










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.47 n.3 Pretoria 2017
http://dx.doi.org/10.4314/sajas.v47i2.15
ARTICLES
Effects of restricted feeding and re-alimentation of dietary protein or energy on compensatory growth of sheep
W. AddahI, #; A. AyantundeII; E.K. OkineIII
IDepartment of Animal Science, University for Development Studies, P. O. Box TL 1882, Tamale, Ghana
IIInternational Livestock Research Institute, BP 9478, Ouagadougou, Burkina Faso
IIIDepartment of Biological Sciences, University of Lethbridge, Lethbridge, Alberta, Canada T1K 3M4
ABSTRACT
The study investigated the effects of re-alimenting dietary protein or energy on growth, carcass characteristics and meat eating quality parameters of sheep. Twenty-seven intact rams (~9 months; 11.3 ± 0.5 kg) were randomly divided into three groups. Each group was fed a maintenance diet (MT) containing, on dry matter (DM) basis, 105 g/kg crude protein (CP) and 8.4 MJ/kg DM metabolizable energy (ME) for 30 days. Thereafter, they were continually fed the same MT or re-alimented with a high protein diet (HP) containing 169 g/kg DM CP and 9.3 MJ/kg ME or a high energy diet (HE) containing 123 g/kg DM CP and 10.6 MJ/kg ME for an additional 30 days to determine the effects of re-alimentation of protein or energy on their growth performance and carcass characteristics. During the initial 30-day period, DM intake (DMI) and growth performance were similar among the three groups. However, upon re-alimentation, average daily gain (ADG) and feed efficiency of sheep re-alimented with HP were greater than those maintained continually on MT or re-alimented with HE. Sheep on HP had higher feed efficiency, ADG and heavier carcasses than those fed MT or re-alimented with HE during the whole 60-day period. Growth of most viscera was less responsive to the restriction-re-alimentation feeding regimen except for the weights of the lungs, heart and intestines. Meat from sheep re-alimented with HE had a more intense 'sheepy' flavour than those fed MT or re-alimented with HP, but juiciness and tenderness were not affected. The higher ADG of sheep re-alimented with protein may be related more to enhanced efficiency of feed utilization than to higher DMI.
Keywords: average daily gain, feed restriction, nutrient utilization efficiency, visceral organs
Introduction
Sheep are a source of meat, income, food security and short-term cash reserves. In most African cultures of northern Ghana and Nigeria, sheep also have a social benefit, in that live animals have socio-cultural values far beyond their market value in terms of live weight. Unlike goats, they are acceptable for payment of the bride price during marriage and for ritual sacrifices (Okali & Sumberg, 1985). Urban livestock traders in West Africa often buy young animals in the non-festive season and then fatten them for sale at Christmas and the Islamic festival of Idle Fitir when demand for mutton is high (Okali & Upton, 1984; Baah et al., 2012). However, feeding these animals between these periods is often a challenge, because most natural pastures are destroyed by wild bushfires in the dry season, and providing nutritious feed throughout the year is constrained by feed availability and cost (Addah, 2014).
To address these challenges, urban livestock traders and farmers usually restrict dietary nutrient intake during feed unavailability to maintenance levels and re-aliment these nutrients toward the festive season to induce compensatory growth. A 40% restriction of ad libitum intake followed by re-alimentation has been used to induce compensatory growth, improve growth rate, and produce leaner carcasses in growing lambs (Abouheif et al., 2013). Compensatory growth has also been induced to alter the slaughter weight of animals to meet the taste of consumers (Owens et al., 1993). However, it is not known whether re-alimenting dietary energy or protein would induce the necessary compensatory growth and whether the improved growth could be translated into improvement in carcass weight and meat quality. Some previous studies suggested that improved growth performance during the re-alimented period was because of increased appetite and feed intake (Marais et al., 1991), but others (Ryan et al., 1993a; Ryan et al., 1993b; Keogh et al., 2015) hypothesized that reduced maintenance requirements and greater deposition of protein were responsible for compensatory growth during the initial stages of re-alimentation, and compensatory growth in the later stages was attributed to greater DMI. Yet compensatory growth following re-alimentation of nutrients has been reported in sheep with lower feed intake (Kamalzadeh et al., 1997). Various body organs have been found to react differently during restriction and re-alimentation. Early maturing organs such as the head, feet and viscera structures are less affected by restriction-re-alimentation feeding regimes than late-maturing ones such as the liver, gastrointestinal tract, heart and kidneys (Kamalzadeh et al., 1998). The economic benefits of compensatory growth, therefore, have remained doubtful because some earlier studies attributed observed increases in live weight gain to accumulation of water in the gut (Drew & Reid, 1975) and fat in the meat (Turgeon et al., 1986; Hornick et al., 2000). The mechanisms and benefits of the practice of inducing compensatory growth have not been elucidated and conclusively established in sheep. It is not known whether re-alimentation of dietary protein or energy would induce the greatest compensatory growth response and relative effects on meat eating quality parameters. Hence the present study investigated the effects of re-alimenting dietary protein or energy on growth, carcass characteristics and meat eating quality parameters of sheep.
Materials and Methods
The study was conducted at the Ruminant Production Unit of the University for Development Studies, Nyankpala (latitude 9.3965° N; longitude 0.9892° W). The area has two main seasons; the dry season and the rainy season. The study was conducted during the dry season from 28 October to 27 December 2015. Data on environmental conditions during this period were obtained from weather records of Savanna Agricultural Research Institute (SARI, 2015), situated 50 m from the location of the experiment. The data included temperature (18-35 °C); rainfall (20-39 mm); relative humidity (27-40%); and sunshine (80-87%).
A total of 27 intact West African Dwarf growing ram lambs (~9 months) were purchased from a local market in the northern region of Ghana. After seven days of quarantine and prophylactic treatment against worms and ticks, they were castrated by a qualified veterinary technician. The sheep were then identified with plastic identification tags (Fearing Int. Ltd, Northampton, UK) and were all group-fed ad libitum a diet containing crushed corn (380 g/kg), whole cotton seed (420 g/kg), peanut haulms (150 g/kg) and mineralvitamin premix (50 g/kg), which was calculated to supply 9.3 MJ/kg and 125 g/kg CP on a DM basis. A month after castration, they were randomly assigned to nine wooden pens (2.5 m χ 1.5 m) with concrete floors, each pen consisting of three wethers, at the University for Development Studies, Tamale, Ghana.
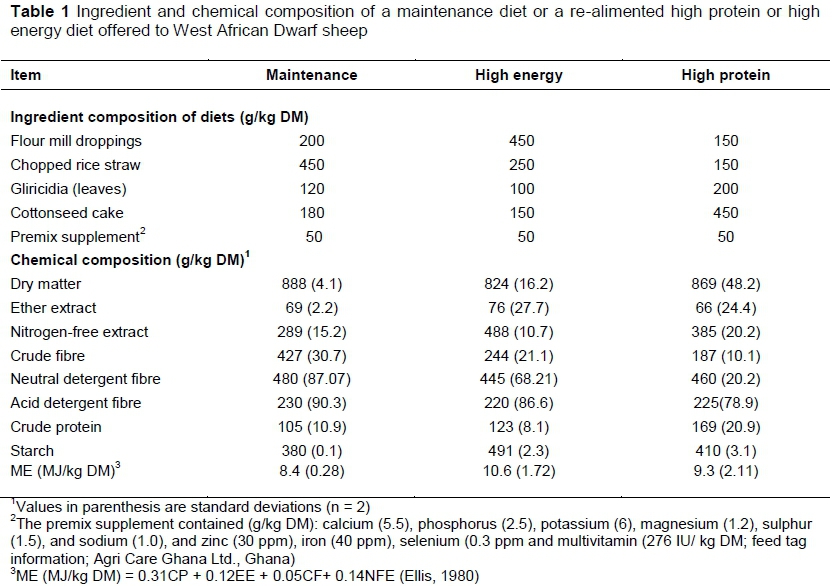
All sheep (11.3 ± 0.5 kg) were then fed an MT diet (10.5% CP; 8.4 MJ/kg ME) (NRC, 2007) for 30 days, after which each pen was randomly assigned to one of three dietary treatments (n = 3) in a completely randomized design in which sheep were fed either the same MT or a re-alimented diet that was higher in protein (HP) (169 g/kg DM CP; 9.3 MJ/kg ME) or in energy (HE) (123 g/kg DM CP; 10.6 MJ/kg ME) for 30 additional days to allow for moderate-to-rapid growth potential (NRC, 2007). Hence dietary CP was increased by 60.9% in the HP diet, and dietary ME was increased 34% in the HE diet compared with MT (Table 1). The MT diet was formulated to allow for body maintenance at ad libitum intake for the 60 days of the experiment. The ingredients and nutrient composition of the diets are indicated in Table 1. Sheep were weighed on two consecutive days at the beginning of the experiment and every two weeks thereafter until the end of the experiment, when two consecutive weights were taken. The average of the consecutive weights at the beginning of the study and at the end were used as the initial and final weights, respectively.
Feed was mixed manually and delivered daily as a total mixed ration. The daily amount of feed offered was recorded and leftovers were collected daily, weighed and sampled before being discarded. Samples of feed and leftovers that were collected daily were composited into bi-weekly samples, subsampled and stored (-40 oC) for subsequent proximate analysis. The DM of each dietary ingredient was also determined (60 oC for 48 h) on weekly samples composited from samples of each ingredient collected daily prior to preparing the ration. This made it possible to adjust the proportions of ingredients in the diet whenever there was an appreciable change in the DM concentration of the ingredients. Animals were offered their feed every morning (08:30-09:15). The quantities of feed offered daily were adjusted to meet appetites of animals and to ensure minimal feed leftovers without limiting intake. Fresh water (~15 L) was supplied daily per pen. Dry matter intake of each pen was calculated as feed DM offered minus DM of the left-overs. The DMI, average daily gain (ADG), and feed efficiency (expressed as ADG/daily DMI) were estimated separately for the periods when the sheep were fed the MT diet and the re-alimented diets (HP and HE), and for the whole 60-day experimental period.
The experimental subjects were cared for and managed with ethical considerations for conducting scientific experiments. The protocol was approved by a peer-review committee of the Department for Animal Science based on the guidelines for conducting research in the Ethics Policy Handbook (ICEIR, 2010) of the University for Development Studies, prior to the commencement of the experiment.
At the end of the feeding trial, six wethers were randomly sampled from each treatment (two per pen) and transported to a meat processing plant at the University for Development Studies, Tamale, Ghana. The wethers were slaughtered according to standard protocol for slaughtering animals at the plant. Briefly, live weights of animals were recorded before being stunned with a captive bolt pistol and bled for approximately 10 min by severing the main jugular vein. Singeing was done with a gas flame and knife to scrub off the hair on the skin. Evisceration was done immediately after singeing, and the warm carcass and visceral weights of each animal were recorded and expressed as proportions of weight at slaughter. Contents of the reticulo-rumen and intestines were emptied before weighing. Hot carcass weight was recorded with kidney, heart, and pelvic fat present. The forestomach (reticulo-rumen) and intestines (large and small intestines) were removed, emptied and weighed separately. Other visceral organs (liver, lungs, heart, kidneys, and spleen) and visceral fats around the reticulo-rumen (omental), intestine (mesenteric) and abdominal-pelvic region (peritoneal fat) were also similarly removed and weighed separately for each carcass. Prior to weighing, visceral organs were trimmed of attached fat.
The carcass of each animal was halved. Each pair of longissimus dorsi from each carcass was removed and stored (1 °C) for 24 h. Each longissimus dorsi was then sliced into two parts. This resulted in 18 parts per dietary treatment. The parts were griddled in an electric oven (Turfbon, Blue Seal, UK) for 45 min, sliced uniformly and wrapped in blind-labelled aluminium foil before being presented to twelve trained sensory panellists. Panel members were selected and trained according to the American Society for Testing and Material (Schaefer, 1979). The slices were scored for flavour intensity, juiciness and tenderness using a 0 to 8 assessment scale previously described by Fisher et al. (2000) and Priolo et al. (2001).
Feed offered and leftovers sampled bi-weekly were used to determine DM. Dry matter of ingredients, feed offered and leftovers was determined in a force-air oven at 60 °C for 48 hours instead of the conventional 105 °C for 24 hours because the diets contained Gliricidia and whole cotton seed (Table 1). These ingredients contain oils, carotenes, vitamins and other volatile compounds. Determining DM at 105 °C for 24 hours usually leads to underestimation of DM concentration because all these compounds that volatilizes at higher temperature are erroneously calculated as moisture when DM of the feed is determined at 105 °C (Porter et al., 1995; Addah et al., 2012).
Feed samples composited into bi-weekly samples were subsampled for proximate analysis according to the official methods of analyses described by Association of Official Analytical Chemists (AOAC, 1990). All analyses were done in duplicate. The protocol was used to determine the concentration of DM, CP calculated as total N χ 6·25 (ID 968.06), ether extract (EE); accelerated solvent extraction (ID 920.39C) (Richter, et al., 1996), crude fibre (CF) (ID 962.09) and nitrogen-free extract (NFE). Nitrogen-free extract was computed using the formula (g/kg DM): NFE = 100 - (CP + CF + EE + ash). All nutrient constituents were expressed on DM basis.
Bi-weekly sub-samples of each diet were also air-dried and ground through a 1-mm screen to analyse ash-inclusive neutral detergent fibre (aNDF) and acid detergent insoluble fibre (ADF), using the Van Soest method (Van Soest, 1991). The neutral detergent solution dissolves the easily digestible plant cell contents (proteins, sugars and lipids) and pectins, leaving a fibrous residue of aNDF, which is primarily cell wall components (cellulose, hemicellulose and lignin). Sodium sulfite also helps remove some nitrogenous matter. The hemicellulose component of aNDF is then dissolved by refluxing again with an acid detergent solution, leaving a residue of cellulose and lignin (ADF). The addition of heat-stable amylase is useful in the removal of starch and inactivation of potential contaminating enzymes. The solution for analysis of aNDF thus had sodium sulfite and α-amylase added to it, whereas ADF was analysed without α-amylase. To determine starch, air-dried feed samples were ball-ground with a mixer mill (MM 400, Retsch Inc. Newtown, Pa, USA). Starch was determined by hydrolysing to α-glucose polymers using amyloglucosidase (Megazyme Int. Ltd., Wicklow, Ireland) plus 1,4 α-D-glucan glucano-hydrolase (Brennfag Canada Inc., Toronto, On., Canada), as described by Herrera-Saldana et al. (1990). Samples were read on a Spectrotroquant Pharo 300 spectrophotometer (JP Selecta SA, Barcelona, Spain) at a wavelength of 490 nm. Metabolizable energy of diets was calculated from the equation ME (MJ/kg DM) = 0.31CP + 0.12EE + 0.05CF+ 0.14NFE (Ellis, 1980) (Table 1).
Data were analysed using the MIXED model procedure of SAS 9.3 (SAS Institute, Cary NC). All data on growth performance (DMI, weight gain, ADG, feed efficiency), carcass weight and meat quality (flavour intensity, juiciness, tenderness) of sheep were analysed for the fixed effects of dietary treatments in a completely randomized design with pen as the experimental unit in the statistical model below:
Yij = μ + Ti + ey
where: Yy is the observation (DMI, weight gain
ADG, feed efficiency, carcass weight, tenderness)
μ is the overall mean effect Ti
Ti is the effect of dietary treatments (MT, HE or HP)
εy is the residual error effect
Differences in least square means for growth performance data were compared with the PDIFF option of LSMEANS. All least square means that showed significant differences were separated by Fisher's pairwise t-test and were declared statistically significant at P <0.05, whereas P <0.10 were reported and discussed as trends.
Results
Differential CP and ME concentrations of HP and HE, respectively, are shown in Table 1. Crude protein content was 64 g/kg DM greater in HP than MT, whereas ME concentration was 2.2 MJ/kg DM higher in HE than the maintenance diet. The DMI and growth performance are shown in Table 2. During the initial 30-day restriction period when the sheep were fed the MT, DMI and weights were similar (P = 0.480 or less) among the three dietary treatments. Upon re-alimentation, DMI was still not different (P = 0.149) among treatments, but ADG was improved (P = 0.001) for sheep re-alimented with the HP compared with those re-alimented with HE or continuously fed the MT. The ADG, however, was higher (P = 0.001) for sheep fed the MT diet than for those re-alimented with the HE. Re-alimentation with the HP improved (P = 0.015) the efficiency of feed utilization compared with re-alimentation with HE or feeding MT, but the difference between HE and MT was not significant (P = 0.700). The efficiency of feed utilization did not differ (P = 0.700) between HE and MT.
The overall (60 days) growth performance of the sheep showed a greater (P = 0.011) ADG for sheep on HP than for those on HE or MT. Even though ADG was not significantly different between MT and HE, there was a trend (P = 0.111) towards greater ADG for sheep fed the MT diet (0.16 kg/d) than those fed the HE diet (0.12 kg/d). Consistent with growth differences during the re-alimented period, sheep fed the HP utilized their feed more efficiently for gain (P = 0.030) than those that were fed the HE or MT during the entire 60-day period. However, the difference between MT and HE was not significant (P = 0.336) in this period. Final weights of sheep fed the HP diet were 26% and 9% greater (P = 0.039) than those fed the HE or MT, respectively.
Carcass weights and proportions of visceral organs relative to slaughter weight of sheep fed the MT diet and those re-alimented with HE or HP are shown in Table 3. Warm carcass weight was heavier (P = 0.039) for sheep fed HP than for those fed HE and MT. Re-alimentation had no effect (P = 0.129 or less) on the weights of most viscera. However, notable exceptions included the lungs, intestines (small and large) and heart; weights of lungs, expressed as a proportion of slaughter weights were greater (P = 0.051) for sheep re-alimented with the HE and HP than for those fed the MT. The hearts of sheep fed the MT diet were 1.1% of slaughter weight compared with only 0.6% and 0.7% for those re-alimented with HE and HP, respectively. Protein re-alimentation also increased (P = 0.001) the growth of intestines (empty) compared with re-alimentation with HE or continuous feeding of the MT.
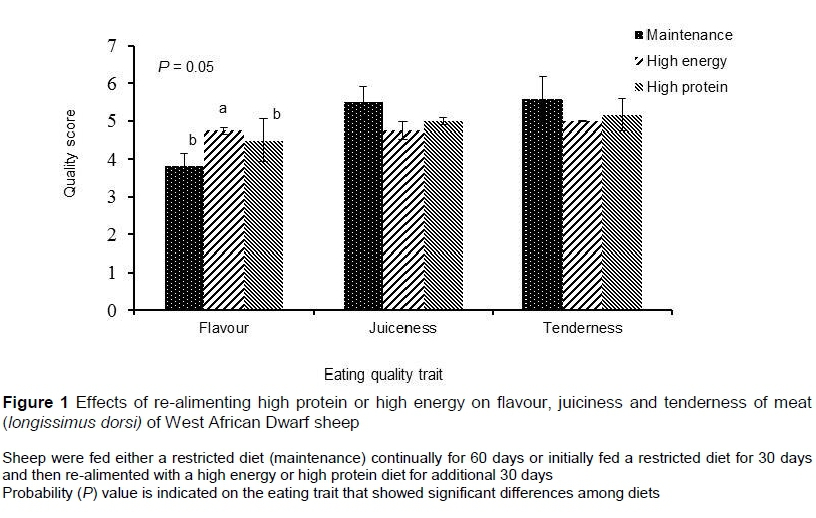
Tenderness and juiciness of meat was similar among sheep fed the various diets. However, the intensity of flavour was slightly stronger (P = 0.05) for sheep re-alimented with HE than for those re-alimented with HP or MT (Figure 1).
Discussion
Sheep on the MT feeding regimen were fed to allow for body maintenance, whereas those in the HP and HE groups were fed to allow for rapid to moderate growth at ad libitum intake. The slower rate of growth observed during the restriction period compared with the re-alimentation period was therefore expected because the sheep were fed only to the level of body maintenance. Upon re-alimentation, however, the ADG for sheep fed the HP increased twofold compared with those fed the HE. Drouillard et al. (1991) found that during the first two weeks of re-alimentation, ADG of sheep re-alimented with protein was more rapid than ADG of sheep re-alimented with energy. They observed that sheep re-alimented with protein accumulated and regained a large proportion of their body protein, which was lost during the previous restriction, than sheep re-alimented with energy. These data suggest that re-alimentation with protein was more effective at inducing compensatory growth following a period of nutrient restriction than re-alimentation with energy. Increased efficiency of nutrient utilization during re-alimentation appears to be the reason for the higher efficiency of feed utilization during the period of re-alimentation in this and other studies (Kamalzadeh et al., 1997; Keogh et al., 2015) because feed intake was not different during the period of re-alimentation. Efficiency of protein utilization (body protein gained : dietary CP consumed) has been found to be greater for sheep re-alimented with a protein diet than for those re-alimented with an energy diet (Drouillard et al., 1991). In this study, the efficiency of weight gain per feed intake indicated that the sheep on the MT gained only 0.22 kg body weight in the first 30 days of the restricted period compared with 0.69 kg in the last 30 days of the same restricted period. This suggests that although the sudden imposition of nutrient restriction initially reduced the ADG of sheep during the first 30-day period, they were able to adapt to this shock by increasing their efficiency of nutrient absorption and utilization. This resulted in higher ADG in the second 30-day period and in the whole 60-day period. This supposition is evidenced because the animals in the MT regimen were fed the same restricted diet throughout the experiment, yet ADG was slightly greater during the second half of the study than the first. This assertion is corroborated by a previous study, which suggested that in arid and semi-arid zones, where dry periods can last for months and feed availability is limited, ruminants have adaptation capacity to withstand moderate malnutrition for medium and long-term duration (Mora et al., 1996).
Despite copious literature on compensatory growth, the mechanisms and cues underpinning the phenomenon are not well elucidated. Some studies (Yambayamba et al., 1996; Keogh et al., 2015) have indicated that lower requirements for body maintenance during re-alimentation following a period of restriction, coupled with sustained capacity for protein synthesis derived from the period restriction, was responsible for the phenomenon. However, Drouillard et al. (1991) concluded that metabolic activity of visceral tissues was not reduced during re-alimentation and reduced metabolic activity alone could not be the cause of the phenomenon of compensatory growth in previously restricted lambs. More recently, Keogh et al. (2016) demonstrated that although increased cell proliferation and rapid growth in animals undergoing compensatory growth were associated with the recovery of liver weight from the period of restriction to the period of re-alimentation, at the end of the re-alimentation period, liver weights were not different between continuously fed animals and the restricted-re-alimented group. They therefore suggested that recovery of liver function, rather than liver weight during compensatory growth, should provide better cues for understanding the molecular basis of compensatory growth. The inconsistencies in these reports suggest that a complex interaction of many factors, including endocrine processes, may be involved in the phenomenon of compensatory growth. These factors may play a role in the higher ADG and efficiency of feed utilization associated with compensatory growth during the re-alimented period. Hornick et al. (2000) suggested that secretions of insulin and growth hormones increase during re-alimentation, thereby allowing more nutrients to be used for growth processes. Hormonal analysis was not done in this study, however. Measurements of the concentrations of insulin, growth hormone and thyroxin during nutrient restriction and re-alimentation may provide useful insights into the interplay between nutrition and endocrinology on compensatory growth.
Feeding sheep throughout the year is a major challenge to rural and urban sheep farmers and traders in sub-Saharan Africa. To ameliorate this challenge, sheep farmers and traders often keep their sheep on a maintenance diet during feed shortages and lower market demand for sheep (Ayantunde et al., 2008). However, as the Islamic festival of Ramadan and other religious festivals draw nearer, they re-aliment them with nutrient-dense supplementary feeds to catch up weight lost during feed scarcity, before selling them. Abegaz et al. (1996) reported that sheep that lost 10% of their weight during restricted feeding in the dry season were able to achieve the same weight as those that were offered supplementary feed throughout the season by growing at a faster rate during re-alimentation. In the present experiment, the improvements in ADG of sheep fed the HP compared with the other diets could be attributed to greater efficiency of weight gain per feed intake rather than to increased DMI because DMI did not differ among sheep during this period. This is consistent with data reported by Drouillard et al. (1991), Abouheif et al. (2015) and Keogh et al. (2015), who found that improvements during compensatory growth were attributed to greater efficiency of feed utilization during re-alimentation, but contradicts observations by Ryan et al. (1993b), who indicated that the phenomenon of compensatory growth in sheep and cattle was attributable to greater feed intake during re-alimentation.
There are doubts about the economic benefits of compensatory growth, because observed increases in live weight gain have been attributed to accumulation of water in the gut (Drew & Reid, 1975) and fat in the muscle (Hornick et al., 2000; Turgeon et al., 1986). Similarly, restoration of ad libitum intake after 20-28% restricted intake increased the compensatory growth rate of sheep. However, this was associated with a concomitant increase in gut content and a reduction in carcass weight and dressing percentage (Winter et al., 1976 ). The results of the present study, however, differed slightly from these previous reports (Drew & Reid, 1975; Winter et al., 1976; Turgeon et al., 1986; Hornick et al., 2000) because carcass weights were improved by re-alimentation with the HP diet. Literature suggests that the deposition of protein and fats is greater both in visceral and non-visceral components of the empty body of sheep following re-alimentation with protein than with energy (Drouillard et al., 1991; Keogh et al., 2016). Addah et al. (2016) observed that re-alimentation with protein stimulated compensatory growth more than re-alimentation with energy but growth rates were similar between pigs fed the restricted diet and the protein diet. In pigs, compensatory responses do not appear to be dependent on dietary protein concentration when they were fed a protein-restricted diet (14.4% CP) and re- alimented with a higher dietary CP of up to 18% (Critser et al., 1995). More data on the phenomenon of compensatory growth between ruminants and non-ruminants are needed to elucidate the differences in the responses of these two groups.
Re-alimentation of sheep with the HP diet resulted in 3 kg more carcass than re-alimentation with HE, but the weight of visceral fat as a proportion of slaughter weight was not different among sheep in the three dietary regimens. This study therefore suggest that when sheep are re-alimented with protein instead of energy, the gain in bodyweight does not emanate from increment in gastrointestinal content, as previously thought (Drew & Reid, 1975). Rather such improvements are the result of initial lean tissue accretion during the early period of re-alimentation, which would last for only few weeks, after which increased feed intake would lead to fat deposition (Hornick et al., 2000; Keogh et al., 2016). Indeed Marais et al. (1991) reported increased protein deposition in Dorper sheep following a restricted-re-alimentation feeding. Consequently, compensatory growth may occur in two stages during re-alimentation; the first stage involves greater deposition of protein during the earlier stages of re-feeding, whereas a greater proportion of fat is deposited during the late period of re-feeding (Turgeon et al.,1986; Hornick et al., 2000). This experiment was designed with 30 days restricted feeding and 30 days re-alimentation. Hence the short duration in each period probably did not allow for increased fat deposition in the latter part of the re-alimentation period. The non-responsiveness of most visceral organs to either HE or HP re-alimentation was consistent with earlier literature (Owens et al., 1993; Kamalzadeh et al., 1998), suggesting a greater priority for use of available nutrients by highly metabolic and early maturing body components such as visceral organs, despite the nutritional status of the animal. The growth of most visceral organs is less responsive to nutrient alterations than late-maturing body components such as pelvis and intramuscular fat (Kamalzadeh et al., 1998; Owens et al., 1993). The weights of intestines, heart, lung and liver expressed as a proportion of slaughter weight in this study are comparable with those reported by Kamalzadeh et al. (1998) for Texel-Flemish crossbred sheep after a period of ad libitum intake restrictions. Although the difference between the weights of the heart for MT and the re-alimented group (HE and HP) was small, it was still worth noting because the present finding is contradictory to the data reported in the literature (Kamalzadeh et al., 1998; Keogh et al., 2015) and is therefore difficult to explain. As an essential body organ, nutrient alterations are expected to have minimal effects on the growth of the heart. Nonetheless, cardiac tissue is distinct from tissues of other visceral organs and the difference in weight may be related to differential response of the smooth involuntary tissue of heart to nutrient deficiency compared with the other visceral organs.
Meat from sheep re-alimented with HE had a more intense 'sheepy' flavour than that of sheep fed the HP (Figure 1). The characteristic flavour of meat of any species is influenced greatly by genetic differences in fatty acid metabolism and composition. However, dietary modifications, albeit small, can increase the intensity of this flavour within species. Flavour intensity of meat is related to the type of fat deposited in the meat, which is influenced by dietary nutrition (Melton, 1983; Wood et al., 1999). In sheep, the concentration of methyl-branched-chain fatty acids was found to be higher when they were fed a restricted forage diet followed by a high energy diet (Priolo et al., 2001).
Tenderness and juiciness were not affected by re-alimentation of either energy or protein because there is preferential deposition of protein over adipose following a period dietary restriction. Meat tenderness is reported to increase deposition of fat. Indeed tenderness and juiciness are closely related to eating quality traits: the more tender the meat, the more quickly the juices are released by chewing, and hence the more juicy the meat appears (Smith & Carpenter, 1976). It has also been reported that re-alimented sheep not only achieve similar growth rates to unrestricted counterparts, but also have similar meat composition as unrestricted animals (Thornton et al., 1979).
Conclusion
The results from this study suggest that re-alimentation of protein compared with energy was more effective at inducing compensatory growth and improving the carcass weight of sheep. It appears that the mechanisms associated with compensatory growth may be related more to improved efficiency of feed utilization during re-alimentation than to increased feed intake. Most non-carcass and metabolically active viscera however, were non-responsive to restricted-re-alimented feeding regimens. Sheep re-alimented with energy produced meat with a more intense 'sheepy' flavour than those re-alimented with protein or fed a maintenance diet.
Acknowledgements
We are grateful to staff at the Ruminants Section of the Department of Animal Science of the University for Development Studies, Nyankpala Campus for taking care of the sheep. Technical assistance of Mr. Samuel Addy in the laboratory analyses of feed samples is also appreciated.
Authors' Contributions
EKO provided funds and technical support for the project. AW designed the project, collected the data and did the statistical analysis. AA and EKO also wrote and edited the manuscript until it was submitted to the journal to be considered for publication.
Conflict of Interest Declaration
The authors declare that there is no conflict of interest.
References
Abegaz, S., Tiyo, D. & Gizachew, L., 1996. Compensatory growth in Horro lambs of Ethiopia. In: Small ruminant research and development in Africa. Eds: S.H.B. Lebbie. & E. Kagwini. Proc. 3rd Biennial Conference of the African Small Ruminant Research Network, Kampala, Uganda. pp. 209-213. [ Links ]
Abouheif, M., Al-Owaimer, A., Kraidees, M., Metwally, H. & Shafey, T., 2013. Effect of restricted feeding and realimentation on feed performance and carcass characteristics of growing lambs. Rev. Bras. Zootec. 42, 95-101. [ Links ]
Addah, W., 2014. Utilization of natural pasture for cattle production in Ghana: Can ensiling help? Eds: W. Addah, F. Adzitey, & R. Lomo. Proc. of the 32nd Biennial Conference of the Ghana Animal Science Association, Tamale, Ghana. pp. 270-278. [ Links ]
Addah, W., Baah, J., Okine, E.K. & McAllister, T.A. 2012. A third-generation esterase inoculant alters fermentation pattern and improves aerobic stability of barley silage and the efficiency of body weight gain of growing feedlot cattle. J. Anim. Sci. 90, 1541-15452. [ Links ]
Addah, W., Dzewu, R.R.R. & Alenyorege, B., 2016. Effects of dietary restriction followed by high dietary energy or protein on compensatory growth of Ashanti Black χ Large White crossbred weaner pigs. Trop. Anim. Health Prod. 48, 145-150. [ Links ]
Ayantunde, A.A., Fernández-Rivera, S. & Dan-Gomma, A., 2008. Sheep fattening with groundnut haulms and millet bran in the West African Sahel. Rev. Elev. Med. Vet. Pays. Trop. 61, 215-220. [ Links ]
Baah, J., Tuah, A.K, Addah, W. & Tait, R.M., 2012. Small ruminant production characteristics in urban households in Ghana. Livest. Res. Rural Dev. 24 (5) 2012. [ Links ]
Critser, D.J., Miller, P.S. & Lewis, A.J., 1995. The effects of dietary protein concentration on growth and development. Edward Arnold. London. [ Links ]
Drew, K.R. & Reid, J.T., 1975. Compensatory growth in immature sheep. I. The effects of weight loss and re-alimentation on the whole body composition. J. Agric. Sci. (Cambridge). 85, 193. [ Links ]
Drouillard, J.S., Klopfenstein, T.J., Britton, R.A., Bauer, M.L., Gramlich, S.M., Wester, T.J. & Ferrel, C.L., 1991. Growth, body composition, and visceral organ mass and metabolism in lambs during and after metabolizable protein or net energy restrictions. J. Anim. Sci. 69, 3357-3375. [ Links ]
Ellis, N., 1980. The nutrient composition of Sudanese animal feeds. Bulletin No. 1. Northern and Central Sudan. Animal Nutrition Research Laboratory, Kuku, Khartoum North. Appendix 1, p. 19. [ Links ]
Fisher, A.V., Enser, M., Richardson, R.I., Wood, J.D., Nute, G.R., Kurt, E., Sinclair, L.A. & Wilkinson, R.G., 2000. Fatty acid composition and eating quality of lamb types derived from four diverse breed production systems. Meat Sci. 55, 141-147. [ Links ]
Herrera-Saldana, R.E., Huber, J.T. & Poore, M.H., 1990. Dry matter, crude protein, and starch degradability of five cereal grains. J. Dairy Sci. 73, 2386-2393. [ Links ]
Hornick, J.L., Van Eenaeme, C., Gérard, O. & Dufrasne, I., 2000. Mechanisms of reduced and compensatory growth. Domestic Anim. Endocrinol. 19, 121-132. [ Links ]
ICEIR (Institute for Continuing Education and Interdisciplinary Research), 2010. Ethics policy handbook. University for Development Studies. p. 10. [ Links ]
Kamalzadeh, A., Koops, W. J., Van Bruchem, J., Tamminga, S. & Zwart, D., 1998. Feed quality restriction and compensatory growth in growing sheep: development of body organs. Small Rumin. Res. 71-82. [ Links ]
Kamalzadeh, A., Van Bruchem, J., Koops, W.J., Tamminga, S. & Zwart, D., 1997. Feed quality restriction and compensatory growth in growing sheep: Feed intake, digestion, nitrogen balance and modelling changes in feed efficiency. Livest. Prod. Sci. 52, 209-217. [ Links ]
Keogh, K., Kenny, D.A., Cormican, P., Kelly, A.K. & Waters, S.M., 2016. Effect of dietary restriction and subsequent re- alimentation on the transcriptional profile of hepatic tissue in cattle. BMC Genomics 17, 244 [ Links ]
Keogh, K., Waters, S.M., Kelly, A.K. & Kenny, D.A., 2015. Feed restriction and subsequent realimentation in Holstein Friesian bulls: I. Effect on animal performance; muscle, fat, and linear body measurements; and slaughter characteristics. J. Anim. Sci. 93:3578-3589. [ Links ]
Marais, P.G., Van der Merwe, H.J. & Du Toit, J.E.J., 1991. The effect of compensatory growth on feed intake, growth rate, body composition and efficiency of feed utilization in Dorper sheep. South Afri. J. Anim. Sci. 21, 80-88. [ Links ]
Melton, S.L., 1983. Effect of forage feeding on beef flavor. Food Technol. 37, 239-248. [ Links ]
Mora, O., Shimada, A. & Ruiz, F.J. 1996. The effect of the length and severity of feed restriction on weight, carcass measurements and body composition of goats. J. Agric. Sci. 127, 549-553. [ Links ]
NRC, 2007. Nutrient requirements of small ruminants, sheep, goats, cervids, and New World camelids. National Research Council. National Academic Press, Washington DC, USA. [ Links ]
Okali, C. & Sumberg, J.E., 1985. Sheep and goats, men and women: Household relations and small ruminant development in south-west Nigeria. Agric. Syst. 18, 39-59. [ Links ]
Okali, C. & Upton, M., 1984. The market potential for increased small ruminant production in southwest Nigeria. Eds: J.E. Sumberg, & K. Cassaday. Proc. of the workshop on small ruminant production systems in the humid zone of West Africa, Ibadan, Nigeria. pp. 68-74. [ Links ]
Owens, F.N., Dubeski, P. & Hanson, C.F., 1993. Factors that alter the growth and development of ruminants. J. Anim. Sci. 71, 3138-3150. [ Links ]
Porter, M.G., Steen, R.W.J., Kilpatrick, D.J., Gordon, F.J., Mayne, C.S., Poots, R.E., Unsworth, E.F. & Pippard, C.J. 1995. Electrometric titration as a method of predicting the chemical composition and corrected dry matter concentration of silage. Anim. Feed Sci. Technol. 56, 217-230. [ Links ]
Priolo, A., Micol, D. & Agabriel, J., 2001. Effects of grass feeding systems on ruminant meat colour and favour. A review. Anim. Res. 50, 185-200. [ Links ]
Richter, B.E., Jones, B.A., Ezzell, J.L., Porter, N.L., Avdalovic, N. & Pohl, C. 1996. Accelerated solvent extraction: A technique for sample preparation. Anal. Chem. 6:1033-1039. [ Links ]
Ryan, W.J., Williams, I.H. & Moir, R.J., 1993b. Compensatory growth in sheep and cattle. II. Changes in body composition and tissue weights. Aust. J. Agric. Res. 44, 1623-1633. [ Links ]
Ryan, W.J., Williams, I.H., Moir, R.J., 1993a. Compensatory growth in sheep and cattle. I. Growth pattern and feed intake. Aust. J. Agric. Res. 44, 1609-1621. [ Links ]
SARI, 2015. Annual weather report for 2015. Savannah Agriculture Research Institute, Nyankpala, Tolon District, Ghana. 65 pp. [ Links ]
Schaefer, E.E. 1976. ASTM manual on consumer sensory evaluation. American Society for Testing and Materials (ASTM). ASTM special technical publication No. 682. [ Links ]
Smith, G.C. & Carpenter, Z.L. 1976. Eating quality of meat animal products and their fat content. Proc. of a symposium of the US Board on Agriculture and Renewable Resources, Commission on Natural Resources, Food and Nutrition Board, Assembly of Life Sciences and National Research Council,. National Academies Press, Washington, D.C., USA. pp. 147-182. [ Links ]
Thornton, R.F., Hood, R.L., Jones, P.N. & Re, V.M., 1979. Compensatory growth in sheep. Aust. J. Agric. Res. 30, 135-151. [ Links ]
Turgeon, O.A., Brink, D.R., Bartle, S.J., Klopfenstein, T.J. & Ferrell, C.L., 1986. Effects of growth rate and compensatory growth on body composition in lambs. J. Anim. Sci. 63, 770-780. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Winter, W.H., Tulloh, N.M. & Murray, D.M., 1976. The effect of compensatory growth in sheep on empty body weight, carcass weight and the weights of some offals. J. Agric. Sci. 87, 433-441. [ Links ]
Wood, J.D., Enser, M., Fisher, A.V., Nute, G.R., Richardson, R.I. & Sheard, P.R., 1999. Manipulating meat quality and composition. Proc. Nutr. Soc. 58, 363-370. [ Links ]
Yambayamba, E.S.K., Price M.A. & Foxcroft, G.R., 1996. Hormonal status, metabolic changes and resting metabolic rate in beef heifers undergoing compensatory growth. J. Anim. Sci. 74, 57-69. [ Links ]
Received 5 October 2016
Accepted 12 April 2017
First published online 3 May 2017
# Corresponding author: addweseh@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}