










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.47 no.6 Pretoria 2017
http://dx.doi.org/10.4314/sajas.v47i6.18
Effects of dietary Aloe vera crude extracts on digestive enzyme activities and muscle proximate composition of GIFT tilapia juveniles
N.N. GabrielI, #; J. QiangII, III; X.Y. MaII; P. XuII, III; D.N. NakwayaI
IDepartment of Fisheries and Aquatic Sciences, Sam Nujoma Campus, University of Namibia Henties Bay, Namibia
IIWuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081, China
IIIChina Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization, Ministry of Agriculture, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081, Jiangsu, China
ABSTRACT
This study investigated the effects of dietary Aloe vera powder extract on the activities of digestive enzymes and muscle composition of Oreochromis niloticus (GIFT strain) juveniles. Five dietary groups were formulated in which A. vera powder was added into a tilapia feed at 0.5%, 1.0%, 2.0%, and 4%/kg feed, and fed for eight weeks. Amylase, trypsin, and lipase activities in the liver and along the alimentary canal (stomach, midgut, and hindgut) varied significantly between dietary groups. Amylase activity was significantly enhanced in the liver of fish fed 0.5, 1, and 2% dietary A. vera, and in the stomach of those fed 1%. Total amylase activity in the gastrointestinal tract (total gut) was significantly enhanced in fish fed 0.5% and 1% compared with control and those fed 4% dietary A. vera. Trypsin activity was significantly enhanced in the stomach of fish supplemented with 0.5% dietary A. vera and in the duodenum of those supplemented with 1%, 2%, and 4% dietary A. vera when compared with the control. Lipase activity was increased in the duodenum of fish fed 1.0% dietary A. vera when compared with the control. Muscle moisture content was significantly lower in all A. vera-supplemented fish, whereas protein was lower in those fed 2% and 4% A. vera diet when compared with the control. Based on second-order polynomial regression analysis, dietary A. vera inclusion level less than or equal to 1.76%, 1.82%, 2.10%/kg feed was determined to be suitable in enhancing carbohydrate, protein and lipid digestion, respectively, in GIFT tilapia in this study.
Keywords: Aquaculture, digestion enhancers, freshwater fish, medicinal herbs
Introduction
Over the years, fish farmers have used pharmaceutical drugs to prevent stress-related conditions, which affect production, especially in intensive fish farming systems. The success of commercial antibiotics in aquaculture and livestock and poultry farming lies in their ability to promote growth, enhance feed conversion efficiency and prevent diseases (Shalaby et al., 2006). In China, Thailand and Vietnam, both semi-intensive and intensive shrimp production have been reported to rely heavily on chemical inputs per unit ton of harvested produce (Rico et al., 2013). However, the continuous use of antibiotics and other chemicals has numerous shortcomings, such as the risk of the pathogens developing resistance, the problem of drug residues in treated animals, the effects on human health, and environmental pollution (Wu et al., 2013). Thus, many nations have strict regulations that limit the use of antibiotics in animal farming (Dibner & Richards, 2005). At present, there is an urgent need to explore alternatives to pharmaceutical drugs that could be used to improve growth performance and disease control in intensive fish farming systems sustainably.
Several herbal extracts have been reported to improve nutrient utilization, growth performance, survival, and body composition of fish in aquaculture. For instance, dietary Astragalus membranaceus reportedly enhanced amylase and lipase activities, and subsequently improved feed conversion ratio, feed intake, weight gain and specific growth rate of Oreochromis niloticus juveniles (Zahran et al., 2014). In part, similar findings were observed in Labeo rohita juveniles when fed a diet supplemented with Mucuna pruriens extracts (Ojha et al., 2014), and Achyranthes aspera extracts (Vasudeva et al., 2006), respectively. In addition, improved survival rate, and disease resistance was reported in Ictalurus punctatus juveniles fed dietary Origanum vulgare extracts (Zheng et al., 2009), and in Cyprinus carpio fed dietary Rheum officinal extracts (Xie et al., 2008). This is an indication that medicinal herbal extracts could be used as alternatives to pharmaceutical drugs in fish farming.
Aloe vera (synonym Aloe barbadensis), a perennial succulent stemless tropical and subtropical herb has been scientifically proven to be rich in biological active compounds (Langmead et al., 2004). Throughout history, this herb has been used in humans to cure disorders associated with the digestive system such as poor digestion, constipation, anorexia, abdominal distension, colitis, and conditions such as diabetes, asthma, and uterine pain (Sahu et al., 2013). It has been also used to treat abrasions, burns and skin irritations, acne, and dermatitis (Puvabanditsin & Vongtongsri, 2005; Sahu et al., 2013). Several studies have reported antibacterial anti-inflammatory immune-modulatory effects (Gautam et al., 2004; Madan et al., 2008; Moorthy et al., 2009), and antioxidation properties (El-Shemy et al., 2010) of Aloe vera. Recently, its potential to serve as an alternative growth promoter, appetizer and digestive stimulant has been reported in C. carpio (Alishahi et al., 2010), Oncorhynchus mykiss (Heidarieh et al., 2013; Golestan et al., 2015), and terrestrial animals such as poultry (Mehala & Moorthy, 2008; Bolu et al., 2013). To date, there have been limited reports on the effects of A. vera extracts on digestive enzyme activities, and proximate muscle composition in farmed Nile tilapia. Therefore, this study was conducted to test the hypothesis that dietary supplementation of standard tilapia diet with A. vera powder would enhance the digestive enzyme activities and fillet composition of GIFT tilapia. This experiment was crucial in that it would expand knowledge of the way in which medicinal herbal extracts affect feed utilization and subsequently growth performance in farmed fish.
Material and methods
Five iso-nitrogenous (31.7% crude protein), iso-energetic (672 kJ/100 g) and iso-lipid (7.34%) diets were formulated to contain 0%, 0.5%, 1%, 2%, and 4% (kg-1 feed) of A. vera powder crude extract. A. vera crude extract was a commercial product (Jiangsu Zhe Ya Food Co. Ltd) as demonstrated in the authors' previous studies (Gabriel et al. 2015a; 2015b) (Table 1). Briefly, all feedstuffs (fish meal, corn starch, soybean oil, soybean meal, cotton seed meal, rapeseed meal, choline chloride, vitamin C phosphate ester, Calcium dihydrogen phosphate, and cellulose) for each diet were powdered, and mixed mechanically in a food mixer for 40 min. Water was then added gradually until a paste was obtained. The paste for each diet was then pelleted into 16-mm diameter granular feed using a laboratory feed machine and then dried at ambient temperature. Each diet was sieved through a micron sieve to produce 0.2-0.8 mm pellets, which were packed in plastic-lined bags, and stored at 4 °C until use.
A total of 375 healthy GIFT tilapia juveniles (average bodyweight 4.83 ± 0.01 g) were obtained from the tilapia breeding centre of Freshwater Fisheries Research Center (FFRC) in Wuxi. They were transported in polythene bags filled with oxygen. The fish were stocked in cylindrical blue plastic tanks (0.6 m2 x 0.85 m), supplied with 300 L de-chlorinated freshwater at 28.6 ± 0.12 °C, pH 7.93 ± 0.24, dissolved oxygen (DO) 6.62 ± 0.16 mg/L (YSI 650 MDS multiprobe system, YSI Inc. USA) under a natural photoperiod, continuous aeration and water-recirculating system. To maintain water quality, two thirds of the cultured water was exchanged with dechlorinated freshwater of similar temperature three times a week. The fish were habituated for seven days. During this period, fish were fed thrice daily (09:00; 13:00; 17:00) with a commercial diet (No. 5271, 35% crude protein, Ningbo Tech-Bank Co. Ltd, Yuyao City, China) until apparent satiation. After the habituation period, fish were randomly distributed into 15 tanks in five triplicate dietary groups at a stocking density of 25 fish/tank. The fish were hand-fed the experimental diets for 60 days, 6 days a week, 3 times a day (09:00; 13:00; 17:00).
After 24 hours of the last experimental feeding, three fish from each replicate were sedated (200 mg MS-222/L of water), and liver and gut (stomach, midgut and hindgut) were gently collected and pooled to determine amylase, trypsin, and lipase activities, respectively. Single tissues were weighed and homogenized in ice-cold phosphate buffer saline (PBS, pH 7.4) at 1:9 ratio using a bead homogenizer (Scientz-48, Ningbo Scientz Biotechnology Co. Ltd, China). The homogenates were centrifuged at 14489.28g, 4 °C for 15 min. The supernatants were stored at -20 °C as crude enzyme extracts without further purification.
Digestive enzymes activities of each sample were analysed in triplicate using commercial fish enzyme-linked immunosorbent assay (ELISA) kits (Shanghai Lengton Biological Technology Co. Ltd., China) followed by spectrophotometry (Bioteck Instruments, Inc, USA). Each ELISA kit included a standard solution (0.5 ml), its concentration and corresponding optical density (OD) at 450 nm for 10 min were measured using a spectrophotometer. This standard concentration and OD were used to establish a standard linear regression equation for each enzyme test kit. The standard linear equation for amylase (PAMY, catalogue No. BPE90037), trypsin (catalogue No. BPE94113) and lipase (PL, Catalogue No. BPE94273) were y = 1320.9X + 32.38, y = 152.7X + 9.68, and y = 182.35X+ 5.50, respectively. To determine y or the activity of each enzyme in each tissue, X was substituted with OD 450 nmm, 10 min values of the sample.
Dorsal muscles (fillets) from three fish in each replicate were collected and stored at -20 °C for proximate composition analyses conducted by the standard method of AOAC (1990). Moisture content was determined by oven drying at 105 °C until constant weight and expressed as a percentage (% moisture = wet weight weight/ sample weight x 100). Crude protein (nitrogen x 6.25) was determined by the Kjeldahl method using the Auto Kjeldahl system (FOSS KT260, Switzerland) and was expressed as a percentage. Crude lipid was determined by an ether extraction system (Ankom XT15, USA) and was expressed as a percentage (% lipid = weight of the residue/ weight of the sample taken x 100). Ash was determined by burning the dry samples at 560 °C for 5 hours and was expressed as a percentage (% ash = weight of dry sample/ weight of sample x 100).
Data were expressed as mean ± standard error (M±SE), and were analysed using one-way analysis of variance (ANOVA) in SPSS. Duncan's multiple range test with significance set at 95% was used to separate treatment means. The second-order polynomial regression model (Zeitoun et al., 1976) was used to estimate the optimum dietary A. vera requirement suitable for maximum digestive enzyme activities in GIFT tilapia juveniles.
Results
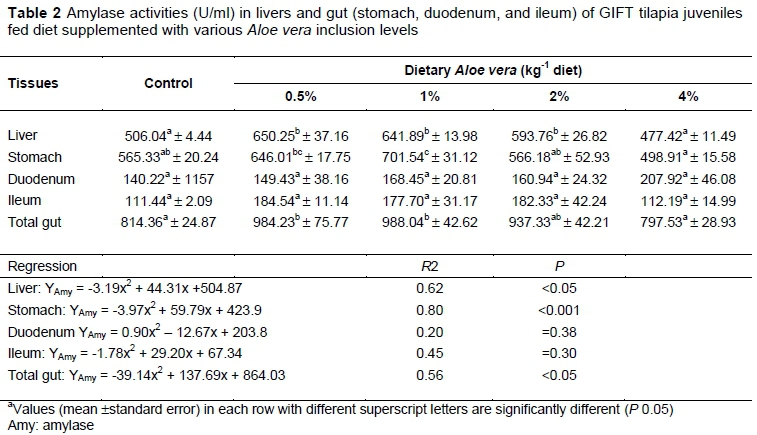
Amylase, trypsin, and lipase activities in liver,and gut sections (stomach, midgut, hindgut, and total gut) varied significantly (P < 0.05) between dietary groups. Amylase activity in livers of fish supplemented with 0.5%, 1%, and 2% A. vera was higher (P< 0.05) than those fed the control, and 4% diets (Table 2). Amylase activities in the stomach of fish fed the 1.0% diet were higher (P< 0.05) than those fed 2.0% and 4% diets. Total amylase activities in the gastrointestinal tract (total gut) were higher (P< 0.05) in fish fed 0.5% and 1% compared with those fed the control and 4% diets. Amylase activities responded quadratically to the dietary A. vera inclusion levels in the liver (YAmy = -3.19x2 + 44.31x +504.87, R2 =0.62, P = 0.003), stomach (YAmy = -3.97x2 + 59.79x + 423.9, R2= 0.80, P = 0.00), duodenum (YAmy = 0.90x2 - 12.67x + 203.8, R2 = 0.20, P = 0.38), and ileum (YAmy = -1.78x2 + 29.20x + 67.34, R2 =0.45, P= 0.30), respectively. Based on the second-order polynomial analysis (Figure 1), the optimum dietary A. vera inclusion level (%) required for maximum total amylase activity was determined as 1.76% (YAmy. = -39.135x2 + 137.69x + 864.03, P = 0.007).
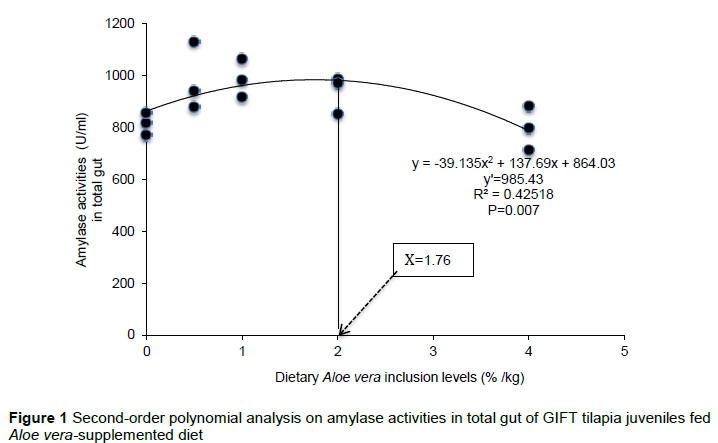
In addition, 0.5%, 1% and 2% dietary A. vera had no effect (P> 0.05) on trypsin activities in the liver. However, 4% dietary A. vera presented lower (P< 0.05) trypsin activities in the liver when compared with the control (Table 3). Trypsin activities were higher (P< 0.05) in the stomachs of fish supplemented with 0.5%, and 4% dietary A. vera and in duodenum of those supplemented with 1%, 2%, and 4% dietary A. vera when compared with the control. No significant difference (P> 0.05) was observed in the total trypsin activities of the gastrointestinal tract among groups. Trypsin activities responded quadratically to the dietary A. vera inclusion levels in the liver (YTryp = -0.12x2- 4.97x + 133.12, R2 0.50, P = 0.20), stomach (YTryp = 0.49x2 - 9.22x + 118.9, R2= 0.70, P = 0.01), duodenum (YTryp = -1.7x2 + 4.3620x + 12.93, R2 = 0.30, P= 0.24), and ileum (YTryp = -0.80x2 - 1.47x + 9.73, R2 =0. 16, P = 0.28), respectively. Based on the second-order polynomial analysis (Figure 2), the most suitable dietary A. vera inclusion level (%) for maximum total trypsin activities was estimated to be 1.82% (YTryp. = 3.1733x2 - 11.527x + 137.85, P = 0.051).
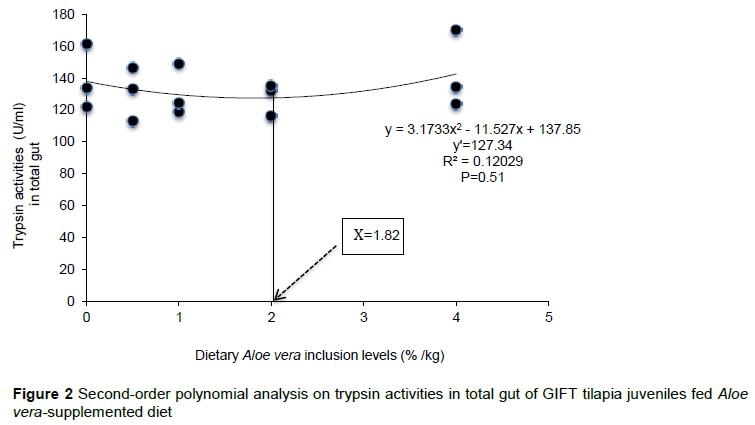
Lipase activities in liver, stomach, ileum, and total gut did not differ significantly (P> 0.05) among dietary groups, except in the duodenum of fish fed 1.0% dietary A. vera when compared with unsupplemented fish. However, high lipase activities were observed in A. vera-supplemented fish compared with unsupplemented ones (Table 4). Similar to amylase and trypsin, lipase activities responded quadratically to the dietary A. vera inclusion levels in liver (YLip = -0.25x2- 6.20x + 126.29, R2 = 0.30, P = 0.19), stomach (YLip = 0.11x2 - 3.64x + 160.75, R2 = 0.23, P = 0.26), duodenum (YLip = -0.43x2 - 8.38x + 1.47, R2 =0.43, P = 0.11), and ileum (YLip = -0.04x2 - 0.21x + 24.02, R2 =0.17, P= 0.26), respectively. Based on the second-order polynomial analysis (Figure 3), the optimum dietary A. vera inclusion level (%) for maximum total lipase concentration could be less than or equal to 2.10% (YLip = -1.6667x2 - 6.9933x + 196.71, P = 0.81).
A. vera-supplemented fish showed lower (P< 0.05) muscle moisture content when compared with control (Table 5). Similarly, muscle protein content in fish fed 2% and 4% A. vera supplemented diet was significantly lower (P< 0.05) compared with unsupplemented ones and those fed 0.5%, 1%, and 2% dietary A. vera. No significant difference (P> 0.05) was observed in crude lipid and ash among dietary groups.
Discussion
The authors' previous study (Gabriel et al., 2015) reported that dietary A. vera improved growth, feed utilization efficiency, feed intake, and health parameters in GIFT tilapia. Overall, an inclusion level less than 2% Aloe vera/kg feed seemed suitable. Similar results were partly reported in common carp (Cyprinus carpio) (Mahdavi et al., 2013), and rainbow trout (Oncorhynchus mykiss) (Heidarieh et al., 2013) after being fed with dietary A. vera, respectively. Improved growth and feed utilization parameters in these fish were assumed to be the result of A. vera's ability to increase nutrient digestibility, absorption and assimilation capacity, partly through improved digestive enzymes and healthy intestinal microflora. Hence, evaluation of digestive enzymes profiles in digestive organs such as liver and gastrointestinal tract may in part be useful in determining nutrient digestion capability and feed utilization efficiency of fish (Buddington et al., 1997), especially following dietary herbal administration.
Oreochromis niloticus alimentary tracts (stomach, duodenum, and ileum) possess a variety of key carbohydrate (amylase, sucrase, maltase and cellulase), protein (trypsin, chymotrypsin and pepsin) and fat (lipase) metabolizing enzymes (Oyedapo et al., 2005). The distribution and activities of these digestive enzymes differ along the gut and digestive organs (liver, pancreas and gallbladder) (Oyedapo et al., 2005). Their distribution and activities depend mostly on the nature and composition of the diet administered to the animals (Ismat et al., 2013). Previous researchers have indicated that medicinal herbal extracts stimulate digestion with enhanced bile acid concentration and stimulate the pancreas and increased secretion of digestive enzymes (lipases, amylases and proteases) in fish (Bhosale et al., 2010). This in part supports the results of the present study, in which amylase activity was significantly higher in liver, stomach section, and total gut, especially in fish fed 0.5% and 1% A. vera diet. Furthermore, variations in the effects of dietary A. vera on digestive enzymes activities throughout the digestive tract of GIFT-tilapia juveniles were observed in this study. This is partly in accordance with the study by Zahran et al. (2014), which demonstrated that Astragalus extracts (polysaccharides) significantly increased amylase activity, and had no significant influence on lipase activity in Nile tilapia. This lack of significant influence of dietary Astragalus on lipase activity in Nile tilapia seemed to be a result of a low intake of lipid from plant-related diets (Tengjaroenkul et al., 2000). El-Dakar (2015) reported that basil (Ocimum basilicum) extracts significantly increased serum lipase activity in Sparus aurata fingerlings, while protease activity was reported to increase significantly in Labeo rohita fingerlings after being fed with Mucuna pruriens ethanolic extracts (Ojha et al., 2014).
Furthermore, based on the second-order polynomial analyses, the optimum dietary A. vera was estimated to be 1.76%, 1.82%, and 2.10% for amylase, trypsin, and lipase concentration, respectively. Dietary A. vera optimum levels suitable for the enhancement of digestive enzymes in this study corresponded closely with dietary A. vera inclusion level, which supported maximum growth performance and feed utilization in the same fish species reported in the authors' previous study (Gabriel et al., 2015). This supports the assumption that medicinal herbal extracts have the ability to improve growth performance in fish by enhancing their nutrient digestibility, absorption and assimilation capacity through improved digestive enzyme distribution, and activities, as narrated earlier in this study. Moreover, improved digestive enzyme activities in the present study, feed utilization, and growth reported in the authors' previous studies (Gabriel et al., 2015) in A. vera-supplemented fish could partly be attributed to a wide range of polysaccharides such as acemannan, galactan, cellulose, pectin, glucogalactomannan, and glucoronic acid that are present in A. vera leaves (Hamman, 2008). Polysaccharides are believed to possess prebiotic properties, which benefit the host by stimulating the growth or activity of one or a limited number of bacteria species already resident in the gut, thus improving the host's gut microflora and ultimately growth through improved feed utilization and digestive enzyme activities (Gabriel et al., 2015).
Similar to growth performance and feed utilization, body composition of fish can be altered by the nutrient concentration, quality of the diet, rations, feeding frequency and other factors (Jobling, 2001). Medicinal herbal extracts including those from guava, camphor trees (Abdelhamid & Soliman, 2012), basil (Ocimum basilicum), Cinnamomum zelanicum, Juglans regia, Mentha piperita (Al-Basali & Mohamad, 2010), and caraway (Ahmad & Abdel-Tawwab, 2011) have been reported to improve fish body composition, respectively. In the present study, dietary Aloe significantly affected some muscle proximate composition parameters of GIFT tilapia juveniles. Fish fed dietary A. vera showed a significant decrease in muscle moisture content compared with the unsupplemented ones. The high muscle moisture content observed in A. vera unsupplemented fish may be a disadvantage in that it increases the fish's susceptibility to microbial spoilage, and oxidative degradation of polyunsaturated fatty acid, and consequently decreases the fish meat quality and shelf life, as reported by Omolara & Omotayo (2008). Despite variation in muscle moisture content between A. vera-supplemented fish and unsupplemented ones, the moisture content of fish in all test groups was within the acceptable range, which is 60%-80% (Olagunju et al., 2012). Furthermore, muscle protein content of the fish supplemented with 2% and 4% dietary A. vera was observed to be significantly lower among groups. This is in accordance with the authors' previous study (Gabriel et al., 2015), which reported that 2% and 4% dietary A. vera resulted in anaemic and stressed fish. Herbal extracts are believed to be toxic, especially at high dosages, leading to unhealthy animals with poor defence mechanisms against stressors (Gabriel et al., 2015). This may be the case in muscle protein content of fish fed 2% dietary A. vera dosage or more. Therefore, a dietary A. vera inclusion level greater than 2% may lead to fish with poor nutritional quality in addition to poor health parameters (Gabriel et al., 2015).
Conclusion
The present study showed that Aloe vera powder extracts have the potential to significantly enhance digestive enzyme activities, and improve muscle proximate composition of GIFT tilapia juveniles. However, before Aloe vera powder extracts could be recommended as a tilapia feed supplement to improve feed digestion, and meat quality, more research, including extract purification and development of a perfect treatment regime, is deemed necessary.
Acknowledgments
The authors are grateful for the financial support provided by the following grants: Special Fund for Agro-scientific Research in the Public Interest (2015JBFR04) and the National Science & Technology Program (2012BAD26B03-1), which made this work possible. They also would like to thank Wuxi fisheries college of NAU, Department of Biotechnology, China, for providing facilities for the research work, and anonymous reviewers for their constructive criticism for the improvement of this piece of writing.
Authors' contributions
N.N.G was the principal investigator of this project, and the writer of the manuscript. J.Q, X.Y.M, and N.N.G designed the project, and were involved in specimen collection (sampling), and lab works. P.X was the overall supervisor of the project. Meanwhile, D.N.N assisted in editing the manuscript.
Conflict of interest declaration
The authors declare that they have no conflict of interest.
References
Abdelhamid, A.M. & Soliman, A.A.A., 2012. Possibility of using dried leaves of guava and camphor trees in tilapia diets. J. Arabian Aquacult. Soc. 7 (1). [ Links ]
Ahmad, H.M. & Abdel-Tawwab, M., 2011. The use of caraway seed meal as a feed additive in fish diets: growth performance, feed utilization, and whole-body composition of Nile tilapia, Oreochromis niloticus (L.) fingerling. Aquaculture. 314, 110-114. [ Links ]
Al Basali, H. & Mohamad, S., 2010. Immune response of common carp (Cyprinus carpio) fed with herbal immunostimulants diets. J Anim Vet Adv 9: 1839-1847. [ Links ]
Alishahi, M., Ranjbar, M.M., Ghorbanpour, M., Peyghan, R., Mesbah, M. & Razi, J.M., 2010. Effects of dietary Aloe vera on some specific and non-specific immunity in the common carp (Cyprinus carpio). Int. J. Vet. Res. 4, 189-195. [ Links ]
AOAC., 1990. Official methods of analysis (15th ed.), Association of Official Analytical Chemists. Washington, DC. [ Links ]
Bhosale, S.V., Bhilave, M.P. & Nadaf, S.B., 2010. Formulation of fish feed using ingredients from plant sources. RJAS. 1, 284-287. [ Links ]
Bolu, S.A., Babalola, T.O., Elelu, N., Ahmed, R.N., Oyetunde, S.A., Ademola, P.F. & Jiddah, S.A., 2013. Effects of supplemental Aloe vera gel in drinking water on some performance histology, hematology, serum constituents and growth of turkey poults challenged with Escherichia coli [J]. Wudpecker J. Agric. Res. 2, 223-229. [ Links ]
Buddington, R.K., Krogdahl, A. & Bakke-Mckellep, A.M., 1997. The intestine of carnivorous fish: Structure and functions and the relations with diet. Acta Physiol. Scandin. 638, 67-80. [ Links ]
Dibner, J.J. & Richards, J.D., 2005. Antibiotic growth promoters in agriculture: History and mode of action. J. Poultry Sci. 84, 634-643. [ Links ]
El-Dakar, Y.A., Shalaby, M.S., Nemetaliah, B.R., Saleh, E.N., Sakr, E.M. & Toutou, M.M., 2015. Possibility of using basil (Ocimum basilicum) supplementation in gilthead sea bream (Sparu aurata) diet Egypt J. Aquactic. Res. http://dx.doi.org/10.1016/j.ejar.2015.03.006. [ Links ]
El-Shemy, H.A., Aboul-Soud, M.A., Nassr-Allah, A.A., Aboul-Enein, K.M., Kabash, A. & Yagi, A., 2010. Antitumor properties and modulation of antioxidant enzyme's activity by Aloe vera leaf active principles isolated via supercritical carbon dioxide extraction. Curr. Med. Chem. 17, 129-138. [ Links ]
Gabriel, N.N., Qiang, J., He, J., Ma, Y.X., Kpundeh, M.D. & Xu, P., 2015. Dietary Aloe vera supplementation on growth performance, some haemato-biochemical parameters and disease resistance against Streptococcus iniae in tilapia (GIFT).Fish Shellfish Immunol.44, 504-514. [ Links ]
Gautam, M., Diwanay, S.S., Gairolac, S., Shinde, Y.S., Jadhav, S.S. & Patwardhan, B.K., 2004. Immune response modulation to DPT vaccine by aqueous extract of Withania somnifera in experimental system. J. Ethnopharma. 4, 841-849. [ Links ]
Golestan, G., Salati, A.P., Keyvanshokooh, S., Zakeri, M. & Moradian, H., 2015. Effect of dietary Aloe vera on growth on growth and lipid peroxidation indices in rainbow trout (Oncorhynchus mykiss). VRF. 6, 63-67. [ Links ]
Hamman, H.J., 2008. Composition and application of Aloe vera leaf gel. Molecules 13, 1599-1616. [ Links ]
Heidarieh, M., Mirvaghefi, R.A., Sepahil, A., Sheikhzadel, N., Shahbazfar, A.M. & Akbari, M., 2013.Effects of dietary Aloe vera on growth performance, skin and gastrointestine morphology in rainbow trout (Oncorhynchus mykiss). Turk. J. Fish. Aquat. Sci. 13, 367-373. [ Links ]
Ismat, N., Ashraf, M., Naeem, M. & Rehman, M., 2013. Effect of different feed ingredients on growth and level of intestinal enzyme secretions in juvenile Labeo rohita, Catlacatla, Cirrhinusmrigala and Hypophthalmicthysmolitrix. Int. J. Aquacult.3, 85-91. [ Links ]
Jobling, M., 2001. Nutrient partitioning and the influence of feed composition on body composition. In: D. Houlihan (ed.). Food intake in fish. Blackwell Science, Oxford, UK, pp. 354-75. [ Links ]
Langmead, L., Makins, R.J. & Rampton, D.S., 2004. Anti-inflammatory effects of Aloe vera gel in human colorectal mucosa in vitro. Aliment. Pharmacol. Ther. 19, 521-527. [ Links ]
Madan, J., Sharma, A.K., Inamdar, N., Rao, H.S. & Singh, R., 2008. Immunomodulatory properties of Aloe vera gel in mice. Int. J. Green Pharma.2, 152-154. [ Links ]
Mahdavi, M., Hajimoradloo, A. & Ghorbani, R., 2013.Effect of Aloe vera extract on growth parameters of common carp (Cyprinus carpio). World J. of Med. Sci. 9, 55-60. [ Links ]
Mehala, C. & Moorthy, M., 2008. Production performance of broilers fed with Aloe vera and Curcuma longa (turmeric). Int. J. Poult. 7, 852-856. [ Links ]
Moorthy, M., Saravanan, S., Mehala, C., Ravi, S., Ravikumar, M., Viswanathan, K. & Edwin, S.C., 2009. Performance of single comb white leghorn layers fed with Aloe vera, Curcuma longa (turmeric) and probiotic. Int. J. Poult. Sci. 8, 775-778. [ Links ]
Ojha, L.M., Chadha, K.N., Saini, P.V., Damroy, S., Gupta, P.C. & Savant, B.P. 2014. Effect of ethanolic extract of Pedalium murex on growth and haematological parameters of Labeo rohita. Proceedings of the National Academy of Sciences, India. Section B: Biological Sciences 84, 997-1003. [ Links ]
Olagunju, A., Muhammad, A., Mada, S.B., Mohammed, A., Mohammed, A.H. & Mahmoud, K.T., 2012. Nutrient composition of Tilapia zilli, hemisynodontismembranacea, Clupeaharengus and Scomberscombrus consumed in Zaria. World J. Life Sci. Medical Research 2, 16-19. [ Links ]
Omolara, O.O. & Omotayo, O.D., 2008. Preliminary studies on the effect of processing methods on the quality of three commonly consumed marine fishes in Nigeria. J. Biokemistri. 21, 1-7. [ Links ]
Oyewopo, A.O., Oremosu, A.A., Akang, E.N., Noronha, C.C. & Okanlawon, A.O., 2011. Effects of Aloe vera (Aloe Barbadensis) aqueous leaf extract on testicular weight sperm count and motility of adult male Sprague-Dawley rats. J. Am. Sci. 7, 31-34. [ Links ]
Puvabanditsin, P. & Vongtongsri, R., 2005.Efficacy of Aloe vera cream in prevention and treatment of sunburn and suntan. J. Med. Assoc. Thai. 88, 173-176. [ Links ]
Rico, A., Phu, M.T., Satapornvanit, K., Min, J., Shahabuddin, A.M., Henriksson, P.J.G., Murray, F.J., Little, D.C., Dalsgaard, A. & Van der Brink, P.J., 2013. Use of veterinary medicines, feed additives and probiotics in four major internationally traded aquaculture species farmed in Asia. Aquaculture. 412, 231-243. [ Links ]
Sahu, K.P., Giri, D.D., Singh, R., Pandey, P., Gupta, S., Shrivastava, K.A., Kumar, A. & Pandey, D.K., 2013. Therapeutic and medicinal uses of Aloe vera: A review. Pharmacol. Pharm. 4, 599-610. [ Links ]
Shalaby, M.A., Khattab, Y.A. & Abdel, R.A.M., 2006. Effects of garlic (Allium sativum) and chloramphenicol on growth performance, physiological parameters and survival of Nile tilapia (Oreochromis niloticus). J. Venom. Anim. Toxins Incl. Trop. Dis. 12, 172-201. [ Links ]
Tengjaroenkul, B., Smith, B.J., Caceci, T. & Smith, S.A. 2000. Distribution of intestinal enzyme activities along the intestinal tract of cultured Nile tilapia, Oreochromis niloticus L. Aquaculture. 182, 317- 327. [ Links ]
Vasudevam, R.Y., Das, B.K., Jyotyrmayee, P. & Chakrabarti, R., 2006. Effect of Achyranthes aspera on the immunity and survival of Labeo rohita infected with Aeromonas hydrophila [J]. Fish Shellfish Immunol.20, 263-273. [ Links ]
Wu, Y.R., Gong, Q-f., Liang, W-w., Chen, M. & He, R.J., 2013. Effect of Sophora flavescens on non-specific immune response of tilapia (GIFT- Oreochromis niloticus) and disease resistance against Streptococcus agalactiae [J]. Fish Shellfish Immunol. 34, 220-227. [ Links ]
Xie, J., Liu, B., Zhou, Q., Su, Y., He, Y., Pan, L., Ge, X. & Xu, P., 2008. Effects of anthraquinone extract from rhubarb Rheum officinale bail on the crowding stress response and growth of common carp Cyprinus carpio var. Jian. Aquaculture. 281, 5-11. [ Links ]
Zahran, E., Risha, E., AbdelHamid, F. & Mahgoub, A.H. 2014. Effects of dietary Astragalus polysaccharides (APS) on growth performance, immunological parameters, digestive enzymes, and intestinal morphology of Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol.38, 149-157. [ Links ]
Zeitoun, I.H., Ullrey, D.E., Magae, D.E., Gill, J.L. & Bergen, W.G., 1976. Quantifying nutrient requirements of fish. J. Fisheries Res. Board of Canada. 33,167- 172. [ Links ]
Zheng, Z.I., Tan, J.Y.W., Liu, H.Y., Zhou, X.H., Xiang, X. & Wang, K.Y., 2009.Evaluation of oregano essential oil (Origanum heracleoticum L.) on growth, antioxidant effect and resistance against Aeromonas hydrophila in channel catfish (Ictalurus punctatus). Aquaculture. 292, 214-218. [ Links ]
Received 19 April 2017
Accepted 19 September 2017
First published online 16 October 2017
# Corresponding author: ngabriel@unam.na


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}