










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.3 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i3.3
ARTICLES
Effect of herbal choline and rumen-protected methionine on lamb performance and blood metabolites
V. Rodriguez-GuerreroI; A. C. LizarazoII, #; S. FerraroI; N. SuárezIII; L. A. MirandaIV; G. D. MendozaI
IDoctorado en Ciencias Agropecuarias, Universidad Autónoma Metropolitana Xochimilco, Mexico
IICentro de Ensenanza Practica e Investigación en Producción y Salud Animal (CEPIPSA), Facultad de Medicina Veterinaria y Zootecnia, Universidad Nacional Autónoma de Mexico
IIIMaestría en Ciencias Agropecuarias, Universidad Autónoma de San Luis Potosí, Mexico
IVUniversidad Autónoma Chapingo, Departamento de Zootecnia, Mexico
ABSTRACT
Twenty-four lambs (Pelibuey x East Friesian), weighing 22.7 ± 3.2 kg, were fed a basal diet of corn silage, oat hay, alfalfa hay, and concentrate (60% forage and 40% concentrate). Treatments consisted of oral doses of rumen-protected methionine (RPM) (0 and 1.5 g/day) and herbal choline (biocholine) (0 and 4 g/day) in a completely random block design with factorial arrangement of treatments, where lambs were blocked by sex. The experiment was conducted for 60 days, and measurements of live weight and dry matter intake were obtained. No effects of the treatments were observed on performance variables (lamb growth, consumption and feed conversion). Non-esterified fatty acids (NEFA) were increased by biocholine and unaffected by methionine (Met). Biocholine increased glucose and cholesterol, whereas methionine increased triglycerides, albumin and plasma protein. The dietary supplementation with biocholine and RPM did not improve lambs' growth; however, biocholine and Met showed a lipotropic effect by mobilizing NEFA and stimulating glucose and cholesterol synthesis.
Keywords: Feed additives, growing sheep, lipid metabolism
Introduction
Methionine (Met) has been recognized as one of the limiting amino acids for growth and milk production in goats and sheep (NRC, 2007). However, its optimal intake for various physiological states is unknown. Previous experiments using rumen-protected methionine (RPM) in growing lambs have not been successful, presumably because the diets were based on low-quality forage with inadequate lysine (Lys) and Met flows (Sanchez et al., 2012). In contrast, grazing heifers supplemented with RPM and complemented with sources of bypass Lys improved average daily gain (ADG) (Gomez et al., 2011). The NRC (2007) indicated that choline requirements have not been established for sheep, but suggested that production could be improved by using it as a supplement. In previous experiments, finishing lambs increased their ADG when they were provided with rumen-protected choline (RPC) during the first stages of growth (Bryant et al., 1999). Recently, the addition of herbal choline improved the final live weight in lambs fed with high grain rations (Godinez-Cruz et al., 2015).
There is a known relationship between the metabolisms of choline and Met. The two are metabolically interchangeable as sources of methyl groups, and play key roles in the energy and protein metabolism in ruminants (Pinotti et al., 2002). Methionine can provide methyl groups for the de novo synthesis of choline (via S-adenosylmethionine). Choline can be oxidized to betaine, with methyl transfer from homocysteine and then Met (Zeisel & Da Costa 2009; Ardalan et al., 2011). Maskal'ová et al. (2014) suggest the importance of evaluating RPM and choline in ruminants and experiments in various physiological statuses. Previous experiments with dairy cows showed that dietary supplementation with RPM and RPC alleviated negative energy balance, thereby improving feed intake and modulating hepatic lipid metabolism (Sun et al., 2016). Another study showed that the combination of RPM and RPC did not improve milk production, but it did result in a moderate interaction in the energy balance, reducing the megacalories (Mcal) lost per day (Zhou et al., 2016). The results obtained in rats ingesting a choline-deficient diet had lower tissue concentrations of Met and S-adenosylmethionine (Zeisel et al., 1989), suggesting that there could be advantages in using both nutrients in growing ruminants. It is thus possible to hypothesize that lamb growth can be improved by the supplementation of Met and choline. Therefore, the objective of this experiment was to evaluate the dietary supplementation of a combination of RPM and biocholine on lambs' growth and their blood metabolites as an indicator of lipid metabolism. Both nutrients have liver functions that affect the lipid and carbohydrate metabolism. Met and choline play important roles in methyl group transfer (Niculesco & Zeisel, 2000) and methyl groups are required for carnitine synthesis essential for fatty acid oxidation (Vaz & Wanders, 2002). Phosphatidylcholine participates in the synthesis and export of triglycerides in very low density lipoproteins (Zeisel, 2006) and this has affected plasma NEFA and cholesterol (Pinotti et al., 2003). Choline has increased liver glycogen (Piepenbrink & Overton, 2003) and Met transamination pathways promote the activation of the transcriptional coactivator PGC-Ια, which is involved in the control of hepatic gluconeogenesis (Tavares et al., 2016); PGC-Ια plays a central role in the regulation of cellular energy metabolism, (Liang & Ward, 2006).
Materials and Methods
The experiment was conducted in the Centro de Ensenanza Practica e Investigación en Producción y Salud Animal (CEPIPSA) Veterinary Faculty of the Universidad Nacional Autónoma de Mexico (19° N, 99° W, 2760 m above sea level) with a mean temperature of 19 °C.
Twenty-four lambs (Pelibuey x East Friesian: 22.7 ± 3.2 kg LW; 12 males and 12 females) were assigned to four treatments, which consisted of two oral doses of RPM (Mepron, Evonik Industries Mexico Co) at 0 and 1.5 g/day and biocholine (Indian Herbs and Technofeed Mexico) at 0 and 4 g/day in a completely random block design with factorial arrangement (2 x 2) where lambs were blocked by sex. The doses of RPM and biocholine were weighed daily and administered orally to each animal before the first meal to ensure consumption. The diet was formulated according to NRC (2007) requirements for growing lambs. After an adaptation period of 15 days, lambs were fed with a basal diet of alfalfa hay (30%), corn stover (15.0%), oat hay (8.0%), cane molasses (5.0%), commercial concentrate (40.0% Ovina Engorda MP) and minerals (1.0%) (Each kg contained Ca 115 g, P 80 g, Mg 20 g, Na 150 g, Cl 230, K 5 g, S 40 g, Mn 2000 mg, Fe 5500 mg, Zn 6000 mg, Se 30 mg, Co 50 mg, I 100 mg, Cu 1000 mg, vitamin A 500,000 IU, vitamin D 150,000 IU and vitamin E 1000 IU.) Lambs were fed in individual pens at 9:00 and 14:00 h, ensuring 100 g of feed refusals per day, and they had access to clean water ad libitum. The experiment lasted 60 days.
Dry matter intake (DMI) was measured on five consecutive days from day 55 to 60. Bodyweight was recorded at 1, 40, and 60 days. Samples of feed and feed refusals were collected daily during the collection period. All samples were frozen at -20 °C until analyses. Feed samples were analysed according to the AOAC (1995) methods for dry matter (DM) and crude protein. Neutral detergent fibre (NDF) and acid detergent fibre (ADF) analyses were conducted according to Van Soest et al. (1991). Faecal samples were collected directly from the rectum every four hours for five consecutive days (Van Cleff et al. (2016), starting on day 50 of the experimental period, to estimate the apparent dry matter digestion. Feed and orts were collected daily during the same period, and those samples were used to determine acid insoluble ash as an internal marker to estimate dry matter digestibility (DMD) (Van Keulen & Young, 1977). Samples of biocholine, containing choline in the form of phosphatidylcholine, were also analysed according to the procedures described previously (ADF, NDF, DM and crude protein). The amino acid content of feed was analysed at Amino Lab, Evonik Industries (AOAC International, 1995) (European Commission, 2009).
Blood samples were collected via jugular venipuncture with vacutainer tubes without anticoagulant at the end of the experiment (day 60) to determine the changes in serum metabolites. The serum was separated by centrifugation (3000 rpm for 30 min) and refrigerated until analysis. Glucose, cholesterol, triglycerides, albumin, total protein and NEFA were determined with a spectrophotometer Cary-100 UV-visible with these methods, namely glucose oxidase-peroxidase (GOD-POD), cholesterol-peroxidase (CHOD-POD), glycerol phosphate dehydrogenase-peroxidase (GPO-POD), Bromocresol green, Biuret method according to Spinreact (Spain) kits, and NEFA by Randox Kit, respectively.
To estimate the in vitro degradation of the biocholine and to demonstrate the protection against the microbial activity, samples of biocholine (500 mg) were incubated in 100 mL amber flasks to estimate the kinetics of gas production with ruminal fluid. The inoculum consisted of rumen liquor obtained as described by Mendoza et al. (1995), using an oesophageal probe from two sheep (34 ± 1.6 kg BW) fed a 50: 50 forage to concentrate ratio. The inoculum was obtained before the morning feeding, and was mixed and strained through eight layers of cheesecloth into a flask flushed with CO2. Then, 10 mL of particle-free ruminal fluid was added to each glass bottle (120 mL capacity), and 80 mL of buffer solution (35 g sodium bicarbonate and 4 g ammonium bicarbonate for 1 L distilled water) (Goering & Van Soest, 1970) was added under a continuous flow of carbon dioxide to maintain anaerobic conditions. Each flask was closed tightly with a rubber stopper and an iron sealing ring and incubated in a water bath at 38 °C. The gas pressure was measured with a pressure gauge (Metron, Model 63100, México) at 0, 2, 4, 6, 8, 10, 14, 18, 24, 30, 36, 42, 48, 60, and 72 hours of incubation (Blümmel & Lebzien, 2001). Pressure values were transformed to gas volumes with the following linear regression equation:

Using the gas volumes at each time fraction, parameters of kinetics of gas production were estimated as follows: lag phase (h), maximal volume (Vm; mL g-1 DM of substrate), and rate (S; h-1) of gas production, from the model proposed by Menke & Steingass (1988):

The fermentation residues were dried at 65 °C overnight before recording. The half time (K0.5 h) required to obtain the Vmax was estimated using the 0.639/k of a first-order kinetics relationship.
Data were analysed as a completely randomized block design using sex as a blocking criterion and the interaction block x treatment as an error term. If the response variable was measured more than once, the data were analysed using repeated analyses (Herrera & Garcia, 2010) with JMP® SAS institute Inc., Cary N.C. (Sall et al., 2012).
Results and Discussion
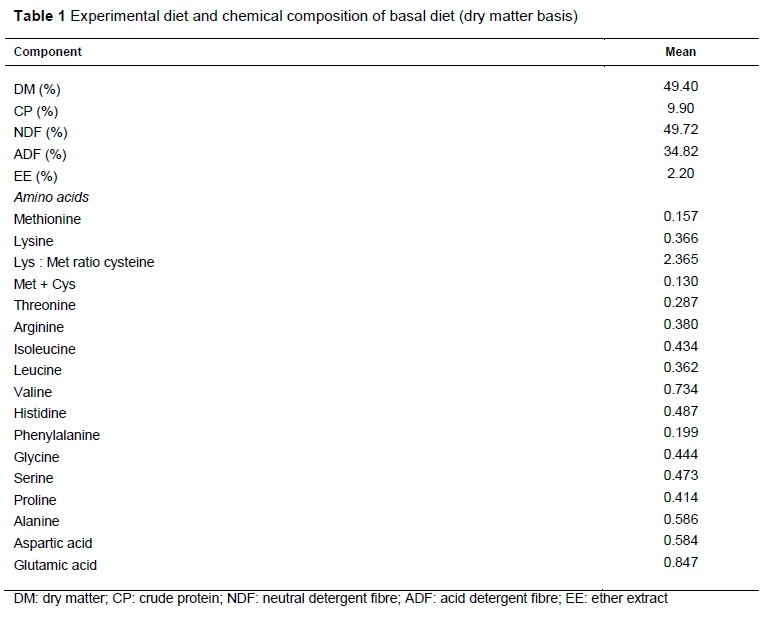
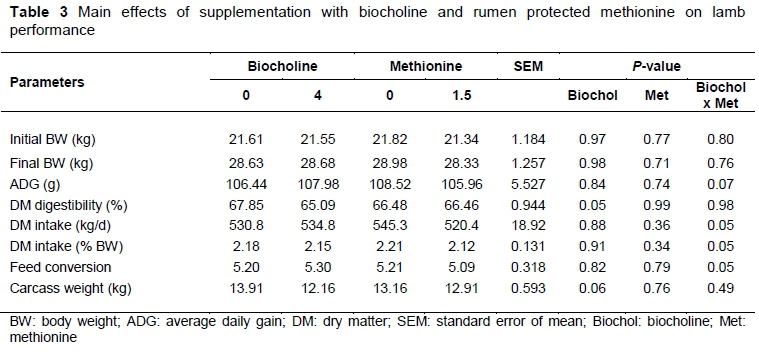
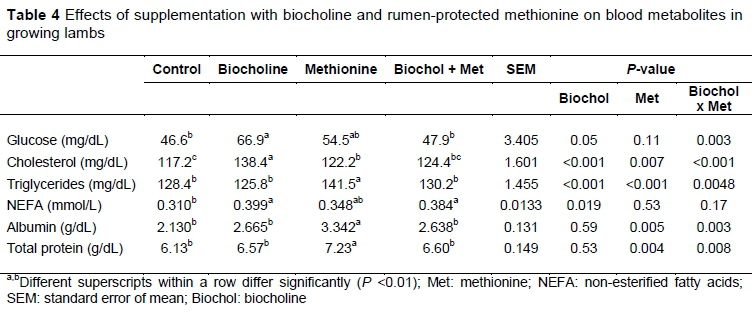
The chemical composition of basal rations is presented in Table 1, highlighting the low content of Lys and Met and their relationship. Table 2 presents the biocholine chemical composition and the parameters of in vitro gas production with ruminal microorganism, highlighting the high lipid content. Regarding lamb performance (Table 3), no treatment effects were observed on bodyweight, DM intake or digestibility. Table 4 shows the blood metabolites. The NEFAs were increased by biocholine (P <0.01) and unaffected by Met. Other blood metabolites, such as triglycerides, showed the interactions of Met and biocholine (P <0.01). Biocholine increased glucose and cholesterol (P <0.01), whereas Met increased triglycerides, albumin and plasma protein.
It has been suggested that the response to RPM and RPC may be positive in dairy cattle because of their high methylation requirements (Brusemeister & Sudekum, 2006), but their effects on production have not been consistent (Davidson et al., 2008; Sun et al., 2016; Zhou et al., 2016). In this experiment, the lack of response in lamb growth to RPM may have been owing to the low Lys content in the basal diet (Table 1). The basal diet in this experiment had a theoretical duodenal flow of metabolizable Lys of 0.2 g/d for treatments without RPM and 0.05 g/d of Met without amino acid. Therefore, when RPM was added, the Lys : Met ratio decreased to 0.13, and Lys limited the growth response in both groups. Sanchez et al. (2012) evaluated 5 g/d of RPM in growing lambs fed with rations based on corn stover and did not find any effects of the amino acid on growth, intake or feed efficiency. In contrast, Gomez et al. (2011) demonstrated that grazing heifers exhibited improved growth in response to the dietary addition of RPM when a source of bypass Lys was included in the supplement. Madsen et al. (2005) found a positive effect in dairy goats in milk production, but only when Met was combined with rumen-protected Lys at a Lys to Met ratio of 2.55 : 1.Baldwin et al. (1993), in experiments with lambs and ewes, demonstrated that neither lactating ewes nor lambs responded to RPM supplementation in diets with extensive protein degradation, which resulted in a limited Lys supply.
The in vitro gas production indicates that half of herbal choline is fermented at 18 hours (Table 2), which would potentially allow an important fraction in the lower tract. However, in vitro incubations reflect only bacterial activity and since in vivo rumen protozoa may be responsible for the engulfment of phosphatidylcholine in the rumen (Mohamed & Chaudhry 2008), ciliates should be evaluated in future studies and confirm the duodenal flow of the herbal choline and ensure the adequate supply of this nutrient in growing ruminants. The ruminal degradation of RPM (Mepron) in vitro and in situ was reported by Lara et al. (2003), whose results for lactating cows confirm the protection (Lara et al., 2006). However, information for herbal choline is scarce. According to the parameters of gas production (Table 3) and the results of Godinez-Cruz et al. (2015), a positive response in lamb growth was expected. However, no effects of biocholine or biocholine-Met supplementation were observed in the lamb performance variables mainly because of the inadequate Lys : Met ratio in the diet.
The results obtained for the half time to reach Vmax suggest that the biocholine source contains fractions of rumen-protected choline. An evaluation of commercial sources of RPC indicated that in vitro or in situ degradation in rumen from encapsulated products varied from 40 to 80% (Brusemeister & Sudekum, 2006; Jayaprakash et al., 2016). The results of Godinez-Cruz et al. (2015), who compared biocholine and an encapsulated choline chloride, indicated that the two sources resulted in the same final degradation, providing more bypass choline than that observed for the unsupplemented control.
The responses in blood metabolites have been varied according to the literature, and the estimated dose per metabolic weight of intestinally available choline appears not to be the determining factor. In this research, some metabolites (triglycerides and NEFA) were affected at a dose of just 6.42 mg/kg BW075, but Li et al. (2015) did not find any such effects in blood cholesterol and triglycerides in lambs receiving an estimated dose from 37 to 116 mg/kg BW075. In dairy cattle, Pinotti et al. (2004) observed a significant increase of plasma cholesterol and a NEFA decrease with a tendency for increased glucose with an estimated effective duodenal dose of 75 mg/kg BW075. In contrast, Chung et al. (2009) did not find an effect on plasma NEFA and glucose in dairy cows dosed with an estimated effective 73 or 147 mg/kg BW0.75. Nor did Zhou et al. (2016), who observed changes in glucose or fatty acids with estimated flows from 72 or 80 mg/kg BW075. The supply of other methyl donors such as folate and betaine (Maio et al., 2000) and the methyl requirements of the physiological stage (Pinotti et al., 2005) must be considered in future studies of choline and Met to obtain better understanding of ruminants' responses to those nutrients.
The higher concentrations of triglycerides and NEFA observed in the present study can be explained by the lipotropic role of choline. Choline is a lipotropic factor and helps optimize the balance between fat protein syntheses in dairy cattle (Sales et al., 2010). It has been demonstrated that RPC supplementation in dairy cows before parturition decreased liver NEFA accumulation and increased liver glycogen (Pinotti et al., 2005). Choline is also necessary for the transport and metabolism of lipid cholesterol (Zeisel & Da Costa, 2009), which would explain the higher blood cholesterol concentrations. The mechanism by which hepatic cholesterol synthesis is stimulated by choline in ruminants is not fully understood, but in other species, choline is required to synthesize the phosphatidylcholine portion in the very low density lipoprotein (VLDL) (Yao & Vance, 1988). However, the lack of an effect on cholesterol in treatments with both choline and Met was unexpected because choline can be used for cholesterol synthesis, as shown in goats (Emmanuel & Kennelly, 1984), and because RPC may spare Met for milk.
Nutrigenomic studies have shown that RPM alters liver key enzymes related to protein and energy metabolism. Some of these enzymes involved in the Met cycle are Met adenosyltransferase (1A MAT1a), and phosphatidylethanolamine methyltransferase (PEMT). Supplementation with RPC in cows has demonstrated that hepatic gene expression associated with antioxidant synthesis and DNA methylation status were improved (Osorio et al., 2014) and studies with liver hepatocytes from rodents show the relationship among the Met and hepatic glucose synthesis through PGC-Ια (Tavares et al., 2016). The relationship between gluconeogenic factors and methylation affected by nutrition levels (Rattanatray et al., 2014) deserves more attention in ruminant studies.
The RPC, when given at lower doses (Li et al., 2015), stimulated the expression of lipoprotein lipase and fatty-acid synthase genes in muscle, but at higher doses, it showed adverse effects on the expression of some acetyl-CoA carboxylase genes. Future studies with both choline and Met that consider the gene expression of key liver enzymes and their regulating factors are required to understand the regulation of cholesterol in ruminants (Viturro et al., 2009).
Conclusions
The RPM and biocholine supplementation did not improve lamb performance in the growing phase. However, both supplements showed beneficial effects in liver metabolism, as reflected in the blood levels of NEFA, glucose and cholesterol, demonstrating that mobilization of lipids exists. More experiments are required to establish the doses of biocholine or RPM to improve lamb growth.
Acknowledgments
The authors thank TechnoFeed, México, Nuproxa, Switzerland, and Indian Herbs Co for donating herbal products and Evonik Industries, México Co. for supplying the rumen-protected methionine. This research was partially supported by Grant no. 38153 CONACYT.
Authors' contributions
GDM, ACL, and VRG conceived and designed the experiments. VRG, ACL, SF, NS and LM performed the experiment and supervised the experimental part related to in vitro gas production. VRG, ACL, and GDM analysed the data. GDM, ACL, SF, and LM contributed reagents, materials and analysis tools. VRG, ACL, GM contributed to the writing of the manuscript. GDM, ACL, and VRG carried out critical reading and drafting of the manuscript. All the authors agreed with the final version to be submitted. They wish to thank MSc Nayelly Sánchez for technical assistance.
Conflict of interest declaration
The authors certify that they have no affiliations with any organization or entity with any financial or non-financial interest in the subject matter or materials discussed in this manuscript.
References
AOAC, 1995. Official methods of analysis. 15th edition. Association of Official Analytical Chemists, Inc., Arlington, Virginia, USA. [ Links ]
Ardalan, M., Dehghan-Banadaky, V., Rezayazdi, K. & Hossein-Zadeh, N.G., 2011 The effect of rumen-protected methionine and choline on plasma metabolites of Holstein dairy cows. J. Agr. Sci. 149, 639-646. doi:10.1017/S0021859610001292. [ Links ]
Baldwin, J.A., Horton, G.M.J., Wohlt, J.E., Palatini, D.D. & Emanuele, S.M., 1993. Rumen-protected methionine for lactation, wool and growth in sheep. Small Rumin. Res. 12, 125-132. [ Links ]
Blümmel, M. & Lebzien, P., 2001. Predicting ruminal microbial efficiencies of dairy ration by in vitro techniques. Livest. Prod. Sci. 68, 107-117. [ Links ]
Brusemeister, F. & Sudekum, K.H., 2006. Rumen-protected choline for dairy cows: The in situ evaluation of a commercial source and literature evaluation of effects on performance and interactions between methionine and choline metabolism. Anim. Res. 55, 93-104. doi.org/10.1051/animres:2006002. [ Links ]
Bryant, T.C., Rivera, J.D., Galyean, M.L., Duff, G.C., Hallford, D.M. & Montgomery, T.H., 1999. Effects of dietary level of ruminally protected choline on performance and carcass characteristics of finishing beef steers and on growth and serum metabolites in lambs. J. Anim. Sci. 77, 2893-2903. [ Links ]
Chung, Y.H., Brown, N.E., Martinez, C.M., Cassidy, T.W. & Varga, G.A., 2009. Effects of rumen-protected choline and dry propylene glycol on feed intake and blood parameters for Holstein dairy cows in early lactation. J. Dairy. Sci. 92, 2729-2736. doi:10.3168/jds.2008-1299. [ Links ]
Davidson, S., Hopkins, B.A., Odle, J., Brownie, C., Fellener, V. & Whitlow, W., 2008. Supplementing Limited methionine diets with rumen-protected methionine, betaine and choline in early lactation Holstein cows. J. Dairy. Sci. 91, 1552-1559. [ Links ]
Emmanuel, B., & Kelly, J.J., 1984. Kinetics of methionine and choline and their incorporation into plasma lipids and milk components in lactating goats. J. Dairy Sci. 67, 1912-1918. doi.org/10.3168/jds.S0022-0302(84)81524-6. [ Links ]
European Commission, 2009. 2009/150/EC Commission regulation laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union L 54,1-130. [ Links ]
Godinez-Cruz, J., Cifuentes-López, O., Cayetano, J., Lee-Rangel, H., Mendoza, G., Vazquez, A. & Roque, A., 2015. Effect of choline inclusion on lamb performance and meat characteristics. J. Anim. Sci. 93, 766, Suppl 3. [ Links ]
Goering, H.K. & Van Soest, P.J., 1970. Forage fiber analysis apparatus, reagents, procedures, and some applications. In H.K. Goering, & P.J. Van Soest (eds). Agriculture handbook, 379. Agricultural Research Service, United States Department of Agriculture. Washington DC, USA. [ Links ]
Gomez, A., Mendoza, G.D., García-Bojali, C., Barcena, R., R, J.A., Crosby, M.M., Pinos, J.M. & Lara, A., 2011. Effect of supplementation with urea, blood meal, and rumen-protected methionine on growth performance of Holstein heifers grazing kikuyu pasture. Trop. Anim. Health. Prod. 43(3), 721-724. [ Links ]
Herrera, A.J.G. & Garcia, C., 2010. Bioestadística en Ciencias Veterinarias, Procedimientos de Análisis de datos en SAS. Edn. Universidad Complutense de Madrid. Madrid, Espana (in Spanish). [ Links ]
Jayaprakash, G., Sathiyabarathi, M., Robert, M.A. & Tamilmani, T., 2016. Rumen-protected choline: a significance effect on dairy cattle nutrition. Vet. World. 9, 837-841. doi:10.14202/vetworld.2016.837-841. [ Links ]
Lara, B.A., Mendoza, G.D., Bárcena, M.R. & Landois, L.P., 2003. Degradabilidad ruminal in situ e in vitro de la metionina protegida. Tec. Pecu. Mex. 41, 91-103 (in Spanish). [ Links ]
Lara, B.A., Mendoza, G.D., Landois, L.A., Bárcena, R., Sanchez, T., Rojo, R., Ayala, J. & Vega, S., 2006. Milk production in Holstein cows supplemented with different levels of ruminally protected methionine. Livest. Sci. 105, 105-108. [ Links ]
Li, H., Wang, H., Yu, L., Wang, M., Liu, S., Sun, L. & Chen, Q., 2015. Effects of supplementation of rumen-protected choline on growth performance, meat quality and gene expression in longissimus dorsi muscle of lambs. Arch. Anim. Nutr. 69, 340-350. [ Links ]
Liang, H, & Ward, W.F., 2006.PGC-1alpha: a key regulator of energy metabolism. Adv Physiol Educ. Dec: 30(4), 145-51. [ Links ]
Madsen, T.G., Nielsen, L. & Nielsen, M.O., 2005. Mammary nutrient uptake in response to dietary supplementation of rumen protected lysine and methionine in late and early lactating dairy goats. Small. Rumin. Res. 56, 151-164. [ Links ]
Maio, R., Dichi, J.B. & Burini, R.C., 2000. Implicacöes do alcoolismo e da doenca hepática crönica sobre o metabolismo de micronutrientes. Arq. Gastroenterol. 37, 120-124 (in Portugese). [ Links ]
Maskal'ová, I., Vajda, V., Krempaský, M.& Bujnák, L., 2014. Rumen degradability and ileal digestibility of proteins and amino acids of feedstuffs for cows. Acta. Vet. Brno. 83, 225-231. doi:10.2754/avb201483030225. [ Links ]
Mendoza, M.G.D., Britton, R.A. & Stock, R.A., 1995. Effect of protozoa and urea level on in vitro starch disappearance and amylolytic activity of ruminal microorganisms. Anim. Feed. Sci. Tech. 54, 315-325. [ Links ]
Menke, K. & Steingass, H., 1988. Estimation of the energetic feed value obtained from chemical analysis and in vitro gas production using rumen fluid. Anim. Res Dev. 28, 9-52. [ Links ]
Mohamed, R. & Chaudhry, A.S., 2008. Methods to study degradation of ruminant feeds. Nutr. Res. Rev. 21, 68-81. doi:10.1017/S0954422408960674. [ Links ]
Niculescu, M.D., Zeisel, S.H., 2002. Diet methyl donors and DNA methylation: Interactions between dietary folate methionine and choline. J. Nutr. 132, 2333S-2335S. [ Links ]
NRC, 2007. Nutrient requirements of small ruminants: Sheep, goats, cervids, and New World camelids. National Academic Press, Washington DC, USA. [ Links ]
Osorio, J.S., Ji, P., Drackley, J.K., Luchini, D. & Loor, J.J. 2014. Smartamine M and MetaSmart supplementation during the peripartal period alter hepatic expression of gene networks in 1-carbon metabolism, inflammation, oxidative stress, and the growth hormone-insulin-like growth factor 1 axis pathways. J. Dairy Sci. 97, 7451-7464. doi: 10.3168/jds.2014-8680. [ Links ]
Piepenbrink, M.S., Overton T.R., 2003. Liver metabolism and production of cows fed increasing amounts of rumen- protected choline during the periparturient period. J. Dairy. Sci. 86, 1722-1733. [ Links ]
Pinotti, L., Baldi, A., Dell'Orto, V., 2002. Comparative mammalian choline metabolism with emphasis on the high yielding dairy cow. Nutr Res Rev 15: 315-332. doi: 10.1079/NRR200247. [ Links ]
Pinotti, L., Baldi A., Politis I., Rebucci R., Sangalli L., Dell'Orto V., 2003. Rumen-protected choline administration to transition cows: Effects on milk production and vitamin E status. J. Vet. Med. A Physiol. Pathol. Clin. Med. 50, 18-21. [ Links ]
Pinotti, L., Campagnoli, A., Sangalli, L., Rebucci, R., Deii'Orto, V. & Baldi, A., 2004. Metabolism of periparturient dairy cows fed rumen-protected choline. J. Anim. Feed. Sci. 13, 551-554. [ Links ]
Pinotti, L., Campagnoli, A., Dell'Orto, V. & Baldi, A., 2005. Choline: Is there a need in the lactating dairy cow? Livest. Prod. Sci. 98, 149-152. [ Links ]
Rattanatray, L., Muhlhausler, B.S., Nicholas, L.M., Morrison, J.L., & McMillen, I.C., 2014. Impact of maternal overnutrition on gluconeogenic factors and methylation of the phosphoenolpyruvate carboxykinase promoter in the fetal and postnatal liver. Pediatric. Res. 75(1), 14-21. [ Links ]
Sales, J., Homolka, P. & Koukolová, V., 2010. Effect of dietary rumen-protected choline on milk production of dairy cows: A meta-analysis. J. Dairy. Sci. 93, 3746-54. doi: 10.3®68/jds.2010-3106. [ Links ]
Sall, J., Lehman, A., Stephens, M. & Creighton, L. 2012. JMP® start statistics: A guide to statistics and data analysis (5th ed.). SAS Institute Inc: Cary, NC, USA. [ Links ]
Sanchez, A.E., Ortega, C.M.E., Mendoza, M.G., Montanez, V.O.D. & Buntinx, D.S.E., 2012. Rastrojo de maíz tratado con urea y metionina protegida en dietas para ovinos en crecimiento. Interciencia. 37, 395-399 (in Spanish). [ Links ]
Sun, F., Cao, Y., Cai, C., Li, S., Yu, C. & Yao, J., 2016. Regulation of nutritional metabolism in transition dairy cows: Energy homeostasis and health in response to post-ruminal choline and methionine. PLOS ONE 11,8, e0160659 doi:10.1371/journal.pone.0160659. [ Links ]
Tavares, C.D.J., Sharabi, K., Dominy, J.E., Lee, Y., Isasa, M., Orozco, J. M. & Puigserver, P., 2016. The Methionine transamination pathway controls hepatic glucose metabolism through regulation of the GCN5 acetyltransferase and the PGC-Ια transcriptional coactivator. J. Biol. Chem. 291(20), 10635-10645. doi: 10.1074/jbc.M115.706200. [ Links ]
Van Cleef, F.O.S., Ezequiel, J.M.B., D'Aurea, A.P., Almeida, M.T.C., Perez, H.L., & Van Cleef, E.H.C.B., 2017. Feeding behavior, nutrient digestibility, feedlot performance, carcass traits, and meat characteristics of crossbred lambs fed high levels of yellow grease or soybean oil. Small. Rumin. Res. 137, 151-156. [ Links ]
Van Keulen, J.V. & Young, B.A., 1977. Evaluation of acid-insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci. 44, 282-287. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Vaz, M.F & Wanders, R.J.A., 2002. Carnitine biosynthesis in mammals. Biochem. J. 361, 417-429. [ Links ]
Viturro, E., Koenning, M., Kroemer, A., Schlamberger, G., Wiedemann, S., Kaske, M. & Meyer, H.H., 2009. Cholesterol synthesis in the lactating cow: Induced expression of candidate genes J. Steroid. Biochem. 115, 62-67. [ Links ]
Yao, Z. & Vance, D.E., 1988. The active synthesis of phosphatidylcholine is required for very low density lipoprotein secretion from rat hepatocytes. J. Biol. Chem. 263, 2998-3004. [ Links ]
Zeisel, S.H., Zola, T., Da Costa, K., & Pomfret, E.A., 1989. Effect of choline deficiency on S-adenosylmethionine and methionine concentrations in rat liver. Biochem. J. 259, 725-729. [ Links ]
Zeisel, S.H., 2006. Choline: Critical role during fetal development and dietary requirements in adults. Annual Review of Nutrition2,1, 229-250. [ Links ]
Zeisel, S.H. & Da Costa, K.A., 2009. Choline: An essential nutrient for public health. Nutr Rev. 67, 615-623. doi:10.1111/j.1753-4887.2009.00246.x [ Links ]
Zhou, Z., Vailati-Riboni, M., Trevisi, E., Drackley, J.K., Luchini, D.N. & Loor, J.J., 2016. Better postpartal performance in dairy cows supplemented with rumen protected methionine compared with choline during the peripartal period. J. Dairy Sci. 99, 1-17. doi: http://dx.doi.org/10.3168/jds.2015-10525 [ Links ]
Received 4 August 2017
Accepted 3 January 2018
First published online 30 January 2018
# Corresponding author: lizarazo@unam.mx


{kind=link}
{kind=link}
{kind=link}
{kind=link}