










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.47 n.2 Pretoria 2017
http://dx.doi.org/10.4314/sajas.v47i2.8
ARTICLES
Effect of supplementing or treating Eragrostis curvula hay with urea or nitrate on its digestibility and in vitro fermentation
F.A. Adejoro; A. Hassen#
Department of Animal and Wildlife Sciences, University of Pretoria, Private bag X20, Hatfield 0028, Pretoria. South Africa
ABSTRACT
The potential of dietary nitrate to reduce enteric methane, apart from it being a source of rumen-degradable nitrogen, has stimulated further research into its use. However, not much has been reported on its suitability in feed treatment. The objective of this study was to determine the effects of urea or nitrate and two methods of non-protein nitrogen (NPN) application (anaerobic pre-treatment versus direct supplementation) to Eragrostis curvula hay, on its in vitro fermentation. An iso-nitrogenous level of NPN (7 g nitrogen/kg feed dry matter (DM)) from either urea or calcium nitrate was used to pre-treat hay by subjecting it to 30 days' anaerobic storage in airtight bottles, or by direct supplementation; each diet having three replicates. Hay samples were dried, milled, and evaluated for in vitro organic matter digestibility (IVOMD), and in vitro gas and methane production, while rumen fluid was analysed for pH, ammonia nitrogen and volatile fatty acids (VFAs). Compared with the untreated hay (control), under both methods of application NPN generally did not increase 48-hour gas production, but did increase the IVOMD of E. curvula hay. However, pre-treatment of E. curvula hay improved IVOMD more than supplementation. Urea inclusion enhanced digestibility more than nitrate, and the urea pre-treatment was more effective than supplementation. In contrast, nitrate inclusion significantly reduced methane production compared with urea and the control, and supplementation of nitrate was more effective in reducing methane than pre-treatment with nitrate. Pre-treatment with nitrate as supplementation increased digestibility and reduced enteric methane emission, indicating the potential of using nitrate as a hydrolytic agent in feed treatment.
Keywords: digestibility, feed treatment, methane emission, non-protein nitrogen, volatile fatty acids
Introduction
Roughages such as grass hay, agro-industrial by-products, and crop residues have remained the main feed resources for ruminant feeding in larger parts of tropical and subtropical Africa (Preston & Leng, 1987). These roughages could become significant ingredients, even in the diet of high-producing animals, if their digestibility could be improved (Wanapat et al., 2009). The availability of rumen ammonia is often a primary deficit when protein-deficient feedstuffs such as grass hays are fed to ruminants, limiting microbial biomass production, digestibility, and feed intake (Hao Trinh et al., 2009). Enteric methane emissions are dependent on the amount of feed consumed by the animal and the composition of the diet, with lower emission values from concentrate than from roughage-based diets (Mirzaei-Aghsaghali & Meheri-Sis, 2011; Gemeda & Hassen, 2014a). Methane per unit of organic matter degraded could be up to two times higher in tropical forages than in legumes (Widiawati et al., 2007; Gemeda & Hassen, 2014a). In the past, many studies have been conducted globally on improving the feeding value of roughages (Wanapat et al., 2009; Shreck, 2013; Gemeda et al., 2014b). One such method involves treating feedstuff with aqueous ammonia or urea through the process of ammoniation (Mapato et al., 2010; Diaz et al., 2013; Moreira Filho et al., 2013). Urea has been used as a hydrolytic agent that helps to disrupt the cell wall structure, and break the ester bonds between lignin, cellulose and hemicellulose, thus improving the digestibility of feedstuffs (Vadiveloo, 2003). Urea is also convertible to ammonia in the rumen, thus, contributing to the synthesis of microbial protein (Schroeder & Titgemeyer, 2008; Mapato et al., 2010; Shreck, 2013).
Urea was a popular NPN source of choice in the feeding system of ruminants. More recently, however, there has been renewed interest in the use of nitrate because of its potential benefits as a source of degradable N, and for methane mitigation (Li et al., 2012). Urea could be replaced with nitrate in low rumen degradable protein diets to provide a source of rumen ammonia, while reducing enteric methane production (Le Huyen et al., 2010; Van Zijderveld et al., 2010; Takahashi, 2011). Under anaerobic conditions, nitrate could undergo conversion to ammonia (Leng, 2008). The major limitation to the use of nitrate and other NPN sources in ruminant feeding is their potential to induce methemoglobinemia as a result of elevated ammonia-N and nitrite in the rumen (Van Zijderveld et al., 2010, Lee & Beauchemin, 2014; Newbold et al., 2014). However, the risk of toxicity could be reduced through gradually adjusting the animals to a high nitrate diet with regulated dosing (Nolan et al., 2010; Lee & Beauchemin, 2014; Newbold et al., 2014). Earlier studies have shown that the method of nitrate application has significantly influenced its utilization and toxic dose in animals (Van Zijvderveld et al., 2010). However, the suitability of nitrate in the treatment of poor-quality roughages to improve their utilization and reduce the enteric methane associated with the use of such feeds has not been evaluated extensively in South Africa. Eragrostis curvula hay is a popular feed resource in South Africa, with mean crude protein content usually of less than 60g/kg DM, which may require supplementation or treatment with NPN (Tesfayohannes et al., 2013). This study, therefore, aims to compare the effects of replacing urea with nitrate in two methods of application (as a feed supplement and in pre-treatment of grass hay) on digestibility, in vitro gas and methane production of Eragrostis curvula hay.
Materials and Methods
This study was conducted at the University of Pretoria's Experimental Farm after the approval of the trial protocol by the Animal Ethics Committee of University of Pretoria (No. EC061-14) had been obtained.
Eragrostis curvula hay was pre-treated or directly supplemented with NPN from two sources (urea and nitrate). The amount of urea or nitrate was set at iso-nitrogenous levels to provide 7 g nitrogen/kg hay DM as the supplemented group. For the second or treated group, hays were mixed and subsequently kept under anaerobic conditions for 30 days prior to sampling. An in vitro digestibility and gas production study was conducted to evaluate the response to NPN (urea or nitrate) supplementation or treatment. Urea and calcium nitrate are feed-grade fertilizers, which were obtained from Introlab Chemicals (Pty) Ltd, Pretoria, South Africa.
Eragrostis curvula hay was collected from the feedstock at the University of Pretoria Experimental Farm, chopped to about 5 cm, and used as experimental diets. Feed treatment was carried out according to the procedure described by Tesfayohannes et al. (2013). Urea and calcium nitrate were solubilized in water, and the entire solution was sprayed onto the hay, the final mix having a moisture content of approximately 40%. Grass hay was thoroughly hand-mixed with additives inside a big plastic container and sampled for the in vitro evaluation as urea- or nitrate-supplemented diets. For the second treatment, grass hay was pre-mixed (as for the supplemented group) with urea or nitrate solutions at the same iso-nitrogenous level (7.0 g N/kg DM), filled into glass bottles, which served as mini silos, compressed, and subsequently sealed anaerobically in a vacuum with three replicates per treatment group. Each bottle was stored at room temperature (25 °C) for 30 days. After the 30-day period, the glass bottles were opened, the upper 5 cm were discarded, and contents were emptied into a big container, hand-mixed and then sampled for in vitro evaluation as urea or nitrate-treated diets. All experimental diets were freeze-dried and ground to pass through a 1-mm screen (Wiley mill) before analysis.
Rumen buffer, macro-mineral and micro-mineral solutions for the experiment were prepared as described by Goering & Van Soest (1970) with the modifications of Mould et al. (2005). The prepared buffer solution was kept in a water bath at 40 °C and continuously purged with CO2 until the solution turned colourless. Rumen fluid was collected from two rumen-cannulated merino wethers fed lucerne hay (Medicago sativa) ad libitum, strained through four layers of cheesecloth into a pre-warmed thermos flask, and transported quickly to the laboratory. Rumen fluid was blended in the laboratory, mixed with the buffermineral solution in the ratio of rumen fluid to buffer solution (3 : 5), and added to each serum bottle, which already contained approximately 400 mg of each diet, under a continuous stream of CO2. Each bottle was sealed with a rubber stopper and crimp seal cap. For each incubation run, four blanks, containing only rumen fluid, were used to correct for the effect of rumen fluid. In each incubation run, each test diet had four replicates per run, and three runs were conducted. All bottles were placed in the incubator at 39 °C and 120 rpm. A semi-automated gas pressure system was used to measure gas pressure according to Theodorou et al. (1994). The system consisted of a digital data tracker (Tracker 220 series indicators, Omega Engineering Inc., Laval, QC, Canada), fitted with a pressure transducer (PX4200-015GI, Omega Engineering Inc.) and a needle tip. Gas pressure was taken at 2, 12, 24 and 48 hours after commencement of incubation, and gas samples were taken to analyse the methane concentration. At the end of the incubations, fermentation was terminated by removing the serum bottles from the incubator, and immersing them in an ice bath to impede further microbial activity. Rumen fluid pH was measured after 48 hours' incubation using a pH meter (XLS 15 Accumet® pH meter, Fisher Scientific, Singapore). Supernatant (5 mL) was collected into vials containing 1 mL 0.5M H2SO4 (for ammonia-N analysis) and 1 mL 25% orthophosphoric acid (for volatile fatty acid (VFA) analysis). All vials were frozen at -20 °C until analysis, as described by Broderick & Kang (1980) for NH3-N, and Webb (1994) for VFA.
The IVOMD of diets was determined with the two-phase digestion method of Tilley & Terry (1963), as modified by Engels & Van der Merwe (1967). In the first stage, 200 mg feed samples were incubated in four replicates of each diet with rumen liquor for 48 hours at 39 °C under anaerobic conditions. Blanks and a standard feed were included in each batch of incubation. This was followed by an acid-pepsin digestion phase for 48 hours. After digestion, the residual material was oven-dried at 105 °C for 18 hours, weighed, and subsequently ashed in a muffle furnace at 550 °C for three hours. The IVOMD was estimated from the weights of the starting material and residuals.
Gas samples from the in vitro incubations were taken with a Hamilton syringe at 2, 12, 24 and 48 hours' incubation (Gemeda & Hassen, 2015). Methane concentration was analysed with gas chromatography (8610C BTU Gas Analyser GC System, SRI Instruments Europe GmbH, Bad Honnef, Germany), pre-equipped with a flame ionization detector. Two blanks were included to correct the methane produced from the inoculum in each run, and two runs were executed. Methane concentration values were related to the total gas production to estimate its concentration (Tavendale et al., 2005), and subsequently converted to energy and mass values (Santoso et al., 2007).
The concentrations of acetate, propionate, butyrate, valerate, and isobutyrate were evaluated as molar proportions (mol/100 mol), while total volatile fatty acid (TVFA) was expressed as mmol/L, and ammonia-N concentration was expressed in mg/dL. Total gas and net methane were expressed in mL/g fermented dry matter and as mass values in relation to IVOMD (g/kg IVOMD) and TVFA. The ratio of non-glucogenic to glucogenic VFAs was calculated and expressed as molar ratio of acetate to propionate (A/P molar ratio).
The in vitro gas production experiments were examined in three experimental runs with four replicates (bottles) per treatment combination. In each run, individual bottles served as analytical replicates and were averaged prior to statistical analysis, while each run served as a statistical replicate. Gas volume produced was plotted against incubation time, and the 0rskov & McDonald (1979) non-linear equation was used to estimate gas production characteristics and to calculate effective gas production (EGP). Data were analysed using the GLM procedure of SAS 9.3 (2010). Single degree of freedom orthogonal contrasts were used to test: i) control versus the average of NPN-included diets; ii) average of urea-included diet versus average of nitrate-included diet; iii) in urea subgroup, supplemented versus treated; and iv) in nitrate subgroup, supplemented versus treated on all parameters. For all statistical analyses, significance was declared at P <0.05 and P <0.001, as applicable.
Results
The chemical composition of E. curvula hay used as substrate in this study had the following nutrient content: crude protein (CP) (48.9 g/ kg dry DM), neutral detergent fibre (NDF) (746 g/kg DM), acid detergent fibre (ADF) (447 g/kg DM), acid detergent lignin (ADL) (79 g/kg DM), neutral detergent insoluble nitrogen (NDIN) (21.5 g/kg), and ash (30.4 g/kg).
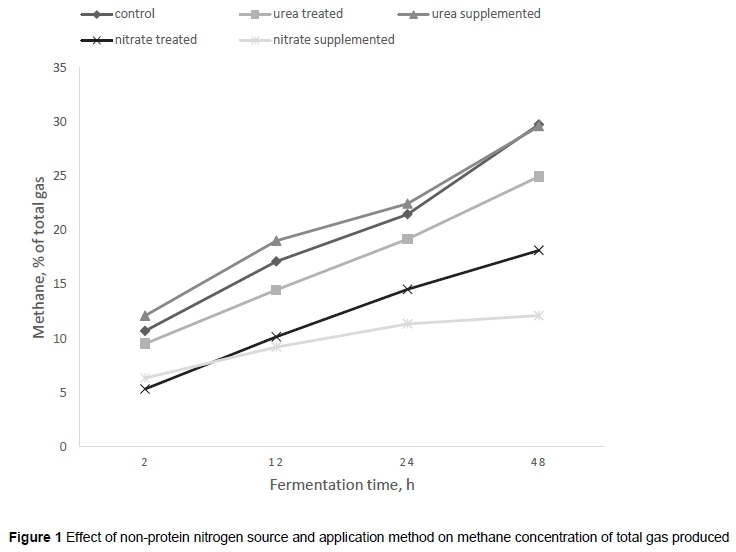
In vitro digestibility and gas production of E. curvula hay supplemented with urea or nitrate and treated with them are summarised in Table 1. Regardless of the source, generally NPN inclusion did not affect total gas production after 48 hours' fermentation compared with the control diet (P >0.05) (Figure 1). Of the two NPN sources, urea generally resulted in higher 48-hour gas production (P <0.05) than nitrate, while the application method did not affect 48-hour gas production in either the urea or the nitrate diet. Generally, NPN inclusion significantly reduced methane production compared with the control diet (P <0.05) (Figure 1). However, of the two NPN sources, nitrate showed a significant (P <0.001) reduction in methane production (by 57.5%) compared with urea, which showed a reduction of 12.5%. The application method (supplemented vs treated) generally did not affect methane production (P >0.05). Although application method did not significantly affect the methane to total gas ratio in either the urea or the nitrate inclusions, there was a significant decrease in methane to total gas ratio with nitrate inclusion compared with urea inclusion and with the control (untreated) diet (P <0.001).
NPN inclusion (urea or nitrate) generally improved (P <0.05) the IVOMD of E. curvula hay, regardless of application method. While the untreated diet recorded 462 g /kg, with the inclusion of urea at 7 g N/kg hay, IVOMD increased to 493 g/kg DM and 502 g/kg DM in urea-supplemented and urea-treated hay, respectively. Similarly, compared with the control (462 g/kg IVOMD), nitrate-supplemented and nitrate-treated diets recorded 482 g/kg and 486 g/kg DM IVOMD, respectively. Urea resulted in significantly higher digestibility compared with nitrate in both the supplementation and the feed treatment. The volume of methane produced per unit of organic matter digested (expressed as mass value) showed that NPN addition significantly (P <0.05) reduced methane (g/kg IVOMD) compared with the control diet, with the average of nitrate-included diets also resulting in significantly lower (P <0.05) methane per unit of organic matter digested compared with urea-included diets. The application method did not result in significant differences in terms of methane per unit of organic matter digested in urea, but the nitrate-supplemented diet recorded lower methane per unit of organic matter digested compared with the nitrate-treated diet (37.9 g/kg versus 58.3 g/kg). Neither NPN source nor application method had any significant effect on TVFA, but the overall effect of NPN inclusion significantly improved TVFA per methane ratio when compared to the control group. However, of the two NPN sources, the nitrate inclusions showed significantly increased TVFA per methane production compared with the urea inclusions. The ratio of total gas production to IVOMD was not significantly different across NPN sources and application methods.
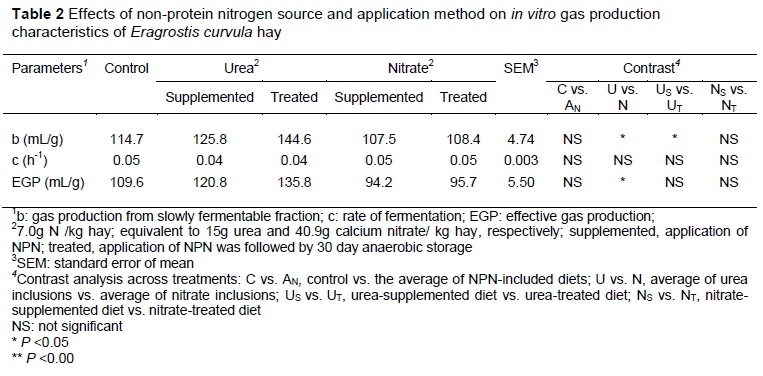
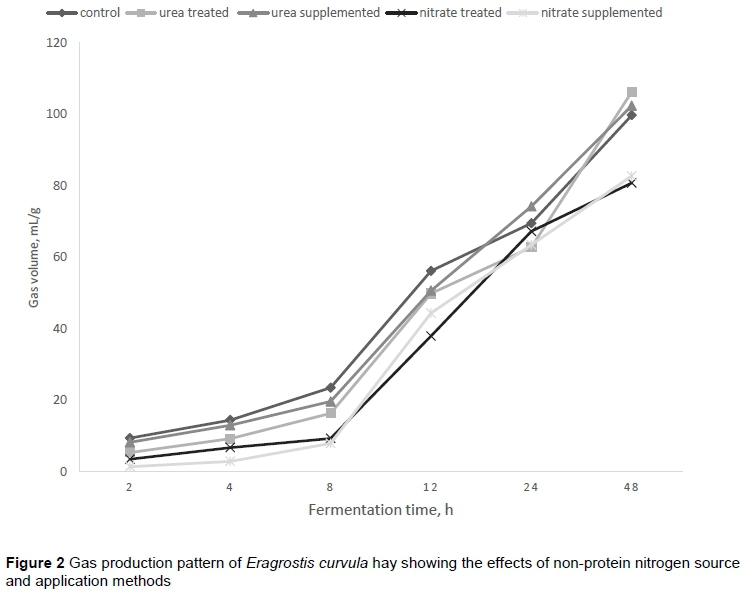
The influence of non-protein nitrogen source and application method on gas production characteristics, which relates to the slowly fermentable fractions (b), rate of gas production (c), and effective gas production of E. curvula hay, is presented in Table 2.
Generally, NPN source did not influence the slowly fermentable fraction compared with the untreated hay. However, urea inclusions resulted in a higher b-value and EGP than nitrate-included diets. In particular, the urea treatment resulted in a higher (P <0.05) b-value than the urea-supplemented diet. Neither NPN source nor application method had an effect (P >0.05) on the rate of gas production. The pattern of cumulative gas production over the 48-hour period is shown in Figure 2. Cumulative gas production increased slowly from 2 to 8 hours. However, as the time of incubation increased, gas production increased more rapidly until 24 hours. Total gas production in urea-supplemented and urea-treated diets tended to increase more rapidly from 12 to 48 hours. The concentration of methane as percentage of total gas produced across the in vitro incubation time is shown in Figure 1. Generally, methane concentration increased as incubation time increased (R2 = 0.99), and, regardless of the method of application, nitrate consistently reduced methane production over the 48-hour incubation period.
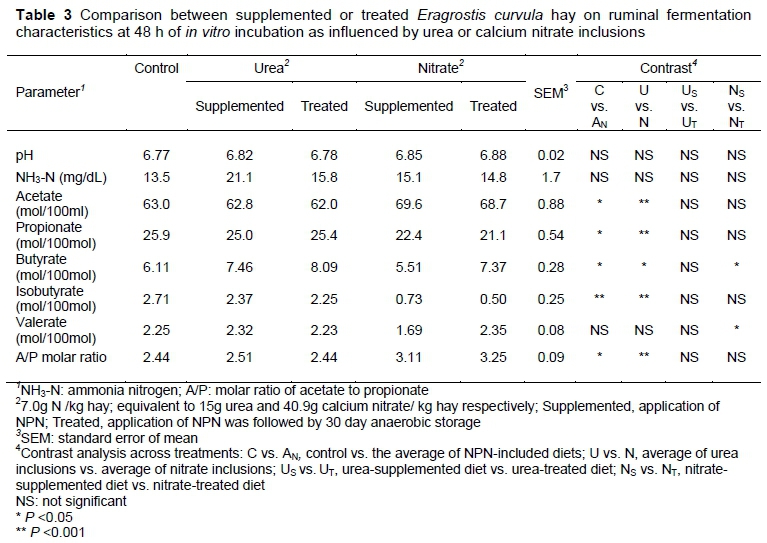
In vitro ruminal fermentation characteristics of E. curvula hay supplemented or treated with or without an NPN source are shown in Table 3. Neither rumen fluid pH nor rumen ammonia-nitrogen concentration was affected by NPN source or method of application (P >0.05). Inclusion of NPN generally increased acetate production compared with the control. Of the two NPN sources, nitrate resulted in higher acetate concentration than urea, regardless of the method of application. Regardless of the method of application, the acetate to propionate ratio was influenced by the addition of nitrogen source, with nitrate diets resulting in significantly higher acetate to propionate molar ratio than urea diets. The nitrate-treated diet had higher (P <0.001) butyrate and valerate concentration than the nitrate-supplemented diet. Other ruminal fermentation parameters did not show significant (P >0.05) differences as a result of NPN source or application method.
Discussion
The result shows that the feed is very low in crude protein, but high in NDF concentration. The very low crude protein content of the hay shows that the addition of an NPN source could improve its feeding value for ruminant animals. As expected, regardless of source, the addition of NPN resulted in higher in vitro digestibility of hay. The significant increase in IVOMD with NPN supplementation and NPN treatment confirms the earlier research hypothesis that very low quality feedstuff responds more readily to NPN inclusion (Mapato et al., 2010). Though total gas production was not significantly affected by the NPN application method, the treated diets resulted in a relatively higher IVOMD than the supplemented diets. The significant increase in digestibility after the urea treatment compared with urea supplementation agrees with previous studies (Uza et al., 2006; Verma et al., 2006). Mgheni et al. (1993) compared the effect of urea supplementation versus urea treatment and found a significant increase in in vitro digestibility, DM intake and growth performance of goats fed urea-treated rice straw, compared with urea-supplemented rice straw. Similarly, Tesfayohannes et al. (2013) reported significant improvement in the digestibility of E. curvula hay, wheat straw, barley straw, ryegrass, maize stover, and oat hay following urea treatment (ammoniation) for 35 days, but noted variations in the responses, depending on the type of feedstuff. Poorer-quality feeds were reported to show more improvement in digestibility compared with good-quality ones, as observed earlier by Goto et al. (1991). The improvement in feed digestibility over the control diet following feed treatment has been attributed to a significant reduction in NDF and the hemicellulose content of poor-quality feeds. This is as a result of the solubilization of fibre fractions resulting from linkage disintegration (Zorrilla-Rios et al., 1985; Mason et al., 1988; Tesfayohannes et al., 2013), disruption of cell wall structure by ammonia and increased swelling of fibre structure, and higher affinity for microbial attachment (Fahey et al., 1993).
Under anaerobic systems, nitrate is able to undergo various biochemical pathways, but the dissimilatory reduction pathway is favoured under very low redox potential, and often in the presence of high organic matter concentrations (Simon, 2002; Leng, 2008). A partial conversion of nitrate to ammonia may have occurred during the nitrate treatment process. This ammonia is partly responsible for the improvements in hay digestibility observed in this study, while the residual nitrate in the treated hay played an additional role in inhibiting methane production in vitro. The release of ammonia and the success of hydrolytic treatment of feedstuffs has been noted to depend on several factors such as biomass quality and conditions of storage, for example amount of moisture, duration of treatment and temperature (Shreck, 2013). Not much has been reported in literature about feed treatment with nitrate, therefore the reaction kinetics are not clearly understood. However, the results of this study show that a considerable breakdown of nitrate to ammonia may have occurred during the feed treatment to account for the significant differences in fermentation and digestibility parameters observed between the nitrate-supplemented and the nitrate-treated diets.
According to Inthapanya et al. (2011), an increase in the methane content of gas with a lengthened time of incubation and a more rapid gas production pattern after eight hours is probably because of the order of substrate degradation in in vitro rumen microbial environment. Soluble carbohydrates and NPN are readily utilized initially, while structural carbohydrates such as NDF and ADF, which generate more methane are degraded later, often requiring a considerable amount of energy utilization (Inthapanya et al., 2011). The ADF content of a feed may significantly affect its methane production pattern, particularly at the later stages of fermentation (Bannink et al., 2010). In the nitrate-supplemented and nitrate-treated diet, the amount of residual nitrate remaining across the incubation times may primarily influence the magnitude of methane reduction. Total methane production was significantly higher in nitrate-treated hay compared with the nitrate-supplemented hay, which indicates that some nitrate was hydrolysed to ammonia during storage, while a lower amount of residual nitrate resulted in a relatively higher value of methane output in nitrate-treated hay compared with nitrate-supplemented hay (15 versus 10 mL/g). The trend in methane concentration across time intervals was similar to that reported by Inthapanya et al. (2011). The significant reduction of methane in both nitrate inclusion methods is an indication that incorporating calcium nitrate via feed supplementation or during feed treatment could help to improve the utilization of poor-quality feed and might improve the efficiency of gross energy utilisation. Enteric methane accounts for 7 - 17% loss of the metabolizable energy of feed in ruminant diets during the digestion process (Robertson & Waghorn, 2002; Hristov et al., 2013). Thus, methane reduction can be regarded as a means of conserving the dietary energy of feedstuff. The significant decrease in methane production with dietary nitrate reported in this study was in agreement with earlier findings by various researchers (Nolan et al., 2010; Hulshof et al., 2012; Sophal et al., 2013). According to Gemeda & Hassen (2015), the feeding value of a feed is determined by its fermentation and digestibility characteristics. Although total gas production showed a significant reduction in nitrate diets compared with urea diets, it appears that the observed reduction is largely owing to the methane component of the total gas, as shown by the methane to total gas ratios. The ratios of methane to IVOMD and TVFA to methane indicate that nitrate diets provide a more efficient digestive process than the urea or control diets. The impact of the nitrate inclusion on rumen methanogenesis was more pronounced than their effect on substrate degradation, as shown by the high TVFA/CH4 and low CH4/IVOMD ratios of nitrate-treated and nitrate-supplemented diets compared with urea inclusions and the control diet. Ammonia nitrogen values in this study did not show significant differences across treatment groups. This may be an indication of comparable uptake of ammonia from the ammonia nitrogen pool for microbial biomass production or that the rate of NPN inclusions used in this study did not lead to accumulation of ammonia nitrogen.
The reduction in the molar proportion of propionate is consistent with previous research findings. In this study, there was a significant reduction in propionate concentration in both nitrate-supplemented and nitrate-treated diets. This agrees with previous studies (Nolan, 2010; Van Zijderveld et al., 2011). Nitrate has been noted to have a higher affinity for hydrogen ions than carbon dioxide, hence inhibiting the pathways that generate propionate and methane (Ungerfeld & Kohn, 2006). This explains the higher values of A/P molar ratios observed in this study.
While this study confirms the reports of earlier findings on the improvement in digestibility of roughages after urea treatment as shown in gas volume and IVOMD values, the result shows that increased digestibility of the hay was followed with a concomitant increase in methane production. This study also shows that replacing urea with nitrate in the treatment of poor-quality roughage resulted in a relatively lower digestibility improvement compared with urea, yet the level of improvement observed for nitrate was better than for the control diet. Moreover, a significant reduction in methane production could be achieved with nitrate treatment to justify its use, because it leads to a more efficient digestion process, as shown by methane/total gas ratio, and TVFA/methane ratio. An appreciable improvement in digestibility, coupled with a considerable reduction in methane production, indicates that calcium nitrate could be incorporated alone or with urea in the feed treatment of poor-quality roughages. However, the cost: benefit ratio needs to be acceptable for wider application.
Conclusion
The results of this experiment suggest that treating Eragrostis curvula hay with urea improved its utilization better than treatment with nitrate. However, with a reduction of enteric methane production and associated improvement in digestibility compared with the control, nitrate could be incorporated as a hydrolytic agent in treating poor-quality roughage feeds, with the aim of improving their utilization.
Acknowledgments
The authors are grateful to the European Community's Framework Programme (FP7/2007-2013) for financial support through the ANIMALCHANGE Project. Additional funding for the research was received from the Department of Science and Technology and National Research Foundation, South Africa. The corresponding author is also grateful to the University of Pretoria for a research bursary award.
Authors' Contribution
FAA and AH designed the study while its implementation, sample collection and data analysis was done by FAA. Both FAA and AH participated in results, statistics and interpretation. FAA wrote the draft manuscript while AH edited and proofread it.
Conflict of Interest Declaration
The authors declare that they have no competing interests.
References
Bannink, A., Smits, M.C.J., Kebreab, E., Mills, J.A., Ellis, J.L., Klop, A., France, J. & Dijkstra, J., 2010. Simulating the effects of grassland management and grass ensiling on methane emission from lactating cows. J. Agric. Sci. 148(01), pp.55-72. [ Links ]
Broderick, G. & Kang, J., 1980. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy Sci. 63: 64-75. [ Links ]
Diaz, E., Ouellet, D.R., Amyot, A., Berthiaume, R. & Thivierge, M.C., 2013. Effect of inoculated or ammoniated high- moisture ear corn on finishing performance of steers. Anim. Feed Sci. Tech. 182(1), 25-32. [ Links ]
Engels, E.A.N. & Van der Merwe, F.J., 1967. Application of an in vitro technique to South African forages with special reference to the effect to certain factors on the results. S. Afr. J. Agric. Sci. 10, 983-995. [ Links ]
Fahey, G.C., Bourquin, L.D., Titgemeyer, E.C. & Atwell, D.G., 1993. Postharvest treatment of fibrous feedstuffs to improve the nutritive value. In: H.G. Jung, D.R. Buxton, R.D. Hatfield, J. Ralph (eds). Forage cell wall structure and digestibility. ASA-CSSA-SSSA., Madison, WI, pp. 715-766. [ Links ]
Gemeda, B.S. & Hassen, A., 2014a. In vitro fermentation, digestibility and methane production of sixteen tropical perennial grass species. Crop Pasture Sci. 65, 479-488 [ Links ]
Gemeda, B.S., Hassen, A. & Odongo, N.E., 2014b. Effect of fibrolytic enzyme products at different levels on in vitro ruminal fermentation of low quality feeds and total mixed ration. J. of Anim. Plant Sci. 24 (5), 1293-1302 [ Links ]
Gemeda, B.S. & Hassen, A., 2015. Effect of tannin and species variation on in vitro digestibility, gas and methane production characteristics of tropical browse pla nts. Asian Austral. J. Anim. 28(2), 188. [ Links ]
Goering, H.K. & Van Soest, P.J., 1970. Forage fibre analyses, apparatus, reagents, procedures, and some applications. USDA Agricultural Handbook No. 379. pp. 1-20. USDA: Washington, DC. [ Links ]
Goto, M., Gordon, A.H. & Chesson, A., 1991. Effect of gaseous ammonia on barley straws showing different rumen degradability. J. Sci. Food Agric. 56(2), 141-153. [ Links ]
Hao Trinh, P., Do Ho, Q., Preston, T. R. & Leng, R. A., 2009. Nitrate as a fermentable nitrogen supplement for goats fed forage based diets low in true protein. Livest. Res. Ru Development, 21(1), unpaginated. Retrieved from http://lrrd.cipav.org.co/lrrd21/1/trin21010.htm. [ Links ]
Hristov, A.N., Oh, J., Firkins, J.L., Dijkstra, J., Kebreab, E., Waghorn, G., Makkar, H.P.S., Adesogan, A.T., Yang, W., Lee, C., Gerber, P.J., Henderson, B. & Tricarico, J.M., 2013. Mitigation of methane and nitrous oxide emissions from animal operations: I. A review of enteric methane mitigation options. J. Anim. Sci. 91(11), 5045-5069. [ Links ]
Hulshof, R.B.A., Berndt, A., Gerrits, W.J.J., Dijkstra, J., Van Zijderveld, S.M., Newbold, J.R. & Perdok, H.B., 2012. Dietary nitrate supplementation reduces methane emission in beef cattle fed sugarcane-based diets. J. Anim. Sci. 90(7), 2317-2323. [ Links ]
Inthapanya, S., Preston, T.R. & Leng, R.A., 2011. Mitigating methane production from ruminants; effect of calcium nitrate as modifier of the fermentation in an in vitro incubation using cassava root as the energy source and leaves of cassava or Mimosa pigra as source of protein. Livest Res. Rural Dev. 23 (2). [ Links ]
Le Huyen, T., Do, H.Q., Preston, T.R. & Leng, R.A., 2010. Nitrate as fermentable nitrogen supplement to reduce rumen methane production. Livest Res. Rural Dev. 22(8), pp. 1-8. [ Links ]
Lee, C. & Beauchemin, K.A., 2014. A review of feeding supplementary nitrate to ruminant animals: Nitrate toxicity, methane emissions, and production performance. Can J. Anim. Sci. 94(4), 557-570. [ Links ]
Leng, R.A., 2008. The potential of feeding nitrate to reduce enteric methane production in ruminants. A Report to the Department of Climate Change, Commonwealth Government of Australia. ACT Canberra, Australia. http://www.penambulbooks.com. [ Links ]
Li, L., Davis J., Nolan J. & Hegarty R., 2012. An initial investigation on rumen fermentation pattern and methane emission of sheep offered diets containing urea or nitrate as the nitrogen source. Anim. Prod. Sci. 52, 653-658. [ Links ]
Mapato, C., Wanapat, M. & Cherdthong, A., 2010. Effects of treatment of straw and dietary level of vegetable oil on lactating dairy cows. Trop Anim. Health Pro. 42(8), 1635-1642. [ Links ]
Mason, V.C., Hartley, R.D., Keene, A.S. & Cabby, J.M., 1988. The effect of ammoniation on the nutritive value of wheat, barley and oat straw. I. Changes and chemical composition in relation to digestibility in vitro and cell wall degradability. Anim. Feed Sci. Tech. 19(1), 159-171. [ Links ]
Mgheni, D.M., Kimambo, A.E., Sundstol, F. & Madsen, J., 1993. Influence of urea treatment or supplementation on degradation, intake and growth performance of goats fed rice straw diets. Anim. Feed Sci. Tech. 44(3), 209-220. [ Links ]
Mirzaei-Aghsaghali, A. & Meheri-Sis, N., 2011. Factors affecting mitigation of methane emission from ruminants I: Feeding strategies. Asian J. Anim. Vet. Adv. 6(9), 888-908. [ Links ]
Moreira Filho, M.A., Alves, A.A., Vale, G.E.S.D., Moreira, A.L. & Rogério, M.C.P., 2013. Nutritional value of hay from maize-crop stubble ammoniated with urea. Rev. Ciênc. Agron. 44(4), 893-901. [ Links ]
Mould, F.L., Morgan, R., Kliem, K.E. & Krystallidou, E., 2005. A review and simplification of the in vitro incubation medium. Anim. Feed Sci. Tech. 123, 155-172. [ Links ]
Newbold, J.R., Van Zijderveld, S.M., Hulshof, R.B.A., Fokkink, W.B., Leng, R.A., Terencio, P., Powers, W.J., Van Adrichem, P.S.J., Paton, N.D. & Perdok, H.B., 2014. The effect of incremental levels of dietary nitrate on methane emissions in Holstein steers and performance in Nelore bulls. J. Anim. Sci. 92(11), 5032-5040. [ Links ]
Nolan, J.V., Hegarty, R.S., Hegarty, J., Godwin, I.R.& Woodgate, R., 2010. Effects of dietary nitrate on fermentation, methane production and digesta kinetics in sheep. Anim. Prod. Sci. 50(8), 801-806. [ Links ]
Ørskov, E.R. & Mcdonald, I., 1979. The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci-Cam. 92: 499-503. [ Links ]
Preston, T.R. & Leng, R.A., 1987. Matching livestock systems to available resources in the tropics and sub tropics. Penambul Books, Armidale, Australia. Web edition. http://utafoundation.org/P&L/preston&leng.htm [ Links ]
Robertson, L.J. & Waghorn, G.C., 2002. Dairy industry perspectives on methane emissions and production from cattle fed pasture or total mixed rations in New Zealand. In: Proceedings of the New Zealand Society of Animal Production. 62, 213-218. [ Links ]
Santoso, B., Mwenya, B., Sar, C. & Takahashi, J., 2007. Methane production and energy partition in sheep fed timothy hay silage- or hay-based diets. J. Anim. Sci. Vet-JITV. 12(1), 27-33. [ Links ]
SAS, 2010. SAS version 9.3. SAS Institute Inc., Carry, NC, USA. [ Links ]
Schroeder, G.F. & Titgemeyer, E.C., 2008. Interaction between protein and energy supply on protein utilization in growing cattle: a review. Livest Sci. 114(1), 1-10. [ Links ]
Shreck, A.L., 2013. Use of alkaline treated crop residues as partial grain replacements for finishing cattle. Ph.D. dissertation in Animal Science, University of Nebraska-Lincoln. [ Links ]
Simon, J., 2002. Enzymology and bioenergetics of respiratory nitrite ammonification. FEMS Microbiol Rev. 26(3), 285-309. [ Links ]
Sophal, C., Khang, D.N., Preston, T.R. & Leng, R.A., 2013. Nitrate replacing urea as a fermentable N source decreases enteric methane production and increases the efficiency of feed utilization in Yellow cattle. Livest. Res. Rural Dev. 25(7). [ Links ]
Takahashi, J., 2011. Some prophylactic options to mitigate methane emission from animal agriculture in Japan. Asian Austral. J. Anim. 24(2), 285-294. [ Links ]
Tavendale, M.H., Meagher, L.P., Pacheco, D., Walker, N., Attwood, G.T. & Sivakumaran, S., 2005. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Tech. 123, 403-419. [ Links ]
Tesfayohannes, S.T., Nsahlai, I.V. & Bengaly, K., 2013. Effect of urea treatment and concentrate proportions on dry matter degradation of different roughages in the rumen of Jersey cows. Global J. Sci. Frontier Res. (D). 13(8). [ Links ]
Theodorou, M.K., Williams, B.A., Dhanoa, M.S., McAllen, A.B. & France, J., 1994. A simple gas production method using pressure transducers to determine the fermentation kinetics of ruminant feed. Anim. Feed Sci. Tech. 48(3), 185-197. [ Links ]
Tilley, J.M.A. & Terry, R.A., 1963. A two-stage technique for the in vitro digestion of forage crops. Grass Forage Sci. 18(2), 104-111. [ Links ]
Ungerfeld, E.M. & Kohn, R.A., 2006. The role of thermodynamics in the control of ruminal fermentation. In: Sejrsen, K., Hvelplund, T. & Nielsen, M.O.(Eds.), Ruminant physiology: Digestion, Metabolism and Impact of Nutrition on Gene Expression, Immunology and Stress. Wageningen Academic Publishers, Wageningen, Netherlands, pp. 55 85. [ Links ]
Uza, D.V., Barde, R.E. & Ayoade, J.A., 2006. The effects of urea treated cassava peels as supplement to West African dwarf (WAD) goats grazing natural pastures. Nig. Vet. J. 26(1), 1-9. [ Links ]
Vadiveloo, J., 2003. The effect of agronomic improvement and urea treatment on the nutritional value of Malaysian rice straw varieties. Anim. Feed Sci. Tech. 108(1), 133-146. [ Links ]
Van Zijderveld, S.M., Gerrits, W.J.J., Apajalahti, J.A., Newbold, J.R., Dijkstra, J., Leng, R.A. & Perdok, H.B., 2010. Nitrate and sulfate: Effective alternative hydrogen sinks for mitigation of ruminal methane production in sheep. J. Dairy Sci. 93, 5856-5866. [ Links ]
Van Zijderveld, S.M., Gerrits, W.J.J., Dijkstra, J., Newbold, J.R., Hulshof, R.B.A. & Perdok, H.B., 2011. Persistency of methane mitigation by dietary nitrate supplementation in dairy cows. J. Dairy Sci. 94, 4028-4038. [ Links ]
Verma, A.K., Singh, P., Dass, R.S. & Mehara, U.R., 2006. Impact of feeding urea ammoniated and urea supplemented wheat straw on intake and utilization of nutrients in crossbred cattle. Indian J. of Anim. Sci. 76(6), [ Links ]
Wanapat, M., Polyrach, S., Boonnop, K., Mapato, C. & Cherdthong, A., 2009. Effect of treating rice straw with urea and calcium hydroxide upon intake, digestibility, rumen fermentation and milk yield of dairy cows. Livest. Sci. 125, 238-243. [ Links ]
Webb, E.C., 1994. Synthesis of long chain fatty acids in ruminants and their effects on meat quality. Chapter 3, page 58-59. [ Links ]
Widiawati, Y. & Thalib, A., 2007. Comparison fermentation kinetics (in vitro) of grass and shrub legume leaves. The pattern of VFA concentration, estimated CH4 and microbial biomass production. J. Anim. Sci. Veteriner-JITV. 18(2). [ Links ]
Zorrilla-Rios, J., Owens, F.N., Horn, G.W. &, McNew, R.W., 1985. Effect of ammoniation of wheat straw on performance and digestion kinetics in cattle. J. Anim. Sci. 60, 841-821. [ Links ]
Received 14 October 2016
Accepted 19 January 2017
First published online 6 February 2017
# Corresponding author: Abubeker.hassen@up.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}