J., Freeman, H.A., Herrero, M. & Van de Steeg, J.A. 2010. Sustaining intensification of smallholder livestock systems in the tropics. Livest. Sci. 130, 95-109. [ Links ]
MDA., 2014. ONU Reinforces the importance of family farming to the world. Ministério da Agricultura Familiar e do Desenvolvimento Agrário (MDA), Brasilia, Brazil. Available at http://www4.planalto.gov.br/consea/comunicacao/noticias/2014/onu-reforca-a-importancia-da-agricultura-familiar-para-o-mundo. Accessed 15 January 2017. [ Links ]
Mertens, D.R., 2002. Gravimetric determination of amylase-treated neutral detergent fiber in feeds with refluxing in beakers or crucibles: collaborative study. J. AOAC Int. 85, 1217-1240. [ Links ]
Monteiro, C.C.F., Melo, A.A.S., Ferreira, M.A., Campos, J.M.S., Souza, J.S.R., Silva, E.T.S., Andrade, R.P.X. & Silva, E.C., 2014. Replacement of wheat bran with spineless cactus (Opuntia ficus indica Mill cv. Gigante) and urea in the diets of Holstein x Gyr heifers. Trop. Anim. Health Prod. 46, 1149-1154. [ Links ]
Nardone, A., Ronchi, B., Lacetera, N., Ranieri, MS. & Bernabucci, U., 2010. Effects of climate changes on animal production and sustainability of livestock systems. Livest. Sci. 130, 57-69 [ Links ]
NRC (National Research Council), 2001. Nutrient requirements of dairy cattle. 7th ed. NRC Academic Science, Washington, DC, USA. [ Links ]
Oliveira, M.C., Campos, J.M.S., Oliveira, A.S., Ferreira, M.A. & Melo, A.A.S., 2016. Benchmarks for milk production systems in the Pernambuco Agreste region, north-eastern Brazil. Rev. Caatinga. 29, 725-734. [ Links ]
Oliveira, M.D.S., Casagrande, A.A. & Oliveira, E.F.S., 2001. Effect of in vitro digestibility of sugarcane varieties on their value as feed for cattle. ARS Veterinária 17, 238-243. [ Links ]
Pina, D.S., Valadares, R.F.D., Valadares Filho, S.C. & Chizzotti, M.L., 2010. Rumen degradation of food protein and microbial protein synthesis. p. 13-46. In: Valadares Filho, S.C., Marcondes, M.I., Chizzotti, M.L. & Paulino, P.V.R, eds. Nutrient requirements of zebu beef cattle - BR-Corte. Suprema, Visconde do Rio Branco, MG, BR. [ Links ]
Siqueira, M.C.B., Ferreira, M.A., Monnerat, J. P.I. dos S., Silva, J. de L., Costa, C.T.F., Conceição, M.G. da., de Andrade, R. de P.X., Barros, L.J.A. & Melo, T.T.B., 2017. Optimizing the use of spineless cactus in the diets of cattle: Total and partial digestibility, fiber dynamics and ruminal parameters. Anim. Feed Sci. Technol. 226, 56-64. [ Links ]
Sniffen, C.J., O'Connor, J.D., Van Soest, P.J., Fox, D.G. & Russel, J.B., 1992. A net carbohydrate and protein system for evaluating cattle diets; II. Carbohydrate and protein availability. J. Anim. Sci. 70, 3562-3577. [ Links ]
Sundrum, A., 2015. Metabolic disorders in the transition period indicate that the dairy cows' ability to adapt is overstressed. Animals 5, 978-1020. [ Links ]
Uddin, M.d.J., Khandaker, Z.H., Khan, M.d.J. & Khan, M.M.H,. 2015. Dynamics of microbial protein synthesis in the rumen - A review. Uddin. Ann. Vet. Ani. Sci. 2, 117-131. [ Links ]
Valente, T.N.P., Detmann, E. & Sampaio, C.B., 2015. Review: Recent advances in evaluation of bags made from different textiles used in situ ruminal degradation. Can. J. Anim. Sci. 95, 493-498. [ Links ]
Van Soest, P.J., 1994. Nutritional ecology of the ruminant. 2nd ed. Cornell University Press, Ithaca, USA. [ Links ]
Received 12 June 2017
Accepted 19 October 2017
First published online 23 February 2018
# Corresponding author: juanachagas@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}