










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.48 n.2 Pretoria 2018
http://dx.doi.org/10.4314/sajas.v48i2.13
Effects of grape pomace and vitamin E on performance, antioxidant status, immune response, gut morphology and histopathological responses in broiler chickens
S. K. EbrahimzadehI; B. NavidshadI, #; P. FarhoomandII; F. Mirzaei AghjehgheshlaghI
IDepartment of Animal Science, University of Mohaghegh Ardabili, Ardabil, Iran
IIDepartment of Animal Science, Urmia University, Urmia, Iran
ABSTRACT
We investigated the effects of grape pomace (GP) and vitamin E on the performance, antioxidant status, immune response, gut morphology and histopathological responses in broiler chickens. Two hundred and fifty male broiler chickens (Ross 308) were allocated to five dietary treatments (50 birds/treatment) in a completely randomized design. The experimental diets were as follows: i) Control corn-soybean meal diet (C); ii) C + Vitamin E (200 mg/kg of α-tocopherol acetate); iii) The diet containing 5%GP; iv) The diet containing 7.5% GP; v) The diet containing 10% GP. No differences were found in performance of the experimental birds. Birds fed 7.5% GP showed a significant increase in superoxide dismutase (SOD), glutathione peroxidise (GSH-Px) activity and reduced malondialdehyde (MDA) concentration. Antibody titer against Newcastle disease virus (NDV) at 42 days (d) of age in the control and 7.5% GP fed groups were lower than others. The primary IgM concentration of birds fed 10% GP was higher than the birds fed the control diet, the vitamin E supplemented diet or the diet containing 5% GP. The secondary titer against sheep red blood cells (SRBC) was increased significantly in the birds fed 10% GP and the secondary IgG concentration of birds fed 10% GP was higher than the birds fed the control diet and the diets containing 5% or 7.5% GP. In duodenum part of small intestine, villus height/crypt depth ratio and muscularis thickness were decreased. The results of the present study suggest that the inclusion of up to 10% GP in diets did not adversely affect broiler chickens' performance and improved the antioxidant and immune responses of broiler chickens.
Keywords: Glutathione peroxidise, polyphenol, poultry, red blood cells α-tocopherol acetate, sheep, superoxide dismutase
Introduction
Grapes (Vitis vinifera) are one of the world largest fruit crop with annual production of 77 million metric tons (FAOSTAT, 2013). Processing of grapes for ethanol, fruit juice and wine production results in huge quantities of by-products including stems, skins, seeds and peels and these residues are about 20% of the weight of the grape used in making wine (Llobera and Canellas, 2007). These by-products are rich in a wide range of polyphenols like flavonoids, monomeric phenolic compounds, catechins, and epicatechins (Dorri et al., 2012). The grape polyphenols could act as powerful antioxidants by scavenging free-radicals and terminating oxidative reactions (Brenes et al., 2016). During the processing of grapes, economical, and ecological problems are caused by a large volume of produced residues. It is anticipated that any useful application of these by-products for maintenance of the environmental equilibrium and an economic evaluation of the raw material could be considered (Abarghuei et al., 2010).
The high polyphenol content and high level of fiber are the major limitations of using grape by-products in poultry diet. Grape pomace (GP) contains high level of fiber and polymeric polyphenols as procyanidins could be bound and precipitated both dietary and endogenous proteins (Jansman et al., 1989). However, the beneficial effects of the use of polyphenols have been shown in both in vivo and in vitro studies (Alonso et al., 2002; Torres et al., 2002; Viveros et al., 2011) due to their antimicrobial and antioxidant activity.
In previous studies, the results of grape by-product utilization on the performance of chickens have been inconsistent. For example, Hughes et al. (2005) reported that the addition of grape seed extract (GSE) at the level of 30 g/kg (90.2% total phenolics) decreased the performance of chickens, while Goni et al. (2007) and Brenes et al. (2008) stated that GP supplementation at the level of 30 g/kg (4.86% total phenolics) improved antioxidant status without any negative effect on the performance of chickens. Therefore, it is suggested that the difference in the amount of phoyphenols in GP and GSE causes a difference in results, which could bind with the diet proteins and digestive enzymes (Jansman et al., 1989).
Polyphenols extract from different plants have been successfully used to help protect against diseases caused by many pathogen (Tepe et al., 2004). Therefore, polyphenols of grape by-products could be used as a feed supplement to improve antioxidant status and immunity of birds, whereas the physiological effects of these molecules are not completely known. Earlier, degenerative changes in liver and intestine of poultry on feeding of high polyphenols diets had been reported. Significant pathological changes in most of the tissues like lungs, liver, kidney, heart, intestine, and pancreas in birds fed on 100% red sorghum have been reported by Sannamani (2002).
Vitamin E as a supplement in poultry diet plays an important role in the health and productive performances. Immune response, effective growth and development, quality of meat, negative consequences of various stresses could be affected and improved by vitamin E supplementation (Surai and Sparks, 2001; Surai, 2006; Surai & Fisinin, 2010). Many research studies and various commercial products that have been introduced in recent years revealed the need for vitamin E supplement for poultry. Recently, the replacement of synthetic vitamins with natural ingredients has been considered. Of course, the physiological and biochemical functions of these compounds, including polyphenols, should be considered. Vitamin E has a main role in antioxidant system of the body. It is located in the biological membranes, where lipid peroxidation occurs. Polyphenols and other compounds of antioxidant system can help vitamin E by recycling or decreasing free radical load (Surai ,2002). Iqbal et al. (2015) found that replacement of 75 ppm vitamin E with grape polyphenols improved antioxidant status and immune response of broilers without any negative effect on histopathology of organs. Also, antioxidant activity equal to vitamin E in broiler chickens fed diet supplemented with grape pomace concentrate (GPC) was reported by Brenes et al. (2008).
This study was designed to investigate the effects of GP and vitamin E on performance, antioxidant status, immune response, blood parameters, gut morphology and liver histopathological response in broiler chickens from 0 to 42 days of age.
Materials and Methods
Red grape pomace (containing skins, pulp, seeds) was obtained from the TATAO Corporation (Urmia-West Azerbaijan, Iran). The collected GP was mixed and used for sun-drying. Proximate composition of GP was analyzed according to procedures described by the Association of Official Analytical Chemists (AOAC, 2000) (Table 1). Total polyphenol content in GP was determined after the extraction with methanol/HCl 99/1 (v/v) as an extraction solvent by using Folin-Ciocalteu colorimetric method (Singleton & Rossi, 1965). Gallic acid was employed as a calibration standard and results were expressed as Gallic acid equivalents (mg Gallic acid/g of dried samples). GP was used as a source of dietary fiber and polyphenols in the chicken diets.
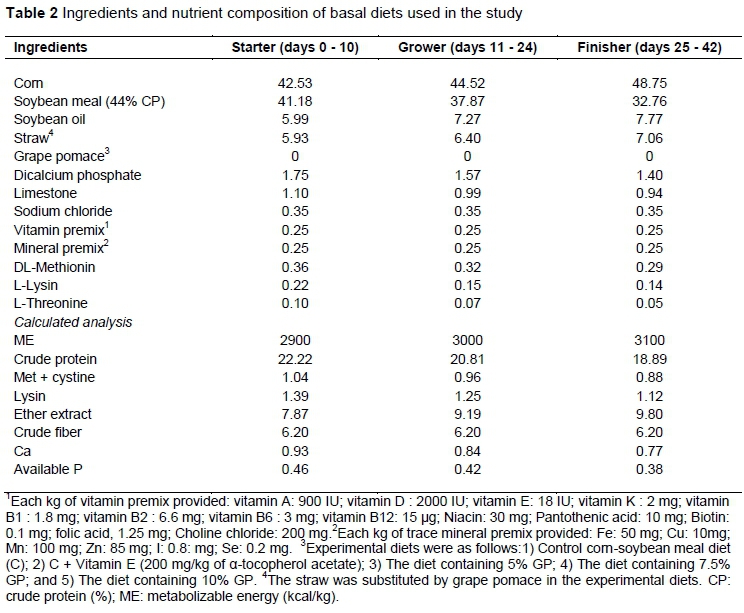
Two hundred and fifty male broiler chickens (Ross 308) at one day old were purchased from a local hatchery. All birds were allocated to five treatment groups in a completely randomized design, with 5 replicate pens/treatment and 10 chickens/pen. Dietary treatments (Table 2) in mash form were formulated to meet the nutrient requirements of broilers provided by Ross broiler's manual (Aviagen, 2014). The experimental diets were as follows: i) Control corn-soybean meal diet (C); ii) C + Vitamin E (200 mg/kg of α-tocopherol acetate); iii) Diet containing 5%GP; iv) Diet containing 7.5% GP; and v) Diet containing 10% GP. The straw was substituted by GP in the experimental diets in order to equalize the level of dietary fiber. Chicks were raised from 1 to 42 days of age and were fed starter (0 to 10 d), grower (11 to 24 d), and finisher diets (25 to 42 d). Chickens were raised in floor pens (100 × 120 cm) and had free access to feed and water for the entire experimental period (days 0 to 42). The room temperature gradually decreased from 33 to 22 °C on day 28 and then remained constant thereafter. The lighting program was a period of 20 h of light and 4 h of darkness. All experimental procedures were evaluated and approved by the animal welfare committee of the department of animal science, University of Mohaghegh Ardabili.
Birds weight gain and feed consumption were recorded for the starter (0 to 10 d), grower (11 to 24 d), and finisher (25 to 42 d) phases. The values of average daily feed intake (ADFI) and average daily gain (ADG) were adjusted for mortality rate and recorded in different phases. The feed to gain ratio (F:G) was calculated. The economic evaluation was done in terms of feed cost per kg live weight gain during the differentiation and overall study period through F:G multiplied by per kg diet price. At 42 days of age, two birds with a body weight close to the pen mean were chosen randomly from each replicate and sacrificed. The carcass, abdominal fat pad, liver, and heart were harvested, weighed and data are presented based on percent of live weight of each bird.
At 42 d of age, two blood samples of 2 birds/replicate were obtained from the wing vein following approximately 8 h fasting for the determination of serum biochemical parameters and antioxidative status. The first samples were collected into vials containing EDTA and centrifuged for 10 min at 3000×g; then plasma was collected and stored at −20 ◦ C until the analysis. The concentrations of total protein, glucose, uric acid, triglycerides (TG), total cholesterol (CHOL), high-density lipoprotein cholesterol (HDL-C), low-density lipoprotein cholesterol (LDL-C), aspartate aminotransferase (AST) and alanine aminotransferase (ALT) in plasma samples were determined using an automatic biochemical analyzer (HITACHI912.Japan)., following the instructions of the corresponding reagent kit (Audit Diagnostics Co. Ireland).
The second blood sample was collected in a heparinized graduated centrifuge tube to obtain hemolysate. This hemolysate used for evaluating antioxidant status by measuring the malondialdehyde (MDA), superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) activity. The GSH-Px activity was determined using a commercially available enzyme kit (Ransel, RANDOX/RS-504 supplied by Randox Laboratories, Crumlin, UK). The SOD activity was determined using the commercial enzyme kit (Ransod, RANDOX/SD-125 supplied by Randox Laboratories). The MDA was analyzed by using a commercial kit (Biocore Diagnostik Co.Germany), MDA was measured as a decomposed product of lipid peroxidation with 2-TBA by using the colorimetric method with a spectrophotometer at a wavelength of 532 nm (Placer et al., 1966).
On day 9 of age, Newcastle and influenza antigens were injected subcutaneously with 0.2 ml per chick with a dual vaccine of Newcastle-influenza. Also, the chicks were orally vaccinated against Newcastle disease at 19 d of age. To assay the primary and secondary antibody responses against SRBC, at 14 and 35 d of age, 2 birds/replicate were immunized intramuscularly with 0.25 ml 10% SRBC in phosphate-buffered saline (Leshchinsky & Klasing, 2001). On days 21 and 42, the same birds were bled to determine antibody titers against SRBC and Newcastle disease virus (NDV). Subsequently, antibody titers against NDV were separately measured by the hemagglutination inhibition method (HI). For the anti-SRBC antibodies measurement by the microhemagglutination technique as described by Qureshi and Havanstein (1994), serum from each sample was collected, heat inactivated at 56 °C for 30 min, and then analyzed for total, IgM and IgG (mercaptoethanol-resistant).
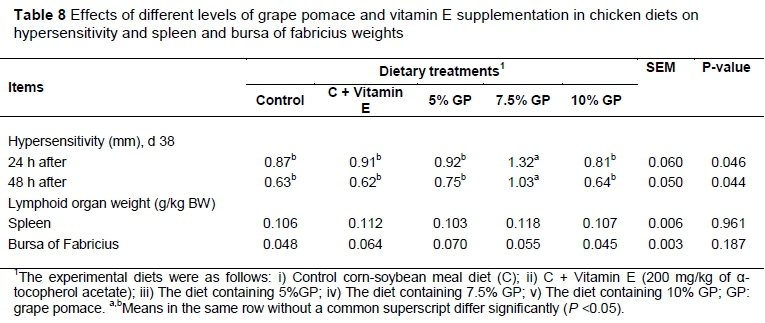
The cutaneous basophil hypersensitivity test by using phytohemagglutinin applied for Cellular immunity assessment. At d 38, the toe web between the third and fourth digits of the right foot was measured in millimeters with a constant tension micrometer. Immediately after measurement, 100 μg of phytohemagglutinin (suspended in 0.10 ml of sterile saline) was injected into the toe web. The toe web swelling was measured 24 and 48 h after injection. The response was determined by subtracting the skin thickness of the first measurement of the skin thickness of the second measurement (Corrier & DeLoach, 1990). Lymphoid organs, including the spleen and bursa of Fabricius were evaluated after slaughter on d 42 of the experiment and data are presented based on percent of live weight of each bird.
On d 42, two chicks from each replicate were sacrificed following overnight fasting to limit intestinal throughput. The whole length of the small intestine was removed, and samples were taken from different parts of the small intestine (duodenum, jejunum and ileum). The samples were gently flushed twice with physiological saline (1% NaCl) to remove intestinal contents and placed in 10% formalin for fixation. The 5-6 μ sections were made by paraffin embedding method and stained with hematoxylin-eosin. The micrometrical analysis was done using Dino-Lite Digital Microscope, Digital Dino-Lite Eye-Piece and Dino-Capture 2 software on microphotographs. The variables measured were villus height, crypt depth and thickness of the muscularis layer. The ratio of villus height to crypt depth was calculated.
After sacrificing two chicks from each replicate, samples were taken from the liver of each bird. Liver weights were measured, and all data are presented based on percent of live weight of each bird. Then the liver tissue samples were collected, and samples were fixed in 10% formalin and transmitted to the histological laboratory for the detection of any histopathological changes. The 5μ sections were made by paraffin embedding method and stained with hematoxylin-eosin. The micrometrical analysis (diameters of cells and nucleus) was done using Dino-Lite Digital Microscope, Digital Dino-Lite Eyepiece and Dino-Capture 2 software on microphotographs.
Data were analyzed in a completely randomized design using the General Linear Models (GLM) procedures of SAS (SAS Institute Inc, 2001). When the differences were significant (P <0.05), mean values between treatments were compared using the Duncan test.
Results
The effects of feeding diets containing vitamin E and increasing concentration of GP on broiler body weight (BW), ADG, ADFI and F:G are summarized in Table 3.
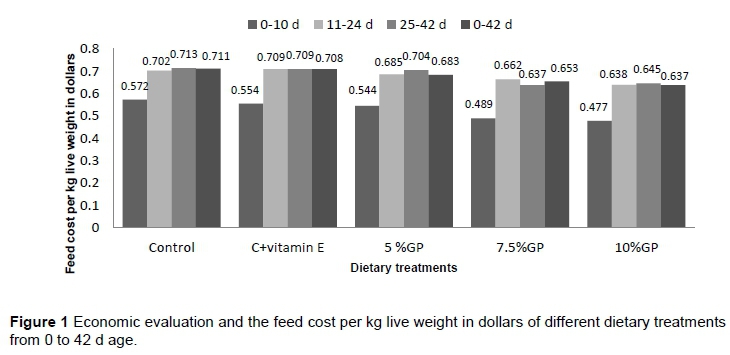
Inclusion of vitamin E and different levels of grape pomace in diets resulted in lower feed cost and cost per kg live weight gain (Figure 1).
The cost per kg live weight gain was reduced during the overall study period. The effects of experimental diets on carcass characteristics, fat, and yields of birds are presented in Table 4. Our current data indicate that GP supplementation was no significant effect on carcass, abdominal fat, liver, or heart relative weights in broilers.
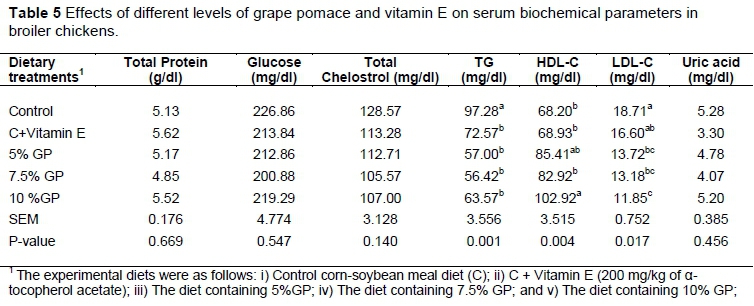
The effect of experimental diets on serum biochemical parameters is reported in Table 5. The total protein, uric acid, glucose, and cholesterol concentrations were not affected by dietary treatments. Feeding GP and vitamin E diets significantly (P <0.01) decreased serum TG and broilers fed the diet containing 7.5% GP were lowest TG concentrations (56.42 mg/dl) than in those fed the control diet (Table 5). The LDL-C levels were significantly (P <0.05) decreased in GP fed groups and broilers fed the diet containing 10% GP were lower LDL-C concentrations (11.85 mg/dl). Whereas the HDL-C level was increased significantly (P <0.01) by the dietary GP supplementation in the 10% GP fed birds in comparison with the control group.
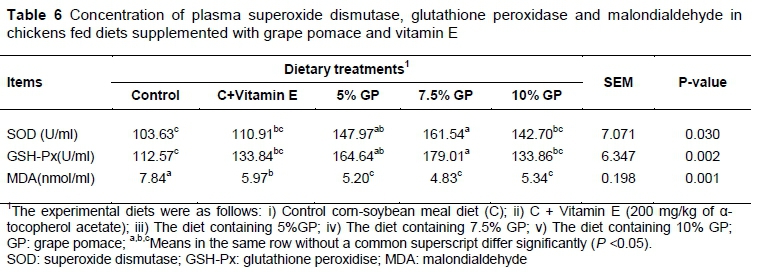
Compared to control group, birds fed supplemented diets (7.5% GP) showed an increase in GSH-Px activity up to 179.01 IU/L (P <0.01) and SOD activity up to 161.54 IU/L (P <0.05). The inclusion of GP in the diets significantly (P <0.01) reduced MDA concentration and broilers fed the diet containing 7.5% GP were lowest MDA concentrations (4.83 nmol/ml) than in those fed the control diet (Table 6). Although vitamin E supplementation improves the activity of antioxidant enzymes and decreases MDA concentration, the highest antioxidant activity and the lowest MDA concentration in birds fed with GP have been observed.
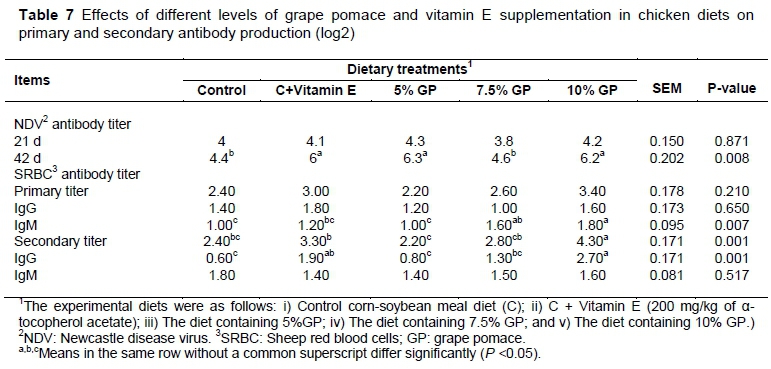
The effects of dietary treatments on antibody production, hypersensitivity and lymphoid organ weights in broilers are shown in Tables 7 and 8. The humoral and cellular immune responses could be improved by supplementing of GP to chicken diets. Antibody titers against ND virus at 42 d were higher (P <0.05) in broilers fed the diet containing GP than control group. The supplementation of GP in the chicken diets failed to have any significant impact on the anti-SRBC titers of total and IgG antibodies at 21 d of age, whereas IgM antibodies were higher (P <0.05) in broilers fed the diet containing 10% GP than control group (Table 7). In the case of secondary response, compared to other experimental groups, there was an increase (P <0.05) in total anti-SRBC titers and IgG antibodies in birds fed the 10% GP treatment (Table 7). In contrast, during the secondary challenge, the SRBC-specific IgM titer in the broiler serum showed no differences across treatments. Although vitamin E supplementation improves the antibody titers against ND virus at 42 d and total anti-SRBC titers and IgG antibodies of secondary response, the highest antibody production in birds fed with GP have been observed.
Compared to control group, toe web thickness within 24 and 48 h after injection in birds fed the diet containing 7.5% GP was increased by phytohaemagglutinin (P <0.05) (Table 8).
The effects of dietary treatment on the relative weights of immune organs in chicken at 42 d of age are reported in Table 8. In the present study,the relative immune organ weights were not significantly affected by dietary treatments (Table 8).
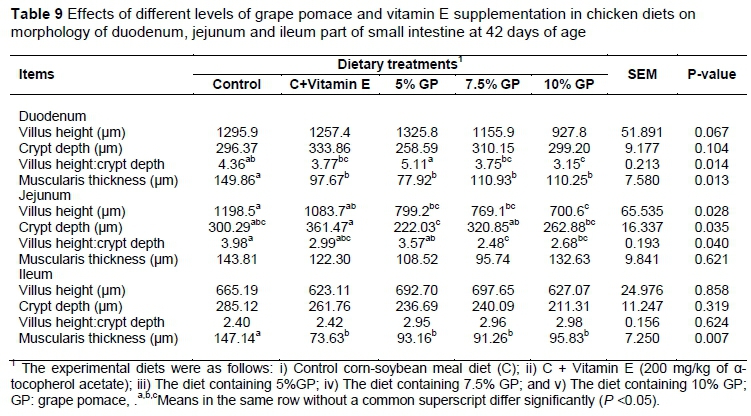
The histomorphometrical measurement results of different parts of small intestine of broiler chicks are shown in Table 9. The villus height/crypt depth ratio and muscularis thickness of chicken duodenum were decreased significantly in 5% GP fed group compared to control group (P <0.05) whereas villus height and crypt depth were not affected by experimental diets.
In jejunum parts of small intestine, villus height and crypt depth were reduced (P< 0.05) and villus height/crypt depth ratio was increased (P <0.05) in broilers fed the 5% GP diet compared with the other groups (Table 9). There was no significant effect of feeding diets on the villus height and crypt depth at the ileum of broilers (P >0.05). Muscularis thickness was decreased significantly in broilers fed vitamin E and GP comparison with the control group (P <0.01).
The effect of dietary treatment on liver histology in broilers are shown in Table 10. No significant (P >0.05) histopathological changes were seen for birds fed different levels of GP compared with the control group. The AST concentration was increased significantly in the birds fed 10% GP (P <0.05). Also, there was no significant (P >0.05) effect of feeding diets on liver relative weights in broilers (Table 4).
Discussion
References in the literature in relation to the use of grape by-products in chicken diets are limited and results are inconsistent. Growth depression in chickens fed diets containing GSE (90.2% total phenolics) has been reported by Lau and King (2003) and Hughes et al. (2005), whereas Hajati et al. (2015) stated that GSE addition at the levels of 300 mg/kg diet improved body weight of broilers. Grape by-products have potential for use as functional ingredients in animal nutrition (Brenes et al., 2008). Recent works show that GP up to 6% and GSE up to 0.25% could be used in chicken diet without any negative effect on performance (Brenes et al., 2008; Chamorro et al., 2013).
High polyphenol content and high level of fiber fraction are the major limitations of using GP in broiler diet. GP contains high level of fiber and polymeric polyphenols as procyanidins could be bound and precipitated both dietary and endogenous proteins. Polyphenolic compounds could be bound to digestive enzymes and proteins located at the luminal side of the intestinal tract and reduce apparent digestibility of protein in polyphenol-containing diets. Due to the interaction of reactive hydroxyl groups of polyphenols with the carbonyl group of protein, they can form complexes with proteins (Jansman et al., 1989; Ortiz et al., 1993). There are reports in literature which indicated that polyphenols are able to inhibit a range of enzymes including α-amylase and α-glucosidase activity (Yilmazer-Musa et al., 2012), α-glucosidase and pancreatic lipase (You et al., 2011), alpha-amylase, and alpha-glucosidase (McDougall et al., 2005). The inhibition of digestive enzymes could be explained by the ability of polymeric polyphenols to form insoluble complexes with proteins in the gastrointestinal tract (Griffiths, 1986; Horigome et al., 1988).
Therefore, nutrient digestion and the growth performance of chicken could be impaired by incorporation of GP at high doses in chicken diets. In general, many different factors, including type of compounds, its dosage and combination with other compounds, can affect the effects of polyphenol compounds on digestion, absorption and metabolism of different nutrients (Martel et al., 2010). Thus, in this study, we used different level of GP and all diets were formulated with same fiber content. Any difference in results should therefore be attributed to the polyphenol content. In the current experiment, we found that, GP contained 3.39% of total polyphenols by the Folin method. Thus, the concentration of polyphenols present in the highest concentration of the GP diet (10%) could be relatively low to produce a growth depression effect.
In this study, the cost per kg live weight gain was reduced during the overall study period. The highest economic returns were observed in the group fed 10% GP diet, which indicated that its use is beneficial for economical broiler production. In agreement with our results, Iqbal et al. (2015) reported that replacement of vitamin E with GP resulted in reduced feed cost. Our current data indicate that GP supplementation had no significant effect on carcass, abdominal fat, liver or heart weights in birds. These results are in agreement with the reports of Brenes et al. (2008) who reported that the inclusion of graded concentrations of GP concentrate did not affect the relative abdominal fat, liver, pancreas, and spleen weight. Recent results from Ebrahimzadeh et al. (2017) showed that dietary inclusion of 10%GP with or without tannase enzyme treatment did not affect chick growth performance. These findings show that the inclusion of enzymes in diets containing GP increased the amount of total polyphenol released in the intestine, although this effect was not accompanied by an increase in performance.
Like the results of this study, administration of GSE reduced blood levels of total TG, LDL-C and increased HDL-C in broilers (Hajati et al., 2015). Hajati et al. (2015) reported that GSE supplementation (150, 300, 450 mg/kg) decreased the concentration of serum glucose, whereas in this study GP supplementation to chicken diets did not have any significant impact on serum glucose. In agreement with our results, Ozgan (2008) reported that inclusion of 1.0% grape seed oil in laying hen diets did not affect the plasma glucose level. Results of the present study showed that GP supplementation decreased TG, LDL-C and increased HDL-C concentration of serum blood of broilers.
The mechanism by which dietary GP supplements affect the concentrations of plasma lipids is not fully understood. However, research has suggested that herbs and herbal products induce hypocholesterolemic effects by reducing the activity of 3-hydroxy-3-methylglutaryl-CoA reductase (HMGCoA reductase), the rate-limiting enzyme in cholesterol synthesis (Hong et al., 2012). In this study, the hypolipidemic effects of GP might not originate from their direct effect on lipid metabolism; however, it could be mediated by their effect on lowering oxidized lipid products. This effect may be associated with an increase in the plasma HDL-C concentration of the respective treatment groups. Grape by-products show their antioxidant characteristics through the attribute of the phenolic compounds to scavenge free radicals by forming complexes with metal ions and preventing the development of singlet oxygen (Yilmaz & Toledo, 2006; Surai, 2014).
In agreement with our results, an increase in blood GSH-Px activity of chronic heat-exposed broiler chickens that fed plant extracts rich in phenolic compounds has been reported by Seven et al. (2008). Lien et al. (2008) reported that layer chickens fed with citrus flavonoids showed a similar positive change in plasma MDA and SOD levels as regarded in our study. Polyphenols can show their antioxidative properties by giving hydrogen from hydroxyl groups positioned along the aromatic ring to terminate the free-radical oxidation of biomolecules (Foti et al., 1994). polyphenols also reduce α-tocopherol radicals (tocopheroxyls), activate antioxidant enzymes, inhibit oxidases and increase levels of uric acid and substances of low molecular weight. Shi et al. (2003) found that the polyphenols in grape have an antioxidant power 20 times higher than vitamin E and 50 times higher than vitamin C.
Both NF-kB and Nrf2 are redox-sensitive transcription factors and play a role in induction of phase II detoxifying-antioxidant defense mechanisms. They cope with oxidative stress through enhancing the expression of several enzymes. Sahin et al. (2010) reported that addition of Epigallocatechin-3-gallate in heat-stressed quail diets, has ability to increase antioxidant enzyme activities, inhibit NF-kB expression, and enhance Nrf2 expression.
In poultry production, it is more effective to use immune stimulators to improve immunity and decrease susceptibility to infectious disease. GP is particularly rich in a wide range of polyphenols and the most significant bioactivity of phenolic compounds from GP is antioxidant activity (Xia et al., 2010; Georgiev et al., 2014). Based on the numerous model studies in vitro, it is anticipated that molecular mechanisms of polyphenol health-promoting properties were related to their antioxidant properties (Andriantsitohaina et al., 2012). Flavonoids have ability to prevent injury caused by free radicals through many way, for example by direct scavenging of reactive oxygen species, activation of antioxidant enzymes, metal chelating activity, mitigation of oxidative stress caused by nitric oxide, reduction of α-tocopheryl radicals, inhibition of oxidases, increase in uric acid levels. and increase in antioxidant properties of low molecular antioxidants (Prochazkova et al., 2011).
Similar to our data, Iqbal et al. (2015) found that replacement of 75 ppm vitamin E with grape polyphenols showed higher (P <0.05) antibody titers against NDV in broilers. Additionally, Acamovic and Brooker (2005) reported that polyphenols and oregano essential oil have an immunostimulating activity in relation to the system of mononuclear phagocyte system, cell, and humoral immunity. Evidence from various studies suggests that the mechanisms on how the plant polyphenols exert their properties, such as inhibition of key enzymes, the modulation of cell receptors or transcription factors, as well as the perturbation of protein aggregates, which can regulate cell functions related to, for example, growth and proliferation, inflammation, apoptosis, angiogenesis, metastasis, and immune responses, in various ways by affecting signal transduction pathways (Zhu et al., 1997; Yang et al., 2009).
In the present study, the relative immune organ weights were not significantly affected by dietary treatments. This result agrees with the results of Brenes et al. (2008) and Rahimi et al. (2011), who reported that broilers fed diet containing GP concentrate and thyme showed no significant differences in the relative weight of the spleen and bursa in compared with control groups.
The main indicators of the response of the intestinal tract to active substances in feeds are mucosa status and their microscopic structure. In the present study, villus height and crypt depth were reduced, particularly in jejunum. Since phenolic compounds are known as xenobiotic to the body, this response can be made to reduce the absorption of these compounds. Whereas, Caspary (1992)assumed that an increase in villus height could increase the digestive and absorptive function of the intestine by increasing the absorptive surface area, expression of brush border enzymes, and nutrient transport systems. In agreement with our results, Viveros et al. (2011) reported that inclusion of GPC and GSE decrease villus height and crypt depth in broiler chickens. Intestinal cell proliferation occurs mainly in the crypts (Iji et al., 2001). Thus, the large crypt suggests a high nutrient requirement for intestinal maintenance and reduced efficiency of the bird. It is anticipated that lower crypt depth as a result of fed GP can be considered as a beneficial way to decrease the cost of intestinal maintenance in birds.
In this study, we also found a decrease in villus height/crypt depth ratio in broilers fed diets. Length villus and a short crypt can improve nutrient absorption, decrease secretion in the gastrointestinal tract, increase disease resistance, and greater overall performance. In this sense, the low villi height/crypt depth ratio could have caused the poorer growth in GP-fed chicks. In contrast, Viveros et al. (2011) reported that an increase in villus height/crypt depth ratio at the jejunum in birds fed GPC diets. Moreover, muscularis thickness was decreased in birds fed GP diet. It is assumed that inclusion of feed antibiotics in chicken diets have ability to decrease thickness of walls and reduce weight of intestine (Miles et al., 2006). The antimicrobial properties of polyphenols have been demonstrated by many in vitro studies (Arunachalam et al., 2003). Thus, the lower crypt depth and epithelial thickness as a result of feeding polyphenols can lead to improve feed utilization and performance.
Intracellular enzymes release into the extracellular space when the liver is injured (Kristensen, 1994). The activity of AST is currently considered to be a very sensitive indicator of hepatocellular disease in avian species. The greater plasma AST concentration in the 10% GP treatment relative to the other treatment may suggest hepatocellular necrosis by polyphenol compounds, which causes an increase in permeability of the cell membrane, resulting in the release of transaminase in the blood stream. Many plant extracts such as grape polyphenols have been shown to play a double-edged role in the regulation of the inflammatory system of the host. Therefore, caution should be carefully taken in intake dosage of polyphenols to avoid switching over from beneficial effects to adverse ones. Regarding histopathological examination, Iqbal et al. (2015) reported that no changes were observed in the livers and kidneys of birds fed low and medium levels of GP polyphenols in diets. Also, mild changes in the livers and kidneys of birds fed high level of GP polyphenols diet suggested that even higher polyphenols levels are relatively nontoxic to birds.
It is worthy to mention that histological (Structural) changes observation in liver, supported the observed biochemical (Functional) changes in blood. In the present study, the AST activity was increased significantly in 10% GP fed group compared to control groups (P <0.05). There was no significant effect of feeding diets on liver weights in broilers. These results are in agreement with the reports of Brenes et al. (2008) who reported that the inclusion of graded concentrations of GPC did not affect the liver weight.
Conclusion
In conclusion, the results from this study suggest that supplemental GP up to 10 percent did not impair performance of broiler chickens. The addition of GP in the broiler diets could increase the immune responses and reduce the feed cost per kg of live weight. The histomorphometrical measurement of small intestine showed decrease in crypt depth and epithelial thickness of chicken duodenum and the villus height and crypt depth was significantly reduced in the jejunum. The physiological effects of polyphenols depend on many different factors and more research will be needed in the future to establish appropriate dosages of grape polyphenols to optimize health benefits and minimize possible negative effects.
Acknowledgements
The authors are grateful to the University of Mohaghegh Ardabil for founding support of the project.
Authors' Contributions
SKE and BN were in charge of the experimental design, implementation and writing the manuscript. PF and FMA conducted the feeding trial and data collection. All authors participated in interpreting and reviewing the results of the study.
Conflict of Interest Declaration
There are no conflicts of interest.
References
Abarghuei, M.J., Rouzbehan, Y. & Alipour, D., 2010. The influence of the grape pomace on the ruminal parameters of sheep. Livest. Sci. 132, 73-79. [ Links ]
Acamovic, T. & Brooker, J.D., 2005. Biochemistry of plant secondary metabolites and their effects in animals. Proc. Nutr. Soc. 64, 403-412. [ Links ]
Alonso, A.M., Guillé, D.A., Barroso, C.G., Puertas, B. & García, A., 2002. Determination of antioxidant activity of wine-byproducts and its correlation withpolyphenolic content. J. Agric. Food Chem. 50, 5832-5836. [ Links ]
Andriantsitohaina, R., Auger, C., Chataigneau, T., Etienne-Selloum, N., Li, H., Martınez, M.C., Schini-Kerth, V. B. & Laher, I., 2012. Molecular mechanisms of the cardiovascular protective effects of polyphenols. Briti. J.Nutri. 108, 1532-1549. [ Links ]
AOAC, 2000. Official Methods of Analysis.17th Ed. Association of Official Analytical Chemists, Arlington, VA. [ Links ]
Arunachalam, M., Raj, M.M., Mohan, N. & Mahadevan, A., 2003. Biodegradation of catechin. Proc. Indian Natl. Sci. Acad 69, 353-370. [ Links ]
Aviagen, 2014. Ross 308 Broiler: Nutrition Specification. Ross Breeders Limited,Newbridge, Midlothian, Scotland, UK. [ Links ]
Brenes, A., Viveros, A., Goni, I., Centeno, C., Sayago-Ayerdi, S.G., Arija, I. & Saura-Calixto, F., 2008. Effect of grape pomace concentrate and vitamin E on digestibility of polyphenols and antioxidant activity in chickens. Poul. Sci. 87, 307-316. [ Links ]
Brenes, A., Viveros, A., Saura, C. & Arija, I., 2016. Use of Polyphenol-rich grape by-products in monogastric nutrition. Anim. Feed Sci.Technol. 1,1-17. [ Links ]
Chamorro, S., Viveros, A., Centeno, C., Romero, C., Arija, I. & Brenes, A., 2013. Effects of dietary grape seed extract on growth performance,amino acid digestibility and plasma lipids and mineral content in broiler chicks. Anim. 7, 555-561. [ Links ]
Caspary, W.F., 1992. Physiology and pathophysiology of intestinalabsorption. Amer. J. Clini.Nutri. 55, 299-308. [ Links ]
Corrier, D.E. & DeLoach, J.R., 1990. Evaluation of cell-mediated,cutaneous basophil hypersensitivity in young chickens by an interdigital skin test. Poul. Sci. 69, 403-408. [ Links ]
Dorri, S., Tabeidian, A.S., Toghyani, M., Jaha-nian, R. & Behnamnejad, F., 2012. Effect of different levels of grape pomace on blood serum and biochemical parameters of broiler chicks at 29 and 49 days of age. Proc. 11th Int. and 4th Natl. Congress on Recycling of Organic Waste in Agriculture. Isfahan, Iran. [ Links ]
Ebrahimzadeh, S.K., Navidshad, B., Farhoomand, P. & MirzaeiAghjehgheshlagh, F., 2017. Effects of exogenous tannase enzyme on growth performance, antioxidant status, immune response, gut morphology and intestinal microflora of chicks fed grape pomace. S. Afr. J. Anim. Sci. 48. [ Links ]
FAO, 2013. STAT-FAO Stastical Data-base, http://faostat3.fao.org [ Links ]
Foti, M., Piatelli, M., Amico, V. & Ruberto, G.,1994. Antioxidant activity of phenolic meroditerpenoids from marine algae. J. Photochem. Photobiol. 26, 159-164. [ Links ]
Georgiev, V., Ananga, A. & Tsolova, V., 2014. Recent advances and uses of grape flavonoids as nutraceuticals. Nutri. 6, 391-415. [ Links ]
Goni, I., Brenes, A., Centeno, C., Viveros, A., Saura-Calixto, A., Rebole, I. & Arija, R., 2007. Effect of Dietary Grape Pomace and Vitamin E on Growth Performance,Nutrient Digestibility, and Susceptibility to Meat Lipid Oxidation in Chickens. Poult. Sci. 86, 508-516. [ Links ]
Griffiths, D. W., 1986. The inhibition of digestive enzymes by polyphenolic compounds. Advan. Experimen. Med. Biol. 199, 509-516. [ Links ]
Hajati, H., Hassanabadi, A., Golian, A., Nassiri-Moghaddam, H. & Nassiri, M.R., 2015. The effect of grape seed extract and vitamin C feed supplementation on some blood parameters and HSP70 gene expression of broiler chickens suffering from chronic heat stress. Itali. J. Ani. Sci. 14, 3273. [ Links ]
Hong, J.C., Steiner, T., Aufy, A. & Lien, T.F., 2012. Effects of supplemental essential oil on growth performance, lipid metabolites and immunity, intestinal characteristics, microbiota and carcass traits in broilers. Livest. Sci. 144,253-262. [ Links ]
Horigome, T., Kumar, R. & Okamoto, K.,1988. Effects of condensed tannins prepared from leaves of fodder plants on digestive enzymes in vitro and in the intestine of rats. Briti. J. Nutrit. 60, 275-278. [ Links ]
Hughes, R.J., Brooker, J.D. & Smyl, C., 2005. Growth rate of broiler chickens given condensed tannins extracted from grape seed. Pages 56-68 in Aust. Poult. Sci. Symp. Poultry Research Foundation, University of Sidney, Sidney, Australia. [ Links ]
Iji, P.A., Saki, A. & Tivey, D.R., 2001. Body and intestinal growth of broiler chicks on a commercial starter diet. 1. Intestinal weight and mucosal development. Briti. Poult. Sci. 42,505-513. [ Links ]
Iqbal, Z., Kamran, Z., Sultan, J.I., Ali, A., Ahmad, S., Shahzad, M.I., Ahsan, U. & Ashraf Sand Sohail, M.U., 2015. Replacement effect of vitamin E with grape polyphenols on antioxidant status, immune, and organs histopathological responses in broilers from 1- to 35-d age. J. Appl. Poul. Res. 24, 127-134. [ Links ]
Jansman, A.J.M., Huisman, J. & van der Poel, A.F.B., 1989. Recent Advances in Research of Antinutritional Factors in Legume Seeds., ed Pudoc, Wageningen, the Netherlands. [ Links ]
Kristensen, S.R., 1994. Mechanisms of cell damage and enzyme release. Danmark Medica Bullteen. 41,423-433. [ Links ]
Lau, D.W. & King, A.J., 2003. Pre-and post-mortem use of grape seed extract in dark poultry meat to inhibit development of thiobarbituric acid reactive substances. J. Agri. Food Chem. 51,1602-1607. [ Links ]
Longstaff, M. & McNab, J. M.,1991. The inhibitory effects of hull polysaccharides and tannins of field beans (Vicia faba L) on the digestion of amino acids, starch and lipid and on digestive enzyme activities in young chicks. Brit. J. Nutri. 65, 199-216. [ Links ]
Leshchinsky, T.V. & Klasing, K.C., 2001. Relationship between the level of dietary vitamin E and the immune response of broiler chickens. Poult. Sci. 80, 1590-1599. [ Links ]
Llobera, A. & Canellas, J.,2007. Dietary fiber content and antiox-idant activity of Manto Negro red grape (Vitis vinifera): Pomace and stem. Food Chem. 101,659-666 [ Links ]
Lien, T.F., Yeh, H.S. & Su, W.T., 2008. Effect of adding extracted hesperetin, naringenin and pectin on egg cholesterol, serum traits and antioxidant activity in laying hens. Arch. Anim. Nutri. 62, 33-43. [ Links ]
Martel, F., Monteiro, R. & Calhau, C., 2010. Effect of polyphenols on the intestinal and placental transport of some bioactive compounds. Nutri. Res. Revi. 23, 47-64. [ Links ]
McDougall, G.J., Shpiro, F., Dobson, P.,Smith, P., Blake, A. & Stewart, D., 2005. Different polyphenolic components of soft fruits inhibit alpha-amylase and alpha-glucosidase. J. Agri. Food Chem. 53, 2760-2766. [ Links ]
Miles, R.D., Butcher, G.D., Henry, P.R. & Littell, R.C., 2006. Effect of antibiotic growth promoters on broiler performance, intestinal growth parameters and qualitative morphology. Poult. Sci. 85, 476-485. [ Links ]
Ortiz, L.T., Centeno, C., Trevino, J.,1993. Tannin in faba bean seeds: effects on the digestion of protein and amino acids in growing chicks. Anim. Feed Sci. Technol. 41, 271-278. [ Links ]
Ozgan, A., 2008. Use of grape seed oil in functional egg production [MSc Thesis]. Adana (Turkey): University of Cukurova. [ Links ]
Placer, Z.A., Cushman, L.L. & Johnson, B.C., 1966. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 16, 359-364. [ Links ]
Prochazkova, D., Bousova, I. & Wilhelmova, N., 2011. Antioxidant and prooxidant properties of flavonoids. Fitoterapia. 82, 513-523. [ Links ]
Qureshi, M.A. & Havanstein, G.B., 1994. A comparison of the immune performance of a 1991 commercial broiler with a 1957 randombred strain when fed "typical" 1957 and 1991 broiler diets. Poult. Sci. 73,1805-1812. [ Links ]
Rahimi, S., Teymouri Zadeh, Z., Karimi Torshizi, M.A., Omidbaigi, R. & Rokni, H., 2011. Effect of the three herbal extracts on growth performance, immune system, blood factors and intestinal selected bacterial population in broiler chickens. J. Agri. Sci. Technol. 13, 527-539. [ Links ]
SAS, 2001. Statistical Analysis Systems/SAS, STAT User's guide Statistics. Version 9.1. [ Links ]
Sahin, K., Orhan, C., Tuzcu, M., Ali, S., Sahin, N.& Hayirli, A., 2010. Epigallocatechin-3-gallate prevents lipid peroxidation and enhances antioxidant defense system via modulating hepatic nuclear transcription factors in heat-stressed quails. Poult. Sci. 89, 2251-2258. [ Links ]
Sannamani, P.G., 2002. Feeding value of sorghum grains vis-a-vis yellow maize for broiler chicks. M. V. Sc. Thesis, submitted to Deemed University, IVRI, Izatnagar, Indi. [ Links ]
Seven, T.P., Seven, I., Yilmaz, M.M. & Ugsims, E.K., 2008. The effects of turkish propolis on growth and carcass charac-teristics in broilers under heat stress. Anim. Feed Sci. Technol. 146, 137-148. [ Links ]
Shi, J., Yu, J., Pohorly, J.E. & Kakuda, Y., 2003. Polyphenolics in grape seeds-biochemistry and functionality. J. Medic. Food. 6, 291-299. [ Links ]
Singleton, V.L. & Rossi, J.A., 1965. Colorimetry of total phenolics with phosphomolybdic phosphotungstic acid reagents. Amer. J. Enol. Viticul. 16,144-158. [ Links ]
Surai, P. F. & Sparks, N. H. C., 2001. Designer eggs: from improvement of egg composition to functional food. Trends in Food Sci. Technol. 12, 7-16. [ Links ]
Surai, P.F., 2002. Natural Antioxidants in Avian Nutrition and Reproduction. Nottingham University Press, Nottingham, UK. [ Links ]
Surai, P. F., 2006. Selenium in Nutrition and Health. Nottingham University Press, Nottingham, UK. [ Links ]
Surai, P.F. & Fisinin, V. I., 2010. Ill health effects of food lipids: consequences of inadequate food processing, storage and cooking. In: F. De Meester, S. Zibadi, R.Ross (eds), Modern Dietary Fat Intakes in Disease Promotion. Humana Press. [ Links ]
Surai, P.F., 2014. Polyphenol compounds in the chicken/animal diet: from the past to the future. J. Anim. Physiol. Anim. Nutr. 98,19-31. [ Links ]
Tepe, B., Daferera, D., Sokmen, M., Polission, M. & Sokmen, A., 2004. In vitro antimicrobial and antioxidants activities of essential oils and various extracts of Thymus eigii. J. Agri. Food Chem. 52, 1132-1137. [ Links ]
Torres, J.L., Varela, B., García, M.T., Carilla, J., Matito, C., Centelles, J.J., Cascante, M., Sort, X. & Bobet, R., 2002. Valorization of grape (Vitis vinifera) byproducts.Antioxidant and biological properties of polyphenolic fractions differing in procyanidin composition and flavonol content. J. Agric. Food Chem. 50, 7548-7555. [ Links ]
Viveros, A., Chamorro, S., Pizarro, M., Arija, I., Centeno, C. & Brenes, A., 2011. Effects of dietary polyphenol-rich grape products on intestinal microflora and gut morphology in broiler chicks. Poult. Sci. 90, 566-578. [ Links ]
Xia, E.Q., Deng, G.F., Ya-Jun,K., Guo, Y.J. & Hua-Bin Li, H.B., 2010. Biological activities of polyphenols from grapes. Inter. J. Mol. Sci. 11, 622-646. [ Links ]
Yang, C.S., Wang, X., Lu, G. & Picinich, S.C., 2009. Cancer antioxidant redox signalling. Life Sci. 7, 1704-1714. [ Links ]
Yilmaz, Y. & Toledo, R.T., 2004. Major flavonoids in grape seeds and skins: antioxidant capacity of catechin, epicatechin, and gallic acid. J. Agri. Food Chem. 52, 255-260. [ Links ]
Yilmazer-Musa, M., Griffith, A.M., Michels, A.J., Schneider, E. & Frei, B., 2012. Grape seed and tea extracts and Catechin 3-gallates are potent inhibitors of a-amylase and a-glucosidase activity. J. Agri. Food Chem. 60, 8924-8929. [ Links ]
You,Q., Chen, F., Wang, X., Luo, P.G. & Jiang, Y., 2011. Inhibitory effects of muscadine anthocyanins on a-glucosidase and pancreatic lipase activities. J. Agri. Food Chem. 59, 9506-9511. [ Links ]
Zhu, M., Phillipson, D., Greengrass, P.M., Bowery, N.E. & Cai, Y., 1997. Plant polyphenols: Biologically active compounds or non-selective binders to proteins Phytomed. 3, 281-285. [ Links ]
Received 16 October 2017
Accepted 10 December 2017
First published online 26 December 2017
# Corresponding author: bnavidshad@uma.ac.ir


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}