










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.47 no.3 Pretoria 2017
http://dx.doi.org/10.4314/sajas.v47i3.12
ARTICLES
Genetic parameters for test-day milk yield in tropical Holstein Friesian cattle fitting a multiple-lactation random regression animal model
S. MeseretI, #; E. NegussieII
IWolaita Sodo University, College of Agriculture, P.O. Box 138, Wolaita Sodo, Ethiopia
IIBiometrical Genetics, Natural Resources Institute (LUKE), 31600 Jokioinen, Finland
ABSTRACT
Accurate estimates of genetic parameters are essential for genetic improvement of milk yield in dairy cattle and for setting up breeding programmes. Estimates of genetic parameters from test-day models, particularly for Holstein Friesian cattle maintained in tropical environments, are scant in the literature. The objective of this study was therefore to estimate genetic parameters for milk yield by fitting a multiple-lactation random regression animal model (RRM) based on data from Ethiopian Holstein Friesian herds. Data were used from the first three lactations of cows that calved between 1997 and 2013. The data comprised 13 421 test-day milk yield records from 800 cows from two large dairy herds. Variance components were estimated using the average information restricted maximum likelihood method fitting an RRM. Heritability estimates for first, second, and third lactations ranged from 0.20 to 0.26, 0.15 to 0.27, and 0.17 to 0.28, respectively. Heritability estimates ranging from 0.15 to 0.28 indicate that effective genetic improvement should be accompanied by a corresponding improvement of the production environment. Across-lactation genetic correlations between first and second, second and third, and first and third lactations, expressed on a 305-day yield basis, were 0.88, 0.83, and 0.70, respectively. These genetic correlations, less than or equal to 0.88, indicate that different lactations are different traits. For an accurate evaluation of the genetic merit of animals for milk yield, lactations should be treated as different, but correlated traits in a multiple-lactation analysis.
Keywords: Genetic correlation, heritability, Legendre polynomial, test-day model
Introduction
Multiple observations scored on a cow during lactation form a classical example of longitudinal data (Diggle et al., 1994). The current method for genetic evaluation uses several daily measurements, usually taken once a month (test-day) on an individual cow repeatedly over the course of lactation (Ferreira et al., 2002; Dzomba et al., 2010). Use of test-day data would offer a practical solution in developing countries, where there is a lack of infrastructure and resources for daily milk recording throughout lactation (Ilatsia et al., 2007; Gebreyohannes et al., 2012). Test-day records result in fewer measurements of milk yield as opposed to daily recording, and thus reduce the cost of milk recording compromising the accuracy of animal evaluation. This is partly because the correlation between test-day measurements of milk yield decreases as time between the measurements increases (Bilal & Khan, 2009). It also allows greater flexibility in milk recording programmes (Schaeffer et al., 2000).
A random regression test-day model (TDM) is the method of choice to improve the quality of animal evaluation for longitudinal traits, and has been implemented in many countries. In particular, multiple-lactation TDM fitting random regression is used for genetic evaluation of production traits in dairy cattle in various countries. The analysis of multiple-lactation data has shown that genetic correlations among lactations are less than unity (Zumbach et al., 2008), indicating that each lactation performance is more of a separate trait than has been appreciated (Powell & Norman, 1981). Furthermore, records of second and later lactations provide more complete information on lifetime performance than those from first lactation only (Powell & Norman, 1981). However, results from the development and application of multi-trait test-day animal evaluations and the necessary estimates of genetic parameters for the various lactations of Holstein Friesian cattle in tropical environments are particularly scanty in scientific literature. A first step towards the possible application of the multiple-lactation TDM in Ethiopia is to estimate the genetic parameters of test- day milk yields records under Ethiopian conditions. Therefore, the objectives of this study were to estimate covariance components and genetic parameters for milk yield fitting a random regression test-day animal model using data on the first three lactations of Holstein Friesian cattle from Ethiopian herds.
Materials and Methods
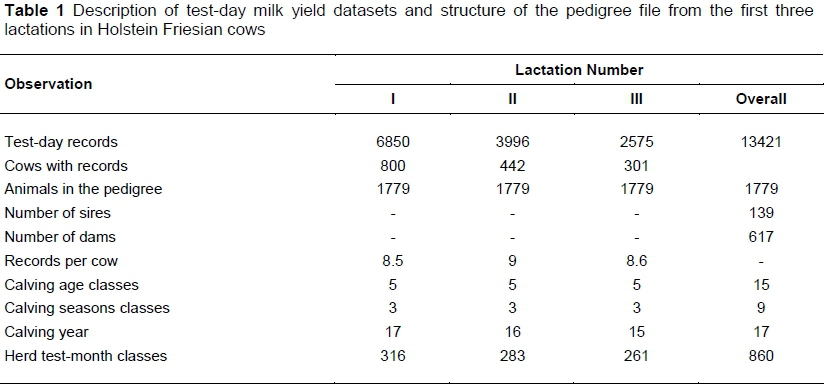
For this study, test-day data were used for the first three lactations of Holstein Friesian cows that calved between 1997 and 2013. They comprised 13 421 test-day milk yield records between 5 and 305 days in milk (DIM) from 800 cows of two herds. Cows had to have test-day records for the first lactation to be included in the dataset. The number of test-day records per lactation ranged from 8 to 10. Five age classes were defined in each lactation. These included cows less than 810, 811 to 990, 991 to 1170, 1171 to 1350 and above 1350 days old at first calving; cows less than 1350, 1351 to 1530, 1531 to 1770, 1771 to 1980 and above 1980 days old at second calving; and cows less than 1800, 1801 to 2040, 2041 to 2280, 2281 to 2520 and above 2520 days at their third calving. Calving months were assigned to one of three seasons, namely long-dry, short-rainy and long-dairy seasons, corresponding with October to February, March to May and June to September, respectively.
The multiple-lactation RRM included the first three lactations as different traits. The statistical model used for the analysis of the data can be described as:
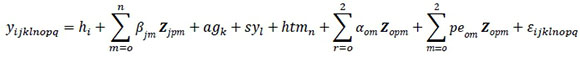
Where: yijkln0pq is the test-day record q of cow o made at DIMop of lactation
hiis the fixed effect of herd I
agkis the fixed effect of age at calving class k
sylis the random effect of sire *calving year interaction l
htmnis the random effects of herd test-day month n
αo is random additive genetic effects
αo is random permanent environmental effects
εijklmnop is the random residual effect
ßjm, α om, and peomare regressions of phenotype on DIM for the f season of calving, and the oth cow's random additive genetic and permanent environmental effects, respectively
Zjpmare the corresponding mth coefficients that describe the shape of the fixed regression lactation curve. Zopmare the corresponding second order Legendre polynomial coefficients that describe the shape of the random regression lactation curves. The fixed regression lactation curve was modelled by a combination of Legendre polynomial and Wilmink functions as co-variables:

Where: Zjp0 ,Zp1 , Zpj2 , Zjp3 and Zpj4 represent coefficients of the third-order orthogonal Legendre polynomial at DIM d plus the Wilmink function (Wilmink, 1987), respectively. In the Wilmink function, the exponential term is generally regarded as a constant (Wilmink, 1987). In the present study, it was assumed to be -0.05. Legendre polynomials in combination with the Wilmink function have been used by various authors (Lidauer et al., 2003; de Melo et al., 2007; Negussie et al., 2008; Santos et al., 2013) in modeling the fixed lactation curves for test-day milk yield in various breeds of dairy cows.
Variance components were estimated by average information restricted maximum likelihood method using the DMU programme, a package for analysing multivariate mixed models (Madsen & Jensen, 2013). Convergence of solutions was determined when the difference between the right-hand side and left-hand side of the mixed model equation (MME) was less than 10-6. The resulting estimates of random regression coefficients were then used to calculate variance components and associated genetic and phenotypic parameters.
For each lactation, additive genetic variance  for DIM dtwas estimated as:
for DIM dtwas estimated as:
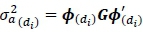
Where G is the covariance matrix of the additive genetic random regression coefficients, and di is DIM.
Similarly, the permanent environmental variance  for DIM dtwas estimated as:
for DIM dtwas estimated as:
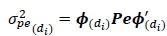
Where: Pe is the covariance matrix of the permanent environment random regression coefficients, and di is DIM.
Heritability for a particular DIM dtin lactation were calculated by dividing the genetic variance  by the sum of permanent environmental
by the sum of permanent environmental  , genetic
, genetic  and residual
and residual  variances for a particular DIM di.
variances for a particular DIM di.
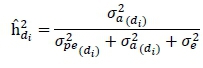
The genetic correlation between two days in lactation diand dj was calculated by dividing the additive genetic covariance between days diand dj by the product of the square root of the genetic variances of days di and dj.
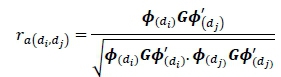
Similarly, the phenotypic correlation was calculated by dividing the phenotypic covariance between days diand dj, by the product of square root of phenotypic variances of day diand dj.
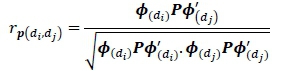
Where: P is the covariance matrix of the phenotypic regression coefficients
The genetic and phenotypic correlations between any two lactations, for example 1 and 2 for the same DIM from multiple-lactation RRM analyses, were calculated by dividing the additive genetic covariance between days diland di2by the product of the square root of the genetic variances of the days diland di2.
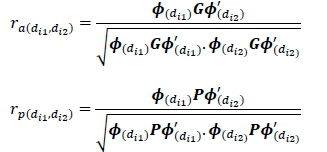
Where: di1 and di2 are DIM in lactation 1 and 2, respectively. G and P are the covariance matrix of the additive genetic and phenotypic regression coefficients from the multiple-lactation analysis, respectively. Genetic and phenotypic correlations on a 305-day basis were calculated as follows:
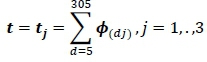
Where: t is a row vector of order quadratic, with elements equal to the sum over days 5 to 305.
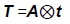
Where: A is 3-by-3 identity matrix.
The genetic correlations between any two lactations, l1and l2for example, between lactation 1 and 2, for 305-day were calculated by dividing the additive genetic covariance between 305-day l1and l2by the product of the square root of the genetic variances of the 305-day l1and l2. The same steps were followed to calculate phenotypic correlations.
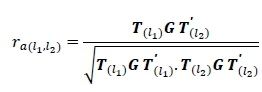
Results
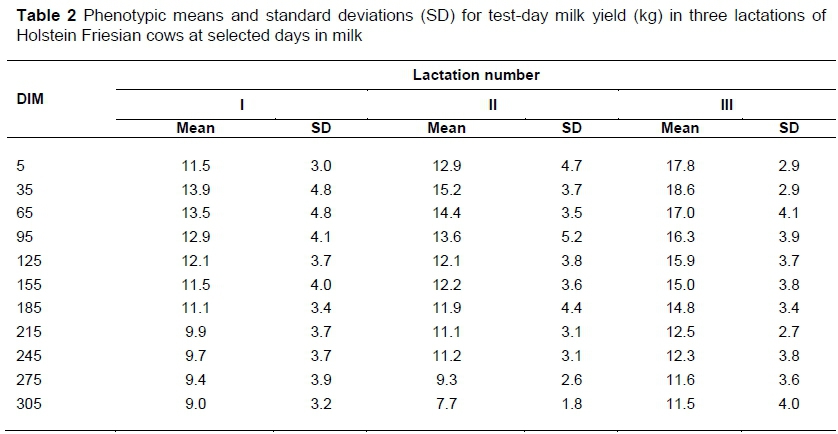
The structure of the dataset and pedigree file are indicated in Table 1. The attrition in number of cows from the first to third calving is consistent with the structure of Holstein herds in Ethiopia, where cows attain, on average, 2.6 lactations (Table 1). The overall means and standard deviations for test-day milk yields for first, second, and third lactations were 11.1 ± 3.9, 13.0 ± 4.7, and 13.9 ± 5.1 kg, respectively. The phenotypic means and standard deviation for test-day milk yield along different DIM in the first three lactations are indicated in Table 2. The phenotypic trend for test-day milk yield for all lactations showed a lower milk yield at the beginning of lactation with peaks at 30 to 35 DIM. After peak lactation, test-day milk yield showed a gradual but consistent decline until the end of the lactation period.
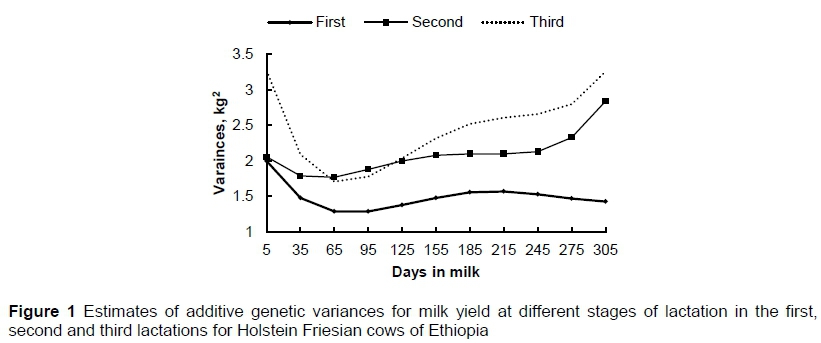
The residual variances estimated from multiple-lactation analysis fitting a multi-trait RRM model were 2.33, 2.75 and 3.06 kg2 for first, second and third lactations, respectively. In Figures 1 and 2, estimates of additive genetic and permanent environmental variances, respectively, are shown for the test-day milk yield at various stages during the first three lactations. The estimated additive genetic variances from first, second and third lactations ranged from 1.29 to 2.00, 1.77 to 2.84 and 1.71 to 3.27 kg2, respectively. The estimated variance of sire by calving year interaction from first, second, and third lactations was 0.09, 0.22, and 0.47 kg2, respectively, and accounts for a small portion of the total variance. In all lactations, the additive genetic variances decreased at the beginning of the lactation trajectory and increased gradually towards the end. In particular, in the second and third lactations, higher variances were observed at the extreme ends of the lactation period. On the other hand, in the first lactation, a gentle decline in the estimates of additive genetic variances was observed towards the end of the lactation. The highest additive genetic variances for first lactation were at the beginning of lactation and the overall pattern of the variances along the lactation trajectory was somewhat consistent after early lactation. In general, the results showed that estimated additive genetic variances for first lactation were lower than those of later lactations. The third lactation had the highest estimated additive genetic variances compared with of the first and second lactations.
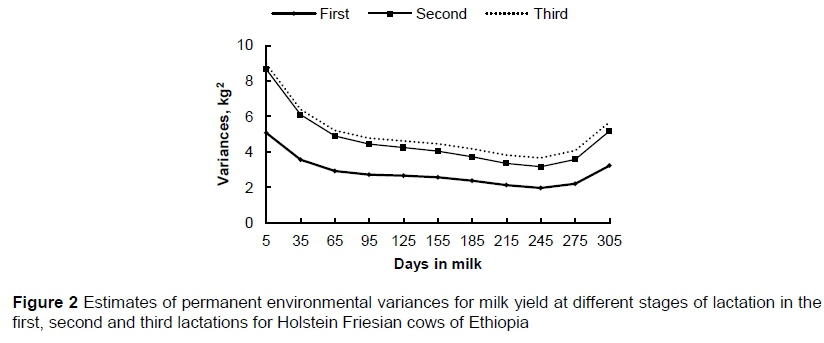
During lactation, the estimated permanent environmental variances ranged from 1.98 to 5.08, 3.17 to 8.66, and 3.67 to 8.95 kg2 in the first, second, and third lactations, respectively. The permanent environmental variances for milk yield were higher than the corresponding additive genetic variances in all three lactations. Similar to the additive genetic variances, the permanent environmental variances showed a marked decline at the beginning of the lactation, were lower in mid lactation, and increased slightly towards the end of lactation in all three parities. Furthermore, the estimated permanent environmental variances showed an increase with increase in lactation number, as did the additive variances.
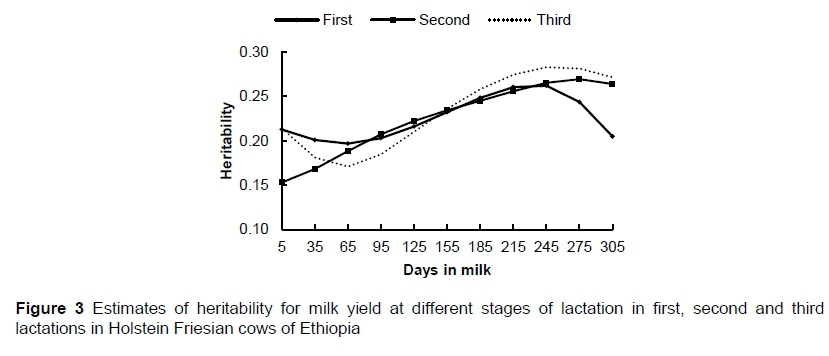
Heritability estimates for test-day milk yield from multiple-lactation RRM analysis are presented in Figure 3. Overall, the estimated heritabilities for test-day milk yield ranged from 0.15 to 0.28 across lactation in the first three lactations. Estimated heritabilities for the first, second, and third lactations ranged from 0.20 to 0.26, 0.15 to 0.27, and 0.17 to 0.28, respectively. In general, the magnitude of the heritability estimates was higher between DIM 230 and 260 compared with the beginning and end (extremes) of lactation for all parities. The difference in heritability estimates between first and second lactations was small for most of the lactation, except at the extremes. The estimates of heritability for the third lactation showed a marked decline in the early stages of lactation, with a sharp increase from DIM 65 to DIM 245, followed by a slight decline thereafter.
Genetic correlations among test-day milk yields at the same DIM in the first three lactations are presented in Table 3. The genetic and phenotypic correlations between lactations ranged from 0.37 to 0.90 and 0.17 to 0.52, respectively, for the different stages of lactation. Genetic correlations between adjacent lactations such as first and second or second and third were higher than those between first and third lactations. Similarly, phenotypic correlations among test-day milk yields at the same DIM in the three lactations were all positive and greater between consecutive lactations than between the first and third lactations. The estimates of genetic correlations calculated on 305-days basis between first and second, first and third and second and third lactations were 0.88, 0.70 and 0.83, respectively. In addition, the estimates of phenotypic correlations on 305-days basis between first and second, first and third and second and third lactations were 0.56, 0.51 and 0.61, respectively.
Discussion
In dairy cows, the level of test-day milk yield differed noticeably between lactations. The results of this study in general showed that the overall mean test-day milk yield in first lactation cows was lower than that of cows in later lactations. Previous studies by Zavadilová et al. f2005) and Tijani et al. (2010) also reported that the overall means for milk yield increased with lactation number. The lower milk production during the first compared with later lactations could be explained because first lactation cows were still growing, and nutrients are therefore partitioned between growth and milk production.
In the present study, the overall means of test-day milk yield for the first three lactations were lower than those reported by Zavadilová et al. (2005) for Holstein cows (18.3, 21.8, and 23 kg for first, second, and third lactations, respectively) in Czech and Tijani et al. (2010) for Holstein cows (18.4, 19.8, and 20.9 kg for first, second, and third lactations, respectively) in Morocco. Similarly, Costa et al. (2008) and Bignardi et al. (2011), working on Brazil Holstein, cattle reported higher first lactation test-day milk yield of 22.53 and 27.45 kg, respectively, compared with the estimates from the current study.
A similar phenotypic trend to this study was reported for first lactation test-day milk yield of Holstein cattle by Shadparvar & Yazdanshenas (2005), Abdullahpour et al. (2010) and Boujenane & Hila (2012). An interesting observation is that in the Ethiopian Holstein Friesian, the peak of lactation was attained at DIM 30, which was earlier compared with that of other populations. For example, for Holstein cows in Brazil (Santos et al., 2013) and Sahiwal cows in India (Dongre & Gandhi, 2014) the peak of lactation was attained at around the eighth and seventh weeks of lactation, respectively.
The goal of selection in dairy cattle is to improve lifetime production of cows, which implies that taking the different lactations into account (Beaumont, 1989) is essential for the estimation of genetic parameters and animal evaluations. In most tropical countries, genetic evaluation of cattle includes different lactations as repeated measures of the same trait, therefore assuming that different lactations are influenced by the same set of genes (Gebreyohannes, 2012). Results from this study indicate that lactations should be regarded as different but correlated traits in genetic evaluations. In this study, residual variance showed an increasing trend with lactation number. A steady rise in residual variance with increase in lactation number was reported by Tong et al. (1979), Druet et al. (2005) and Zavadilová et al. (2005) in dairy cattle and Zumbach et al. (2008) in dairy goats. This study confirms that heterogeneity in additive genetic and permanent environmental variances exists over lactations and across DIM within lactation. Several authors, including Tong et al. (1979), Guo et al. (2002) and Druet et al. (2005), also reported that in dairy cattle variances increased with lactation number. Higher additive genetic variances observed at the extreme ends of the second and third lactations in this study are in line with those of Zavadilová et al. (2005), who reported high genetic variances at the beginning and end of the first three lactations in Czech Holstein. On the contrary, Rekaya et al. (1999) in Spanish Holstein Friesian, Pool et al. (2000) in Netherlands Holstein Friesian and Druet et al. (2003) in French Holstein showed the highest genetic variance in mid lactation and lower estimates at the beginning and end of lactation.
In this study, sire by calving year interaction was included in the random parts of the models mainly to account for the various groups of sires used across different year groups, which improved the variance structure and led to fast convergence of solutions. The genetic variance for milk yield is one indicator for the achievement of genetic improvement programmes in the country. Several studies reported varying trends for additive genetic variances across the lactation period. For instance, Zavadilová et al. (2005) in Czech Holstein, Caccamo et al. (2008) in Italian dairy cows, and Mohammadi et al. (2014) in Iranian Holstein reported higher estimates of additive genetic variances at the beginning and towards the end of the lactation. On the other hand, El Faro et al. (2008) in native Brazilian Caracu heifers reported higher additive genetic variances at the beginning and a declining trend afterwards. Flores & van der Werf (2015) from their work on data from a buffalo population reported higher estimates of additive genetic variance in early and during peak lactation. The higher permanent environmental variance than that of additive genetic variance, observed in this study, could be the limited data used for variance component analysis.
Except at the extremes of the lactation periods, the trend for the estimated heritability was increasing across DIM in all parities. Similar increases in the course of the heritability curve in three lactations were reported by Guo et al. (2002) in Danish Jersey and Zavadilová et al. (2005) in Czech Holstein. The heritabilities among lactations were only slightly different from mid to end of lactation. The heritabilities from third lactation were higher than those of the first and second lactations for this period, while no significant differences in heritability estimates were observed between the first and second lactations. Similar findings have been reported from black and white cattle (Strabel & Misztal, 1999) in Poland and Holstein cows (Zavadilová et al., 2005) in Czech, indicating that heritability increased with increase in parity number. On the contrary, other studies reported that estimates of heritability for consecutive lactations decreased with increase in lactation number in Holstein cows (Liu et al., 2000) in Germany and Jersey cows (Guo et al., 2002) in Danish.
One of the key points in dairy breeding programmes is selection of young bulls, usually based on their daughters' performance in first lactation. Because longevity, and hence performance in later lactations, is also of interest to breeders, the effectiveness of selection schemes depends on the genetic correlations between the first and later lactations (Strabel & Jamrozik, 2006). If genetic correlations between different lactations are less than unity, this implies that different set of genes influence test-day milk yields in different lactations. In this study, the genetic and phenotypic correlations between test-day records depend on the stage of lactation and parity number. The genetic and phenotypic correlations calculated, based on both daily and an accumulated 305-day basis between lactations were higher for adjacent lactations (first and second, and second and third) than those estimated between first and third lactations. Similarly, several authors have reported that genetic correlations of test-day milk yield between the first and second lactations were higher than the correlations between the first and third lactations Czech Holstein cows (Zavadilová et al., 2005), Polish black and white cattle (Strabel & Jamrozik, 2006) and Moroccan Holstein cows (Tijani et al., 2010). In this study, the genetic correlations between lactations were slightly higher than those of other studies. For example, Strabel et al. (2004) in Polish black and white cattle population reported that genetic correlations between adjacent lactations were 0.70 for milk yield, whereas between first and third lactations they were lower (around 0.62). Similarly, Zavadilová et al. (2005) in Czech Holstein cows showed genetic correlations of 0.77, 0.75 and 0.61 between first and second, second and third, and first and third lactations, respectively. In this study, the patterns of genetic correlations showed an increase along DIM and a slight decline towards the end of the lactations. However, Strabel & Jamrozik (2006) in Polish black and white cattle and Holstein cows Tijani et al. (2010) in Moroccan Holstein cows reported genetic correlations across lactations with smallest estimates at the extremes of lactation and highest in the middle of lactation.
Conclusion
Estimates of genetic parameters are essential for genetic improvement of milk yield in dairy cattle and for setting up sustainable dairy breeding programmes. Results from this study indicate that heritability of milk production ranged from 0.15 to 0.30. This suggests that although there is potential for genetic improvement in milk yield of tropical Holstein Friesian, simultaneous improvement of the production environment is essential. Furthermore, results from the multiple-lactation RRM analysis showed that correlations among the first three lactations on 305-d basis ranged from 0.70 to 0.88. This indicates that for accurate evaluation of the genetic merit of animals, the various lactations should be treated as different, but correlated traits in a multiple-lactation analysis.
Acknowledgments
The authors would like to acknowledge the Ethiopian Ministry of Agriculture, National Artificial Insemination Centre for providing the data. The authors are also grateful to the Biometrical Genetics, Natural Resources Institute, Finland, for their support in the data analyses. Addis Ababa University and Wolaita Sodo University are also acknowledged for their financial support.
Authors' Contributions
SM wrote the manuscript, analysed the data, and interpreted the results. EN participated in the analysis and interpretation of data, drafting and revising the manuscript.
Conflict of Interest Declaration
We wish to state that the financial source and data material used in this study are duly acknowledged and that there are no known conflicts of interest associated with this paper.
References
Abdullahpour, R., Shahrbabak, M.M., Nejati-Javaremi, A. & Torshizi, R.V., 2010. Genetic analysis of dairy milk, fat and protein percentage of Iranian first lactation Holstein cattle. World Appl. Sci. J. 10, 1042-1046. [ Links ]
Beaumont, C., 1989. Restricted maximum likelihood estimation of genetic parameters for first three lactations in the Montbéliarde dairy cattle breed. Genet. Sel. Evol. 21, 493-506. [ Links ]
Bignardi, A.B., El Faro, L., Torres Júnior, R.A.A., Cardoso, V.L., Machado, P.F. & Albuquerque, L.G., 2011. Random regression models using different functions to model test-day milk yield of Brazilian Holstein cows. Genet. Mol. Biol. 10, 3565-3575. [ Links ]
Bilal, G. & Khan, M.S., 2009. Use of test-day milk yield for genetic evaluation in dairy cattle. Pakistan Vet. J. 29, 35-41. [ Links ]
Boujenane, I. & Hila, B., 2012. Genetic and non-genetic effects for lactation curve traits in Holstein-Friesian cows. Archiv. Tierzucht 55, 450-457. [ Links ]
Caccamo, M., Veerkamp, R.F., de Jong, G., Pool, M.H., Petriglieri, R. & Licitra, G., 2008. Variance components for test- day milk, fat, and protein yield, and somatic cell score for analyzing management information. J. Dairy Sci. 91, 3268-3276. [ Links ]
Costa, C.N., de Melo, C.M.R., Packer, I.U., de Freitas, A.F., Teixeira, N.M. & Cobuci, J.A., 2008. Genetic parameters for test day milk yield of first lactation Holstein cows estimated by random regression using Legendre polynomials. R. Bras. Zootec. 37, 602-608. [ Links ]
de Melo, C.M.R., Packer, I.U., Costa, C.N. & Machado, P.F., 2007. Genetic parameters for test day milk yields of first lactation Holstein cows by random regression models. Animal, 1, 325-334 [ Links ]
Diggle, P.J., Liang, K.Y. & Zeger, S.L., 1994. Analysis of longitudinal data. Oxford University Press, New York. [ Links ]
Dongre, V.B. & Gandhi, R.S., 2014. Genetic and phenotypic parameters of fortnightly test day and first lactation 305-day or less milk yield in Sahiwal cattle. Int. J. Livest. Res. 4, 17-20. [ Links ]
Druet, T., Jaffrézic, F. & Ducrocq, V., 2005. Estimation of genetic parameters for test day records of dairy traits in the first three lactations. Genet. Sel. Evol. 37, 257-271. [ Links ]
Druet, T., Jaffrezic, F., Boichard, D. & Ducrocq, V., 2003. Modeling lactation curves and estimation of genetic parameters for first lactation test-day records of French Holstein cows. J. Dairy Sci. 86, 2480-2490. [ Links ]
Dzomba, E.F., Nephawe, K.A., Maiwashe, A.N., Cloete, S.W.P., Chimonyo, M., Banga, C.B., Muller, C.J.C. & Dzama K., 2010. Random regression test-day model for the analysis of dairy cattle production data in South Africa. S. Afr. J. Anim. Sci. 40, 273-284. [ Links ]
El Faro, L., Cardoso, V.L. & de Albuquerque, L.G., 2008. Variance component estimates for test-day milk yield applying random regression models. Genet. Mol. Biol. 31, 665-673. [ Links ]
Ferreira, W.J., Teixeira, N.M., Torres, R.A. & Silva, M.V.G.B., 2002. Utilizao da produo de leite no dia do controlenaavaliaogeneticaemgado de leite - Uma reviso. Arch. Latinoam. de Prod. Anim. 10, 46-53. (in Portuguese) [ Links ]
Flores, E.B. & van der Werf, J., 2015. Random regression test day models to estimate genetic parameters for milk yield and milk components in Philippine dairy buffaloes. J. Anim. Breed. Genet. 132, 289-300. [ Links ]
Gebreyohannes, G., Koonawootrittriron, S., Elzo, M.A. & Suwanasopee, T., 2012. Fitness of lactation curve functions to daily and test-day milk data in an Ethiopian dairy cattle population. Annual meeting abstracts, poster presentation. J. Anim. Sci. 90, Supplement 3,15-20 July 2012. [ Links ]
Guo, Z., Lund, M.S., Madsen, P., Korsgaard, I. & Jensen, J., 2002. Genetic parameter estimation for milk yield over multiple parities and various lengths of lactation in Danish Jerseys by random regression models. J. Dairy Sci. 85, 1596-1606. [ Links ]
Ilatsia, E.D., Muasya, T.K., Muhuyi, W.B. & Kahi, A.K., 2007. Genetic and phenotypic parameters for test day milk yield of Sahiwal cattle in the semi-arid tropics. Animal, 1, 185-192. [ Links ]
Lidauer, M., Mäntysaari, E.A. & Strandén, I., 2003. Comparison of test-day models for genetic evaluation of production traits in dairy cattle. Livest. Prod. Sci. 79, 73-86. [ Links ]
Liu, Z., Reinhardt, F & Reents, R., 2000. Estimating parameters of a random regression test day model for first three lactation milk production traits using the covariance function approach. Interbull Bull. 25, 74-80. [ Links ]
Madsen, P. & Jensen, J., 2013. DMU: A package for analysing multivariate mixed models. Version 6, release 5.2. Center for Quantitative Genetics and Genomics Dept. of Molecular Biology and Genetics, University of Aarhus Research Centre Foulum Box 50, 8830 Tjele Denmark. [ Links ]
Mohammadi, A., Alijani, S. & Daghighkia, H., 2014. Comparison of different polynomial functions in random regression model for milk production traits of Iranian Holstein dairy cattle. Ann. Anim. Sci. 14, 55-68. [ Links ]
Negussie, E., Stranden, I. & Mäntysaari, E.A., 2008. Genetic association of clinical mastitis with test-day somatic cell score and milk yield during first lactation of Finnish Ayrshire cows. J. Dairy Sci. 91, 1189-1197. [ Links ]
Pool, M.H., Janss, L.L.G. & Meuwissen, T.H.E., 2000. Genetic parameters of Legendre polynomials for first-parity lactation curves. J. Dairy Sci. 83, 2640-2649. [ Links ]
Powell, R.L. & Norman, H.D., 1981. Different lactations for estimating genetic merit of dairy cows. J. Dairy Sci. 64, 321-330. [ Links ]
Rekaya, R., Carabano, M.J. & Toro, M.A., 1999. Use of test day yields for the genetic evaluation of production traits in Holstein-Friesian cattle. Livest. Prod. Sci. 57, 203-217. [ Links ]
Santos, D.J.A., Peixoto, M.G., Borquis, R.R., Verneque, R.S., Panetto, J.C. & Tonhati, H., 2013. Comparison of random regression models to estimate genetic parameters for milk production in Guzerat (Bos indicus) cows. Genet. Mol. Biol. 12, 143-153. [ Links ]
Schaeffer, L.R., Jamrozik, J., Kistemaker, G.J. & Van Doormaal, B.J., 2000. Experience with a test-day model. J. Dairy Sci. 83, 1135-1144. [ Links ]
Shadparvar, A.A. & Yazdanshenas, M.S., 2005. Genetic parameters of milk yield and milk fat percentage test day records of Iranian Holstein cows. Asian-Aust. J. Anim. Sci. 18, 1231-1236. [ Links ]
Strabel, T. & Jamrozik, J., 2006. Genetic analysis of milk production traits of Polish Black and White cattle using large- scale random regression test-day models. J. Dairy Sci. 89, 3152-3163. [ Links ]
Strabel T. & Misztal I., 1999. Genetic parameters for first and second lactation milk yields of Polish black and white cattle with random regression test-day models. J. Dairy Sci. 82, 2805-2810. [ Links ]
Strabel, T., Ptak, E., Szyda, J. & Jamrozik J., 2004. Multiple-lactation random regression test-day model for Polish Black and White cattle. Interbull Bull. 32, 133-136. [ Links ]
Tijani, A., Rgayai, B., Hammami, H., Gillon, A. & Gengler, N., 2010. Estimation of genetic parameters for test-day milk yields of Moroccan Holstein cows using a random regression test-day model. Proceedings of the 9th World Congress on Genetics Applied to Livestock Production. German Society for Animal Science. 1-6 August 2010, Leipzig, Germany. [ Links ]
Tong, A.K.W., Kennedy, B.W. & Moxley, J.E., 1979. Heritabilities and genetic correlation for first three-lactations from record subject to culling. J. Dairy Sci. 62, 1784-1790. [ Links ]
Wilmink, J.B.M., 1987. Adjustment of test-day milk, fat, and protein yields for age, season, and stage of lactation. Livest. Prod. Sci. 16, 335-348. [ Links ]
Zavadilová, L., Jamrozik, J. & Schaeffer, L.R., 2005. Genetic parameters for test-day model with random regressions for production traits of Czech Holstein cattle. Czech J. Anim. Sci. 50, 142-154. [ Links ]
Zumbach, B., Tsuruta, S., Misztal, I. & Peters, K.J., 2008. Use of a test day model for dairy goat milk yield across lactations in Germany. J. Anim. Breed. Genet. 125, 160-167. [ Links ]
Received 12 December 2016
Accepted 29 March 2017
First published online 26 April 2017
# Corresponding author: meseretselam@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}