










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.47 n.5 Pretoria 2017
http://dx.doi.org/10.4314/sajas.v47i5.7
ARTICLES
Ractopamine-induced changes in sarcoplasmic proteome profile of post-rigor pork semimembranosus muscle
J. WuI, II; M.N. NairI; S.P. SumanI, #; S. LiI; X. LuoII; C.M. BeachIII; B.M. BohrerIV, V; D.D. BolerIV, VI
IDepartment of Animal and Food Sciences, University of Kentucky, Lexington, KY 40546, USA
IICollege of Food Science and Engineering, Shandong Agricultural University, Taian, Shandong, 271018, China
IIIProteomics Core Facility, University of Kentucky, Lexington, KY 40506, USA
IVDepartment of Animal Sciences, The Ohio State University, Columbus, OH 43210, USA
VCurrent Address: Department of Food Science, University of Guelph, Guelph, Ontario N1G 2W1, Canada
VICurrent Address: Department of Animal Sciences, University of Illinois, Urbana, IL 61801, USA
ABSTRACT
Ractopamine is a beta-adrenergic agonist that increases leanness and carcass weight in finishing pigs. Our previous study observed that dietary ractopamine increased the abundance of several glycolytic enzymes in the sarcoplasmic proteome of post-rigor pork longissimus thoracis muscle. Pork semimembranosus is an economically important muscle and demonstrates differences in biochemistry compared with longissimus thoracis. Nonetheless, the effects of ractopamine on sarcoplasmic proteome of semimembranosus have not been evaluated yet. Therefore, this study examined the influence of ractopamine on sarcoplasmic proteome of post-rigor pork semimembranosus. Analyses of sarcoplasmic proteome of semimembranosus muscles from control (CON; diet without ractopamine) and ractopamine-fed (RAC; 7.4 mg/kg for 14 days followed by 10.0 mg/kg for 14 days) barrows revealed that haemoglobin subunit beta, alpha-crystallin B, and titin fragments were over-abundant in CON. In contrast, myosin light chain 1/3 and tripartite motif-containing protein 72 were over-abundant in RAC. The low abundance of haemoglobin subunit beta and alpha crystallin B in RAC could be attributed to fibre type shift (from oxidative to glycolytic) in response to ractopamine. The over-abundance of MLC 1/3 and tripartite motif-containing protein 72 in RAC could be due to the increased myofibrillar protein synthesis and muscle mass in ractopamine-fed pigs. Dietary ractopamine decreased the abundance of sarcoplasmic proteins involved in oxygen transport and chaperone activity, but increased the abundance of proteins involved in muscle contraction and plasma membrane repair in pork semimembranosus muscle.
Keywords: Pork, ractopamine, sarcoplasmic proteome, semimembranosus
Introduction
The consumer demand for meat as a source of high quality animal protein is increasing (Lusk, 2013) and the applications of growth technologies in livestock production play a critical role in achieving global food security (Johnson et al., 2013; Dunshea et al., 2016). Beta-adrenergic agonists are oral growth promotants approved for use in food animals (Johnson et al., 2013). Ractopamine is a beta-agonist approved for use in swine industry to increase growth rate, feed efficiency, carcass yield, and leanness in finishing pigs (Apple et al., 2007; Dunshea, 2012; Pompeu et al., 2017) in more than 25 countries, including the United States, Canada, Australia, Mexico, Philippines, Brazil, sand South Korea (AMI, 2012). Dietary ractopamine increases leanness (Boler et al., 2011) and carcass weight (Stites et al., 1991; Armstrong et al., 2004; Fernandez-Duenas, 2008; Needham & Hoffman, 2015) in pigs, and the increased carcass weight is a consequence of the increase in myofibrillar protein synthesis (Adeola et al., 1992).
Proteomic tools are applied to characterize the role of muscle proteome in meat quality (Suman et al., 2014; Gobert et al., 2014; Joseph et al., 2015; Schilling et al., 2017). The sarcoplasmic proteome, comprising 30% of total muscle proteins, consists of enzymes and proteins critical to metabolism in live skeletal muscle and biochemical processes influencing the properties of post-rigor muscle as food (Scopes, 1970). In this regard, our recent study examined the effect of ractopamine on the sarcoplasmic proteome of post-rigor pig longissimus thoracis and reported that ractopamine increased the abundance of glycolytic enzymes and chaperones (Costa-Lima etal., 2015).
Longissimus thoracis et lumborum and semimembranosus are two economically important pork muscles (Jones & Burson, 2000), which demonstrate differences in biochemical properties, such as pH, mineral content, fatty acid profile, transcriptome, and endogenous antioxidants (Huff-Lonergan et al., 2002; Purchas et al., 2009; Herault et al., 2014). Therefore, it is possible that the ractopamine-induced proteome changes in post-rigor pork longissimus thoracis could be different from those in semimembranosus. Nonetheless, the influence of ractopamine on sarcoplasmic proteome of pork semimembranosus is yet to be evaluated. Therefore, the aim of this study was to examine the effect of ractopamine on sarcoplasmic proteome of post-rigor pork semimembranosus muscle.
Materials and Methods
The animal care protocol (Number: 2011A00000141) was approved by the Institutional Animal Care and Use Committee at The Ohio State University (Columbus, OH, USA). Purebred Berkshire pigs were used in this study because they are often used in branded pork programs (McMullen, 2006). Two-hundred purebred Berkshire pigs (barrows and gilts; average initial bodyweight of 68.9 kg) were used as previously described (Bohrer et al., 2013). All animals were raised under similar conditions at The Ohio State University Western Agricultural Research Station (South Charleston, OH, USA). The pigs were stratified based on initial body weight over 2 blocks and were housed in mixed-sex pens, and pens served as replicates. Each block consisted of 10 pens (5 pens x 2 dietary treatments). Within each dietary treatment, 4 pens had 6 barrows and 4 gilts, whereas 1 pen contained 5 barrows and 5 gilts. Overall pen size was 16.25 m2 (including 3.9 m2 slatted-floor area), with each animal receiving approximately 1.63 m2 floor space. Each pen had a double-nipple water drinker and a 4-hole single-sided box feeder that provided a total of 1.22 m of linear feeder space. Pigs were housed in a curtain-sided and naturally ventilated barn and were provided ad libitum access to feed and water throughout the finishing.
Pigs were provided a 14 d adaptation period prior to the start of the treatment diets. Within each block, pigs in 5 pens were finished on a step-up ractopamine (Paylean; Elanco Animal Health, Greenfield, IN, USA) diet (RAC; 171 g/kg crude protein and 10.4 g/kg total lysine) with 7.4 mg/kg ractopamine for 14 d followed by 10 mg/kg ractopamine for the last 14 d prior to slaughter. The pigs in the other 5 pens within the block were finished on a control diet (CON; 131 g/kg crude protein and 7.6 g/kg total lysine) with 0 mg/kg ractopamine. Diets were analysed to ensure ractopamine inclusion levels were within acceptable tolerances (75 to 125%) of the claim for each diet.
Previous research documented that the effect of ractopamine on color and texture of ready-to-eat pork products (i.e. frankfurters) is sex-specific (Costa-Lima et al., 2014). Therefore, to avoid any potential effect of sex on skeletal muscle proteome, only barrows were selected for proteomic investigation. One barrow was randomly selected from each of the 10 pens in CON (104 kg average body weight) and RAC (107 kg average body weight) at the end of the 28-day feeding period, and these 20 pigs were transported to The Ohio State University Meat Science Laboratory. This approach provided 10 replicates (n = 10) for proteome analysis, of which nine were essentially the same animals used in our previous work (Costa-Lima et al., 2015) on the effect of ractopamine on proteome of pork longissimus thoracis. The pigs were kept overnight in lairage with free access to water, but with no access to feed, before being humanely slaughtered. The carcasses were chilled for 24 h at 4 °C before fabrication. Fresh hams from the right side of the carcasses were collected. From the centre of the inside portion of the hams (IMPS # 402F), a 2.54-cm thick slice of semimembranosus muscle was removed. The muscle samples were individually vacuum-packaged, frozen at -80 °C, and transported in dry ice to the University of Kentucky (Lexington, KY, uSa) for proteome analysis.
Sarcoplasmic proteome was extracted according to the previously described method (Joseph et al., 2012). Five grams of frozen muscle samples were homogenized in 25 mL ice-cold extraction buffer (40 mM Tris, 2 mM EDTA, and pH 8.0). The homogenate was centrifuged at 10,000 × g for 15 min at 4 °C. The supernatant (sarcoplasmic proteome extract) was filtered and utilized. The protein concentration of the sarcoplasmic proteome extract was determined using the Bradford assay (Bio-Rad, Hercules, CA, USA). An aliquot corresponding to 900 µg of protein was mixed with rehydration buffer (optimized to 7 M urea, 2 M thiourea, 40 g/L CHAPS, 20 mM DTT, 5 g/L Bio-Lyte 5/8 ampholyte (Bio-Rad, Hercules, CA, USA), and 0.01 g/L Bromophenol blue) and was loaded onto immobilized pH gradient (IPG) strips (pH 5-8, 17 cm). Gels were subjected to passive rehydration for 16 h, and then subjected to first-dimension isoelectric focusing (IEF) in a Protean IEF cell system (Bio-Rad, Hercules, CA, USA) applying a linear increase in voltage initially and a final rapid voltage ramping to reach a total of 80 kVh. Subsequently, the IPG strips were equilibrated in SDS-containing buffers, first with equilibration buffer I (containing 6 M urea, 0.375 M Tris-HCl, pH 8.8, 20 g/L SDS, 200 g/L glycerol, 20 g/L DTT) for 15 min, followed by equilibration buffer II (containing 6 M urea, 0.375 M Tris-HCl, pH 8.8, 20 g/L SDS, 200 g/L glycerol, 25 g/L iodoacetamide) for 15 min. The proteins were resolved in the second dimension on 13.5% SDS-PAGE (38.5:1 ratio of acrylamide to bis-acrylamide) using Protean II XL system (Bio-Rad, Hercules, CA, USA). The gels were stained using Colloidal Coomassie Blue for 48 h and destained until background was cleared. Samples of both treatments (CON and RAC) were run under the same conditions (2 gels/pig), resulting in a total of 40 gels.
Digital images of the stained gels were captured using Versa Doc (Bio-Rad, Hercules, CA, USA) and analysed using PDQuest (Bio-Rad, Hercules, CA, USA). Images were first subjected to automatic spot detection and matching optimized by the aid of landmark protein spots, and the matched spots were normalized (Meunier et al., 2005). A total of 182 spots were identified and evaluated in the image analyses. For each spot in a given sample, spot quantity values in duplicate gels were averaged for statistical analysis (Joseph et al, 2012). A protein spot was considered to be differentially abundant when it was associated with 1.5-fold intensity difference and 5% significance (P <0.05) level in a pairwise Student's t-test (Joseph et al., 2012).
The spots exhibiting differential abundance between the treatments were excised from the gel using pipet tips, placed in microtubes for destaining by two 30 min washes with 50 mM NH4HCO3/500 g/L CH3CN, vortexed for 10 min, and dried in a vacuum centrifuge. The respective spot was excised from the counterpart treatment to confirm the match. Proteins in the gel fragment were reduced by reaction with 10 mM DTT in 50 mM NH4HCO3 solution and incubation at 57 °C for 30 min. The supernatant was discarded, and the proteins (present at the gel piece) were alkylated by addition of 50 mM NH4HCO3 containing 50 mM iodoacetamide and incubated for 30 min at 25 °C without exposure to light. Further, the gel piece was washed twice with 50 mM NH4HCO3 and once with CH3CN, and then partially dried in a vacuum centrifuge. The dried gel piece was rehydrated with a solution of 40 mM NH4HCO3/90 g/L CH3CN, containing 20 mg/L of proteomic grade trypsin (Sigma, St. Louis, MO, USA) on ice for 1 h. An additional volume of 40 mM NH4HCO3/90 g/L CH3CN was added to cover the sample, and the microtube was incubated for 18 h at 37 °C. Peptides were extracted from the gel piece in 0.1 % trifluoroacetic acid by sonication for 10 min followed by vortexing for 10 min, and then the extraction was repeated using a solution on 500 g/L acetonitrile containing 1 g/L trifluoroacetic acid. The extracts were combined, and the volume was reduced to remove most of the acetonitrile. Peptide extracts were desalted and concentrated using solid phase extraction using 1 mm of Empore C-18 (3M, St. Paul, MN, USA) packed in a 0.1 to 10 µL pipet tip (Sarstedt, Newton, NC, USA). The peptides were eluted in 5 µL of 500 g/L CH3CN/0.1% trifluoroacetic acid.
An aliquot of 0.3 µL of the desalted peptide extract was spotted onto an Opti-ToF 384 well insert (AB Sciex, Foster City, CA, USA) with 0.3 µL of 5 g/L α-cyano-4-hydroxycinnamic acid (Aldrich, St. Louis, MO, USA) in 50 g/L CH3CN/1 g/L trifluoroacetic acid. Crystallized samples were washed with cold 1 g/L trifluoroacetic acid and were analysed using a 4800 MALDI TOF-TOF Proteomics Analyzer (AB Sciex, Foster City, CA, USA). An initial MALDI MS spectrum was acquired for each spot (400 laser shots/spectrum), and a maximum of 15 peaks with a signal-to-noise ratio of more than 20 were automatically selected for MS-MS analysis (1000 laser shots/spectrum) by post-source decay. The peak lists from the MS-MS spectra were submitted for similarity search in the National Center for Biotechnology Information (NCBI) database using Protein Pilot version 4.0 (AB Sciex, Foster City, CA, USA) employing relevant search parameters (Search type: identification; Enzyme: trypsin; Database: porcine nCbi nr; Search effort: thorough; Unused cut off > 1.30, 95% confidence) to identify the proteins. A protein was assigned to be identified if it had at least 2.00 unused cut off value and at least two peptides were identified with 99% confidence.
Results and Discussion
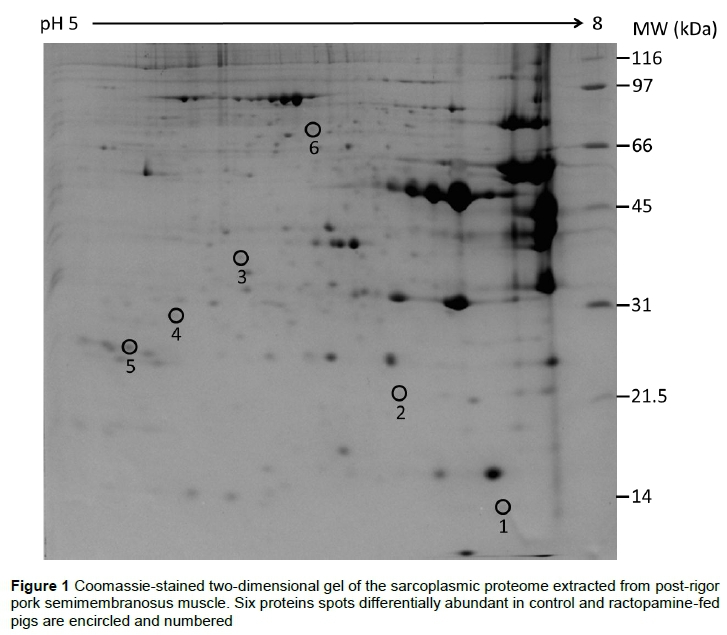
Gel image analyses identified 6 differentially abundant protein spots (Figure 1). Four protein spots were over-abundant (P <0.05) in CON, whereas 2 spots were over-abundant (P <0.05) in RAC. The identity of these proteins and their functional roles are listed in Table 1.
Haemoglobin subunit beta was over-abundant in the CON group (Table 1). Haemoglobin is a tetrameric heme protein comprising 2 alpha (141 residues each) and 2 beta (146 residues each) subunits, and is the major oxygen carrier protein. Based on metabolism, skeletal muscles fibres are classified into 3 types, namely oxidative, oxido-glycolytic, and glycolytic (Peter et al., 1972). Ractopamine functions as a repartitioning agent in skeletal muscles and activates beta-adrenergic receptors, leading to fibre type shifting from oxidative to glycolytic, thereby promoting glycolytic metabolism (Aalhus et al, 1992; Depreux et al., 2002; Gunawan et al., 2007). The decreased abundance of haemoglobin in the RAC group could possibly be attributed to this shift in metabolism from oxidative to glycolytic metabolism in response to ractopamine feeding. Glycolytic metabolism has lower oxygen demand compared to oxidative, which minimizes the necessity for haemoglobin to transport oxygen to the skeletal muscles.
Haemoglobin is a minor contributor to color of fresh meats (Mancini & Hunt, 2005; Suman & Joseph, 2013) and previous proteomic investigation on pork semimembranosus sarcoplasm reported that haemoglobin was of greater abundance in dark muscles than in light muscles (Sayd et al., 2006). Several previous studies observed that fresh meat from pigs fed ractopamine had lower a* values (redness) than meat from animals raised on control diets (Uttaro et al., 1993; Armstrong et al., 2004; Carr et al., 2005a; 2005b; Apple et al., 2008). The decreased abundance of haemoglobin subunit beta in the RAC group suggested that ractopamine feeding may cause a decrease in a* value of pork (Aalhus et al., 1990), attributing to the shift from intermediate fibres to white fibres (Depreux et al., 2002; Gunawan et al., 2007).
Spots 3 and 4 (Fig. 1) were identified as titin fragments and were over-abundant in the CON group (Table 1). Titin is the largest protein in mammals and has a molecular weight of approximately 3,800 kDa; it is the third most abundant myofibrillar protein (Labeit et al., 1997). Although titin is a myofibrillar protein, the appearance of its fragments in sarcoplasm could be due to post-mortem proteolysis (Taylor et al., 1995). The low sequence coverage of titin (Table 1) could be attributed to multiple proteases, such as calpain proteinases (Huff-Lonergan et al., 2010; Lian et al., 2013), lysing the protein into small fragments and the large size (more than 34,000 amino acids) of titin molecules. Degradation of titin has been reported to increase meat tenderness (Huff-Lonergan et al., 1995), and the over-abundance of titin fragments in CON could be due to decreased proteolysis in the RAC group. In support, dietary ractopamine has been reported to decrease proteolytic activity in pork longissimus (Xiong et al., 2006).
Myosin light chain (MLC) 1/3 was over-abundant in the RAC group (Table 1). These findings are also supported by Costa-Lima et al. (2015), who reported a greater abundance of MLC 1/3 in longissimus thoracis of ractopamine-fed pigs than in pigs fed a control diet. Myosin is a myofibrillar protein, and the hexameric myosin molecule consists of 2 heavy chains (MHC), 2 regulatory light chains (RLC), and 2 alkali light chains (MLC 1, MLC 3) in fast-twitch skeletal muscles (Frank & Weeds, 1974). Two alkali light chains (MLC 1 and MLC 3) observed in the sarcoplasmic fraction in the present study could be the products of proteolytic cleavage, which releases the light chain sub-units from the actomyosin complex to sarcoplasm (Gerlemann et al., 2014).
Previous studies documented that dietary ractopamine increased myofibrillar protein synthesis in pork longissimus (Adeola et al., 1992). Furthermore, Helferich et al. (1990) observed an increase in the MLC 1/3 RNA expression in ractopamine-fed porcine longissimus indicating protein accretion and increase in muscle mass. The greater abundance of MLC 1/3 in the RAC group than in the CON group observed in the present study could be due to the increased myofibrillar protein synthesis in response to ractopamine. Several proteomic investigations have been undertaken to identify the relationship between myosin light chains and meat quality. A positive correlation between MLC 1 and shear force has been observed in pork longissimus (Hwang et al., 2005). Furthermore, in beef longissimus thoracis, MLC 1 was of greater abundance in tough meat than in tender meat (Bjarnadottir et al., 2012).
Tripartite motif-containing protein 72 plays a critical role in plasma membrane repair and was over-abundant in the RAC group (Table 1). The cellular mechanism for plasma membrane repair and remodelling, to maintain cellular homeostasis in response to various physiological and pathological conditions, is conserved across eukaryotes (Han & Campbell, 2007). Tripartite motif-containing protein 72 (mitsugumin-53) is an essential component involved in plasma membrane repair in mouse skeletal muscle (Cai et al., 2009). It is well documented that dietary ractopamine increases carcass weight in finishing pigs (Stites et al., 1991; Armstrong et al., 2004; Fernandez-Duenas et al., 2008). Furthermore, an increase in the weight of inside ham (consisting of semimembranosus muscle) was observed in ractopamine-fed pigs (Boler et al., 2011). The increased muscle growth due to ractopamine could possibly demand increased levels of plasma membrane repair proteins to maintain homeostasis. Therefore, the over-abundance of tripartite motif-containing protein 72 in RAC could be attributed to the increased muscle mass accretion in ractopamine-fed animals (Bohrer et al., 2013).
Conclusion
The results of the present study suggest that dietary ractopamine influences the abundance of proteins involved in oxygen transport, chaperone activity, and plasma membrane repair in sarcoplasmic proteome of pork semimembranosus muscle. This change in sarcoplasmic proteome profile may be primarily due to the previously documented muscle fibre type shift (from oxidative to glycolytic) and the increased muscle growth in response to ractopamine feeding. Additionally, the differences between the effects of ractopamine on sarcoplasmic proteome in semimembranosus in the present study and those previously reported in longissimus thoracis (Costa-Lima et al., 2015) warrant further studies on muscle-specific effects of this beta agonist in pork.
Acknowledgments
This is publication number 17-07-049 of the Kentucky Agricultural Experiment Station and is published with the approval of the director. This work is supported by the National Institute of Food and Agriculture, U.S. Department of Agriculture, Hatch-Multistate Project 1008755. Mass spectrometric analysis was performed at the University of Kentucky's Proteomics Core Facility, supported in part by funds from the Office of the Vice President for Research.
Authors' Contributions
Conception and design - SPS, DDB, & XL; Data collection and analyses- JW, BMB, MNN, SL, & CMB; Drafting of paper - JW & MNN; Critical revision and final approval of version to be published - SPS.
Conflict of Interest Declaration
The authors declare that they have no affiliations with any organization or entity with any financial or non-financial interest that could bias the subject matter and outcomes discussed in this manuscript.
References
Aalhus, J.L., Jones, S.D.M., Schaefer, A.L., Tong, A.K.W., Robertson, W.M., Merrill, J.K. & Murray, A.C., 1990. The effect of ractopamine on performance, carcass composition and meat quality of finishing pigs. Can. J. Anim. Sci. 70, 943-952. [ Links ]
Aalhus, J.L., Schaefer, A.L., Murray, A.C. & Jones, S.D.M., 1992. The effect of ractopamine on myofibre distribution and morphology and their relation to meat quality in swine. Meat Sci. 31, 397-409. [ Links ]
Adeola, O., Ball, R.O. & Young, L.G., 1992. Porcine skeletal muscle myofibrillar protein synthesis is stimulated by ractopamine. J. Nutr. 122, 488-495. [ Links ]
AMI., 2012. Growth promotants in meat production: Their use and safety. https://www.meatinstitute.org/index.php?ht=a/GetDocumentAction/i/83421 (Accessed May 29, 2017). [ Links ]
Apple, J.K., Maxwell, C.V., Kutz, B.R., Rakes, L.K., Sawyer, J.T., Johnson, Z.B., Armstrong, T.A., Carr, S.N. & Matzat, P.D., 2008. Interactive effect of ractopamine and dietary fat source on pork quality characteristics of fresh pork chops during simulated retail display. J. Anim. Sci. 86, 2711-2722. [ Links ]
Apple, J.K., Rincker, P.J., McKeith, F.K., Carr, S.N., Armstrong, T.A. & Matzat, P.D., 2007. Review: Meta-analysis of the ractopamine response in finishing swine. Prof. Anim. Sci. 23, 179-196. [ Links ]
Armstrong, T.A., Ivers, D.J., Wagner, J.R., Anderson, D.B., Weldon, W.C. & Berg, E.P., 2004. The effect of dietary ractopamine concentration and duration of feeding on growth performance, carcass characteristics, and meat quality of finishing pigs. J. Anim. Sci. 82, 3245-3253. [ Links ]
Atomi, Y., Yamada, S., Strohman, R. & Nonomura, Y., 1991. Alpha B-crystallin in skeletal muscle: purification and localization. J. Biochem. 110, 812-822. [ Links ]
Bhat, S.P. & Nagineni, C.N., 1989. Alpha B subunit of lens-specific protein alpha-crystallin is present in other ocular and non-ocular tissues. Biochem. Biophys. Res. Commun. 158, 319-325. [ Links ]
Bjarnadottir, S.G., Hollung, K., Hoy, M., Bendixen, E., Codrea, M.C. & Veiseth-Kent, E., 2012. Changes in protein abundance between tender and tough meat from bovine longissimus thoracis muscle assessed by isobaric Tag for Relative and Absolute Quantitation (iTRAQ) and 2-dimensional gel electrophoresis analysis. J. Anim. Sci. 90, 2035-2043. [ Links ]
Bohrer, B.M., Kyle, J.M., Little, K.L., Zerby, H.N. & Boler, D.D., 2013. The effects of a step-up ractopamine feeding program on growth performance and low-sodium ham characteristics of purebred Berkshire pigs. J. Anim. Sci. 91, 5535-5543. [ Links ]
Boler, D.D., Holmer, S.F., Duncan, D.A., Carr, S.N., Ritter, M.J., Stites, C.R., Petry, D.B., Hinson, R.B., Allee, G.L., McKeith, F.K. & Killefer, J., 2011. Fresh meat and further processing characteristics of ham muscles from finishing pigs fed ractopamine hydrochloride. J. Anim. Sci. 89, 210-220. [ Links ]
Cai, C., Masumiya, H., Weisleder, N., Matsuda, N., Nishi, M., Hwang, M., Ko, J.K., Lin, P., Thornton, A., Zhao, X., Pan, Z., Komazaki, S., Brotto, M., Takeshima, H. & Ma, J., 2009. MG53 nucleates assembly of cell membrane repair machinery. Nat. Cell Biol. 11, 56-64. [ Links ]
Carr, S.N., Ivers, D.J., Anderson, D.B., Jones, D.J., Mowrey, D.H., England, M.B., Killefer, J., Rincker, P.J. & McKeith, F.K., 2005a. The effects of ractopamine hydrochloride on lean carcass yields and pork quality characteristics. J. Anim. Sci. 83, 2886-2893. [ Links ]
Carr, S.N., Rincker, P.J., Killefer, J., Baker, D.H., Ellis, M. & McKeith. F.K., 2005b. Effects of different cereal grains and ractopamine hydrochloride on performance, carcass characteristics, and fat quality in late-finishing pigs. J. Anim. Sci. 83, 223-230. [ Links ]
Costa-Lima, B.R., Suman, S.P., Li, S., Beach, C.M., Silva, T.J., Silveira, E.T., Bohrer, B.M. & Boler, D.D., 2015. Dietary ractopamine influences sarcoplasmic proteome profile of pork longissimus thoracis. Meat Sci. 103, 7-12. [ Links ]
Costa-Lima, B.R.C., Canto, A.C.V.C.S., Suman, S.P., Conte-Junior, C.A., Silveira, E.T.F. & Silva, T.J.P., 2014. Sex-specific effect of ractopamine on quality attributes of pork frankfurters. Meat Sci. 96, 799-805. [ Links ]
Depreux, F.F.S., Grant, A.L., Anderson, D.B. & Gerrard, D.E., 2002. Paylean alters myosin heavy chain isoform content in pig muscle. J. Anim. Sci. 80, 1888-1894. [ Links ]
Dunshea, F.R., 2012. Emerging technologies with the potential to improve feed efficiency in swine. In: J. F. Patience, editor, Feed efficiency in swine. Wageningen Academic Publishers, The Netherlands. p. 259-275. [ Links ]
Dunshea, F.R., D'Souza, D.N. & Channon, H.A., 2016. Metabolic modifiers as performance-enhancing technologies for livestock production. Animal Front. 6, 6-14. [ Links ]
Fernandez-Duenas, D.M., Myers, A.J., Scramlin, S.M., Parks, C.W., Carr, S.N., Killefer, J. & McKeith, F.K., 2008. Carcass, meat quality, and sensory characteristics of heavy body weight pigs fed ractopamine hydrochloride (Paylean). J. Anim. Sci. 86, 3544-3550. [ Links ]
Frank, G. & Weeds, A.G., 1974. The amino-acid sequence of the alkali light chains of rabbit skeletal-muscle myosin. Eur. J. Biochem. 44, 317-334. [ Links ]
Gerlemann, G.D., Allee, G.L., Rincker, P.J., Ritter, M.J., Boler, D.D. & Carr, S.N., 2014. Impact of ractopamine hydrochloride on growth, efficiency, and carcass traits of finishing pigs in a three-phase marketing strategy. J. Anim. Sci. 92, 1200-1207. [ Links ]
Gobert, M., Sayd, T., Gatellier, P. & Santé-Lhoutellier, V., 2014. Application to proteomics to understand and modify meat quality. Meat Sci. 98, 539-543. [ Links ]
Gunawan, A.M., Richert, B.T., Schinckel, A.P., Grant, A.L. & Gerrard, D.E., 2007. Ractopamine induces differential gene expression in porcine skeletal muscles. J. Anim. Sci. 85, 2115-2124. [ Links ]
Han, R. & Campbell, K.P., 2007. Dysferlin and muscle membrane repair. Curr. Opin. Cell Biol. 19, 409-416. [ Links ]
Helferich, W.G., Jump, D.B., Anderson, D.B., Skjaerlund, D.M., Merkel, R.A. & Bergen, W.G., 1990. Skeletal muscle alpha-actin synthesis is increased pretranslationally in pigs fed the phenethanolamine ractopamine. Endocrinology 126, 3096-3100. [ Links ]
Herault, F., Vincent, A., Dameron, O., Le Roy, P., Cherel, P. & Damon, M., 2014. The longissimus and semimembranosus muscles display marked differences in their gene expression profiles in pig. PLOS ONE 9, e96491. doi: 10.1371/journal.pone.0096491. [ Links ]
Horwitz, J., 1992. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA. 89, 10449-10453. [ Links ]
Huff Lonergan, E., Zhang, W. & Lonergan, S.M., 2010. Biochemistry of postmortem muscle - Lessons on mechanisms of meat tenderization. Meat Sci. 86, 184-195. [ Links ]
Huff-Lonergan, E., Baas, T.J., Malek, M., Dekkers, J.C.M., Prusa, K. & Rothschild, M.F., 2002. Correlations among selected pork quality traits. J. Anim. Sci. 80, 617-627. [ Links ]
Huff-Lonergan, E., Parrish Jr., F.C. & Robson, R.M., 1995. Effects of postmortem aging time, animal age, and sex on degradation of titin and nebulin in bovine longissimus muscle. J. Anim. Sci. 73, 1064-1073. [ Links ]
Hwang, I. H., Park, B. Y., Kim, J. H., Cho, S. H. & Lee, J. M., 2005. Assessment of postmortem proteolysis by gel-based proteome analysis and its relationship to meat quality traits in pig longissimus. Meat Sci. 69, 79-91. [ Links ]
Johnson, B., Ribeiro, F. & Beckett, J., 2013. Application of growth technologies in enhancing food security and sustainability. Anim. Front. 3, 8-13. [ Links ]
Jones, S. J. & Burson. D.E., 2000. Porcine Myology. National Pork Producers Council, Des Moines, IA 1-73. [ Links ]
Joseph, P., Nair, M.N. & Suman, S.P., 2015. Application of proteomics to characterize and improve color and oxidative stability of muscle foods. Food Res. Int. 76, 938-945. [ Links ]
Joseph, P., Suman, S.P., Rentfrow, G., Li, S. & Beach, C.M., 2012. Proteomics of muscle-specific beef color stability. J. Agr. Food Chem. 60, 3196-3203. [ Links ]
Labeit, S., Kolmerer, B. & Linke, W.A., 1997. The giant protein titin. Emerging roles in physiology and pathophysiology. Circ. Res. 80, 290-294. [ Links ]
Lian, T., Wang, L. & Liu, Y., 2013. A New Insight into the Role of Calpains in Post-mortem Meat Tenderization in Domestic Animals: A review. Asian-Australas. J Anim. Sci. 26, 443-454. [ Links ]
Lusk, J.L., 2013. Role of technology in the global economic importance and viability of animal protein production. Anim. Front. 3, 20-27. [ Links ]
Mancini, R. A. & Hunt, M.C., 2005. Current research in meat color. Meat Sci. 71, 100-121. [ Links ]
McMullen, L.K., 2006. Berkshire swine production and marketing: Berkshire niche market opportunity guidelines. Jones County Extension Office, Iowa State Univ., Anamosa, IA. www.iowaporkcongress.org/download/Mcmullen.pdf (Accessed 7 January 2017. [ Links ])
Meunier, B., Bouley, J., Piec, I., Bernard, C., Picard, B. & Hocquette, J.F., 2005. Data analysis methods for detection of differential protein expression in two-dimensional gel electrophoresis. Anal. Biochem. 340, 226-230. [ Links ]
Needham, T. & Hoffman, L.C., 2015. Carcass traits and cutting yields of entire and immunocastrated pigs fed increasing protein levels with and without ractopamine hydrochloride supplementation. J. Anim. Sci. 93, 4545-4556. [ Links ]
Neufer, P.D. & Benjamin, I.J., 1996. Differential expression of aB-crystallin and hsp27 in skeletal muscle during continuous contractile activity. Relationship to myogenic regulatory factors. J. Biol. Chem. 271, 24089-24095. [ Links ]
Peter, J.B., Barnard, R.J., Edgerton, V.R., Gillespie, C.A. & Stempel, K.E., 1972. Metabolic profiles of three fiber types of skeletal muscle in guinea pigs and rabbits. Biochemistry 11, 2627-2633. [ Links ]
Pompeu, M.A., Rodrigues, L.A., Cavalcanti, L.F.L., Fontes, D.O., & Toral, F.L.B., 2017. A multivariate approach to determine the factors affecting response level of growth, carcass, and meat quality traits in finishing pigs fed ractopamine. J. Anim. Sci. 95, 1644-1659. [ Links ]
Purchas, R.W., Morel, P.C.H., Janz, J.A.M., & Wilkinson, B.H.P., 2009. Chemical composition characteristics of the longissimus and semimembranosus muscles for pigs from New Zealand and Singapore. Meat Sci. 81, 540-548. [ Links ]
Sayd, T., Morzel, M., Chambon, C., Franck, M., Figwer, P., Larzul, C., Le Roy, P., Monin, G., Cherel, P. & Laville, E., 2006. Proteome analysis of the sarcoplasmic fraction of pig semimembranosus muscle: implications on meat color development. J. Agr. Food Chem. 54, 2732-2737. [ Links ]
Schilling, M.W., Suman S.P., Zhang, X., Nair, M.N., Desai, M.A., Cai, K., Ciaramella, M.A. & Allen, P.J., 2017. Proteomic approach to characterize biochemistry of meat quality defects. Meat Sci. doi: 10.1016/j.meatsci.2017.04.018. [ Links ]
Scopes, R.K., 1970. Characterization and study of sarcoplasmic proteins. In: Briskey, E.J. Cassens, R.G. & Marsh, B.B., editors, The physiology and biochemistry of muscle as a food. University of Wisconsin Press, Madison, WI. p. 471-492. [ Links ]
Stites, C.R., McKeith, F.K., Singh, S.D., Bechtel, P.J., Mowrey, D.H. & Jones, D.J., 1991. The effect of ractopamine hydrochloride on the carcass cutting yields of finishing swine. J. Anim. Sci. 69, 3094-3101. [ Links ]
Suman, S.P. & Joseph, P., 2013. Myoglobin chemistry and meat color. Annu. Rev. Food. Sci. Technol. 4, 79-99. [ Links ]
Suman, S.P., Rentfrow, G., Nair, M.N. & Joseph, P., 2014. Proteomics of muscle- and species-specificity in meat color stability. J. Anim. Sci. 92, 875-882. [ Links ]
Taylor, R.G., Geesink, G.H., Thompson, V.F., Koohmaraie, M. & Goll, D.E., 1995. Is Z-disk degradation responsible for postmortem tenderization? J. Anim. Sci. 73, 1351-1367. [ Links ]
Uttaro, B.E., Ball, R.O., Dick, P., Rae, W., Vessie, G. & Jeremiah, L.E., 1993. Effect of ractopamine and sex on growth, carcass characteristics, processing yield, and meat quality characteristics of crossbred swine. J. Anim. Sci. 71, 2439-2449. [ Links ]
Xiong, Y.L., Gower, M.J., Li, C., Elmore, C.A., Cromwell, G.L. & Lindemann, M. D., 2006. Effect of dietary ractopamine on tenderness and postmortem protein degradation of pork muscle. Meat Sci. 73, 600-604. [ Links ]
Received 30 May 2017
Accepted 2 August 2017
First published online 25 August 2017
# Corresponding author: spsuma2@uky.edu


{kind=link}
{kind=link}