










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.46 n.4 Pretoria 2016
http://dx.doi.org/10.4314/sajas.v46i4.10
Mangosteen peel can reduce methane production and rumen biohydrogenation in vitro
P. ShokryzadanI; M.A. RajionII; Y.M. GohI, II, #; I. IshakII; M.F. RamleeII; M. Faseleh JahromiI, III; M. EbrahimiII, #
IInstitute of Tropical Agriculture, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
IIFaculty of Veterinary Medicine, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
IIIAgriculture Biotechnology Research Institute of Iran (ABRII), East and North-East Branch, P.O.B. 91735, 844 Mashhad, Iran
ABSTRACT
Mangosteen peel (MP), an agricultural by-product of tropical countries, has been reported to contain condensed tannins and saponins, which can affect rumen microbes to reduce enteric methane emission. In the present study, the effects of mangosteen peel on in vitro ruminal fermentation, gas production, methane production, fatty acid biohydrogenation, and microbial population were investigated. Results showed that MP at medium and high levels (25 % and 50 % replacing alfalfa) were able to reduce (P <0.05) in vitro methane production without affecting volatile fatty acid (VFA) production and the pH of the substrate. The lowering effect of MP on methane production was because of suppression of the rumen microbial populations, especially total protozoa and total methanogens. MP at the higher level (50%) reduced (P <0.05) the amounts of biohydrogenation for linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3) and the total C18 unsaturated fatty acids (UFA) owing to the reduction of the Butyrivibrio fibrisolvens population, that is, the most important rumen microorganism involved in the biohydrogenation process. In conclusion, mangosteen peel has potential to be used in ruminant livestock feeds, with the advantage of reducing ruminal methane production and biohydrogenation, without adverse effects on ruminal pH and VFA production.
Keywords: agricultural by-product, saponins, condensed tannins, gas production, volatile fatty acid, microbial quantification
Introduction
The rumen houses billions of microbes, predominantly bacteria, protozoa and fungi. It is a suitable place for microbes to multiply because of its anaerobic environment, good mixing of inner materials, and constant pH and temperature. Microbes in the rumen help cellulose and carbohydrate digestion to simple sugars and VFAs to be used by the host (Frandson et al., 2009). Other abundant products of microbial digestion in the rumen are carbon dioxide, methane and ammonia (Hobson, 1997). Important cellulolytic bacteria of the rumen include Ruminococcus albus, Ruminococcus flavefaciens and Fibrobacter succinogens (Faseleh Jahromi et al., 2013). Rumen protozoa consist mainly of Entodinium, Holotrichs, Spirotrichs, Methanobacterium ruminantium and Methanosarcina barkeri, of which the latter two are important rumen methanogens (Faseleh Jahromi et al., 2013).
Ruminant enteric fermentation is responsible for 15 % to 20 % of global methane emissions (Crutzen et al., 1986; Bhatta et al., 2015; Buccioni et al., 2015), which is approximately 4 % of total greenhouse gas production (FAO, 2010). In addition, methane produced during ruminal fermentation causes a loss of 11 % to 13 % of digestible energy (McDonald et al., 2002). Hence, many techniques have been attempted over the years to reduce methane production by ruminants, usually employing dietary manipulation, for example dietary supplementation of antibiotics, halogenated compounds, oils, organic acids, propionate precursors, direct-fed microbials, bacteriocins, vaccines and secondary metabolites of plants.
In addition, owing to high levels of fatty acid biohydrogenation in the rumen, ruminant products usually contain high levels of saturated fatty acids (SFA) and low levels of polyunsaturated fatty acids (PUFA) (Morimoto et al., 2005). Because of increased consumer awareness of the effects of diet on their health, researchers have attempted to alter the fatty acid profile of ruminant meat products to reduce SFA and increase PUFA by lowering biohydrogenation in the rumen, with some success in lamb (Rajion et al., 2001) and chevon (Ebrahimi et al., 2011).
Many tropical plants and their extracts, which normally contain high or medium levels of secondary metabolites, have been used in ruminant feeds, and have been shown to affect the rumen fermentation process and end-products by altering the rumen microbial populations. For example, saponins and condensed tannins have been shown to exert a specific effect against rumen protozoa (Lu & Jorgensen, 1987; Getachew et al., 2000; Wang et al., 2000). Alteration of the rumen microbial population can also affect production of methane and biohydrogenation of fatty acids in the rumen.
Mangosteen (Garcinia mangostana L.), a native tropical fruit that is found in South East Asian countries such as Indonesia, Malaysia, Thailand and Vietnam, consists of the peel (exocarp), pulp and seed. Mangosteen peel contain condensed tannins (16.8% w/w dry matter (DM)) and saponins (10.0% w/w DM), which have been reported to have some effect against rumen protozoa, while the rest of the rumen biomass remains unaltered (Ngamsaeng & Wanapat, 2005). Hence, because of the availability and abundance of mangosteen in Malaysia, it could be used as a feed supplement for ruminants to reduce methane production and fatty acid biohydrogenation in the rumen. In the present study, the effects of mangosteen peel on in vitro gas production, methane production, rumen fermentation, fatty acid biohydrogenation and changes in microbial population were investigated.
Materials and Methods
Fresh mangosteen fruits were purchased from a local market in Serdang, Selangor, Malaysia. The fruit pulps were removed, and the peels were kept to prepare MP powder. The peels were dried in the oven at 60 oC for 72 h. After that, the dried peel were ground and filtered through a 1-mm sieve to obtain the MP powder.
Total saponin content was determined according to Makkar et al. (2007), based on the vanillin-sulfuric acid colorimetric reaction. The results were expressed as mg diosgenin equivalent/g DM of MP. Condensed tannins were determined by the butanol-HCl-iron method according to Porter et al. (1985), as described by Jayanegara et al. (2010). In brief, 0.5 mL of the extract was mixed with 0.1 mL ferric reagent (2.0 g ferric ammonium sulfate in 100 mL of 2 N HCl), followed by 3 mL butanol-HCl (95:5 v/v), and the tube was vortexed. Subsequently, the tube was heated in a boiling water bath at 100 oC for 60 min. The absorbance was read at 550 nm.
In the present study, three treatments were used, namely the control (C, 50% concentrate + 50% alfalfa without MP); medium MP (MMP, 50% concentrate + 25% alfalfa + 25% MP); and high MP (HMP, 50% concentrate + 50% MP). Gas production was determined by the procedure described by Fievez et al. (2005) with minor modifications (amount of substrate and time of carbon dioxide infusion were changed). Rumen fluid was collected before the morning feeding from two rumen-fistulated (Bar Diamond Inc, PO Box 60, Parma, Idaho, USA) goats fed an equal weight mixture of 40% concentrate and 60% alfalfa twice daily at 08:00 and 18:00. Rumen fluid was collected in a thermos flask and transported directly to the laboratory. After that, the rumen fluid was filtered with four layers of cheesecloth, and stirred with a magnetic stirrer while being continuously flushed with carbon dioxide. Phosphate buffer (containing 28.8 g Na2HPO4.12H2O, 6.1 g NaH2PO4.H2O and 1.4 g NH4Cl per litre distilled water, pH 6.8) and bicarbonate buffer (containing 39.2 g of NaHCO3 per litre distilled water, pH 6.8) were mixed (4:1 v/v) with the filtered rumen fluid. A total of 0.25 g dried feed material of each treatment was added to a 100-mL calibrated syringe, and 30 mL of the rumen fluid mixture were also added to each syringe. All the syringes were incubated at 39 °C for 24 h. At the same time, volumes of produced gas were measured after 0 h, 2 h, 4 h, 6 h, 8 h, 10 h, 12 h and 24 h incubation. At each reading time, the syringes were shaken carefully to ensure complete mixing of the incubated contents. Standard hay (University of Hohenheim, Stuttgart, Germany) with an estimated gas production of 49.61 mL/g DM was used to calibrate the in vitro gas production system. After 24 h in vitro incubation, samples of substrate were obtained from the syringes to determine methane and VFA production, using gas-liquid chromatography (Agilent Technologies, Agilent 7890A, Palo Alto, Calif, USA).
After 24 h in vitro incubation, samples of gas were obtained and the concentration of methane for each sample was determined by injection of 500 μL gas from each syringe to the gas-liquid chromatography (Agilent 5890 Series Gas Chromatograph, Wilmington, Del, USA), equipped with a HP-Plot Q column (30 m χ 0.53 mm χ 40 μM) (Agilent Technologies, Wilmington, Del, USA). The carrier gas was nitrogen with a flow rate of 3.5 mL/min, and the temperature of the oven was set at 50 °C. Methane was detected using a thermal conductivity detector in 1 min of run time. Calibration was completed using standard gas prepared by Scott Specialty Gases (Supelco, Bellefonte, Pa, USA), which contains 1% of methane, carbon dioxide, carbon monoxide, oxygen and hydrogen (Faseleh Jahromi et al., 2013).
The amounts of VFA produced were determined according to the method of Ebrahimi et al. (2014) with minor modifications in the gas-liquid chromatography conditions. The details are described below. After sample preparation, 0.5 mL supernatant and 0.5 mL internal standard (20 mmol, 4-methylvaleric acid) were transferred to 2-mL glass tubes and 1 μL of each sample was injected to a gas-liquid chromatography (Agilent Technologies, Agilent 7890A, Palo Alto, Calif, USA) with a flame ionization detector with a Quadrex 007 Series (Quadrex Corporation, New Haven, Conn, 06525 USA) bonded phase fused silica capillary column (15 m, 0.32 mm ID, 0.25 μm film thickness). The temperature of column was set at 70 to 150 °C with temperature programming at the rate of 7 °C/min increments for optimal separation. Temperature of oven, FID and injector were 160 °C, 250 °C, and 230 °C, respectively. Nitrogen with the flow rate of 1.0 mL/min was used as carrier gas. Acetate (20 mmol), propionate (10 mmol) and butyrate (10 mmol) were used as standard solutions to identify the peaks, based on their retention times (Ebrahimi et al., 2015).
Total fatty acids were extracted from the whole syringe content after 24 h of incubation, based on the method of Folch et al. (1957), modified by Rajion et al. (1985), as described by Ebrahimi et al. (2012) using chloroform/methanol 2:1 (v/v) containing butylated hydroxy toluene to prevent oxidation during sample preparation. After complete separation, the lower phase was collected in a round bottom flask and rotary evaporated (Laborota 4000-efficient; Heidolph, Germany) at 70 °C. An internal standard, heneicosanoic acid (C21:0) (Sigma Chemical, St. Louis, Mo, USA), was added to each sample before transmethylation to determine the individual fatty acid concentration in the sample. Transmethylation of the extracted fatty acids to their fatty acid methyl esters (FAME) was carried out using potassium hydroxide (KOH) in methanol and 14% methanolic boron trifluoride (BF3). The FAME were separated by gas chromatography (Agilent 7890N), using a Supelco SP 2560 capillary column of 100 m x 0.25 mm ID χ 0.2 μm film thickness (Supelco, Bellefonte, Pa, USA). The amount of 1 μL was injected by an auto sampler (Agilent Auto Analyzer 7683 B series, Agilent Technologies, Santa Clara, Calif, USA) into the chromatograph equipped with a split/splitless injector and an FID. The carrier gas was nitrogen at a flow rate of 1.2 mL/min. The split ratio was 1 : 20 after injection of 1 μL of the FAME. The injector temperature was programmed at 250 °C, and the detector temperature was 270 °C. The column temperature programme started to run at 150 °C, for 2 min, warmed to 158 °C at 1 °C/min, held for 28 min, warmed to 220 °C at 1 °C/min, and then held for 20 min to achieve satisfactory separation. The peaks of samples were identified, and concentrations calculated based on the retention time and peak area of known standards (Sigma Chemical). The fatty acid concentrations are expressed as g/100 g of the sum of identified peaks measured in each sample. The fatty acid concentrations are expressed as percentage (%) of total identified fatty acids. A reference standard (mix C4-C24 methyl esters; Sigma-Aldrich, Inc., St. Louis, Mo, USA) and CLA standard mix (cis-9 trans-11 and trans-10, cis-12 CLA, Sigma-Aldrich, Inc., St. Louis, Mo, USA) was used to determine recoveries and correction factors to determine individual FA composition.
The biohydrogenation (BH) of PUFA was calculated by the decrease in the amounts of PUFA after 24 h incubation from the initial PUFA at start point (0 h) of incubation as described by Jayanegara et al. (2011):
Biohydrogenation (%) = [PUFA(0h) - PUFA (24h) ÷ PUFA(0h)] x 100
PUFA (0 h): concentration (g/100g of fatty acid) of PUFA at 0 h incubation; PUFA (24 h): concentration (g/100g of FA) of PUFA at 24 h of incubation.
Concentrations of ammonia were determined with the colorimetric method described by Solorzano, (1969), based on the standard curve of ammonium sulfate standard solution, and its relation to the intensity of colour produced in the samples. The intensity of developed colour was determined by measuring optical density of samples at 420 nm using a spectrophotometer (Spectro UV-Vis Auto, Labomed Inc., Culver City, Calif, USA).
One and half millilitres of each rumen fluid sample were used for absolute microbial quantification using real-time polymerase chain reaction. Total DNA was extracted from each sample with the QIA amp DNA Stool Mini Kit (Qiagen Inc., Valencia, Calif, USA) according to the manufacturer's instructions. The extracted DNAs were stored at -20 °C until used.
The quantification assay was based on the standard curve method. To prepare the standard curves, numbers of copies of the target genes were plotted against quantification cycle (Cq). The Cq values were obtained from tenfold serial dilutions of PCR products from plasmid DNA of each target microbial group (total bacteria, total methanogens, total protozoa or Butyrivibrio fibrisolvens). Specific primers for each group of target microorganisms, indicated in Table 1, were used to amplify target genes using conventional PCR. The PCR reaction properties have been described by Faseleh Jahromi et al. (2013). The PCR products of the target microorganisms were run in 1% (w/v) agarose gel, and specific bands were purified using the MEGAquick-spinTM purification kit (iNtRON Biotechnology, Korea). The purified PCR products were then cloned into the PCR 2.1 TOPO vector using PCR 2.1 TOPO TA Cloning Kit (Invitrogen Ltd., USA) according to the manufacturer's instructions. The purity and concentration of plasmid DNA in each sample were measured with a Nanodrop ND-1000 spectrophotometer (Implen NanoPhotometer™, Germany). The number of copies of the plasmid DNA per mL of elution buffer was calculated with this formula, which is available online (www.uri.edu/research/gsc/resources/cndna.html):
Number of copies = Amount of DNA (μg/mL) x 6.022 x 1023/ Length (bp) x 109 x 650
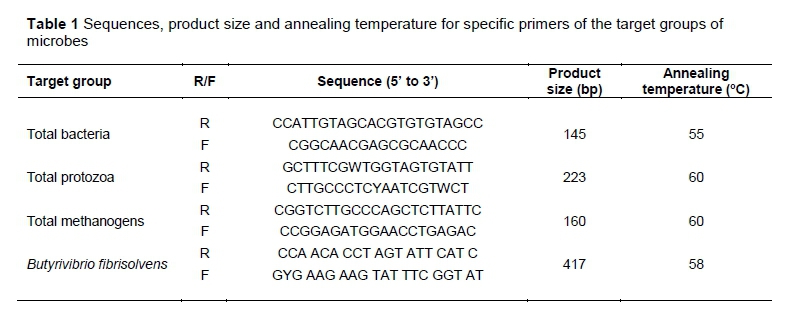
Primers used to quantify the population of various groups of microorganisms are the same as those used to amplify target genes for standard curve preparation (Table 1). Real-time PCR was performed with the BioRad CFX96 Touch (BioRad, USA) using optical grade plates. The PCR reaction was performed on a total volume of 25 μl using the iQTMSYBR Green Supermix (BioRad, USA). Each reaction consisted of 12.5 μl SYBR Green Supermix, 1 μl forward primer, 1 μl reverse primer, 2 μl DNA samples, and 8.5 μl nuclease-free water. No-template control was included in the real-time PCR amplification to rule out cross-contamination. These reaction conditions were applied: an initial 5-min denaturation at 94 °C, followed by 40 cycles of denaturation at 94 °C for 20 s, primer annealing (annealing temperatures for primers are described in Table 1) for 30 s, and extension at 72 °C for 20 s (Navidshad et al., 2012). To confirm the specificity of amplification; melting curve analysis was carried out after the last cycle of each amplification; and PCR products were verified on a 1% (w/v) agarose gel that runs for 40 min at 80 V. The expected sizes of amplified fragments were presented in Table 1.
The amplification efficiency was calculated using the following equation:
Ε (%) = (10-1/slope - 1) x 100
In this equation, E is 100% if a tenfold dilution of DNA template results in a Cq difference of 3.32. Only the data generated from reactions with efficiency between 90 and 110% were used for further analysis.
All the experiments were carried out twice, each treatment in triplicate. Statistical analysis of experimental data was performed by the one-way analysis of variance (one-way ANOVA) procedure of Statistical Analysis System (SAS, 2008) version 9.2. Significantly different means were then further differentiated with Duncan's multiple range test procedure. The results are presented as mean ± standard error of the means. Differences between means with P <0.05 were regarded as being statistically different. Results of microbial quantification are presented as log10 of copy number/mL.
Results
In the present study, the total amounts of condensed tannin and saponin of MP were determined as 16.53 and 10.71 % of DM, respectively.
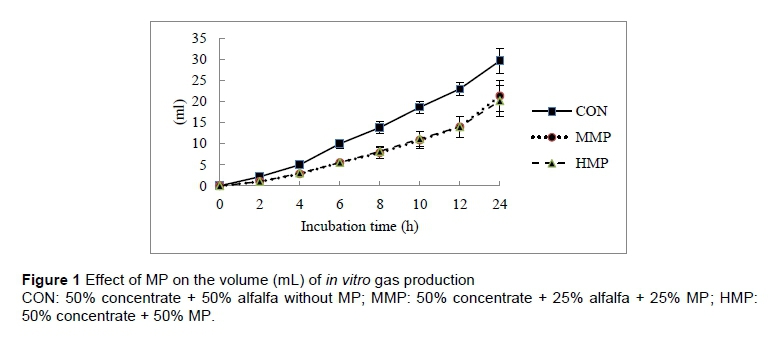
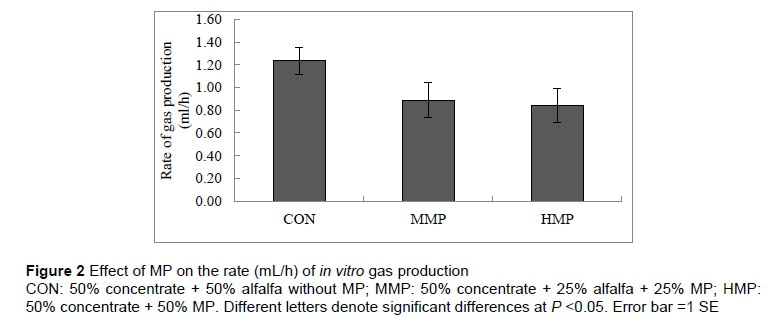
Figure 1 shows the effect of MP on the total volume (mL) of gas production. There was no significant difference between the treatment groups in terms of the amount of gas produced at any reading time-point, although the control group produced numerically higher amounts of gas than the MMP and HMP groups. Similarly, there was no significant difference between the treatment groups in terms of the rate (mL/h) of in vitro gas production after 24 h incubation (Figure 2). The control and HMP group showed numerically the highest (1.2 mL/h) and lowest (0.8 mL/h) rates of gas production, respectively.
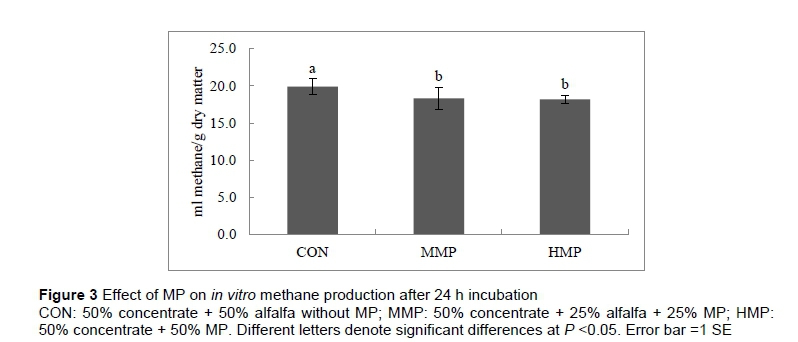
Figure 3 shows the effect of MP on in vitro methane production after 24 h of incubation. Results show a higher (P <0.05) methane production for the control group (19.9 mL methane/g DM) than the two groups containing MP, namely MMP (18.3 mL methane/g DM) and HPM (18.2 mL methane/g DM). In addition, the results showed that MP reduced (P <0.05) the amounts of produced methane per unit of total gas production (0.08 ± 0.01 and 0.08 ± 0.00 for the MMP and HMP groups, respectively, in comparison with 0.09 ± 0.01 for control group; data not shown).
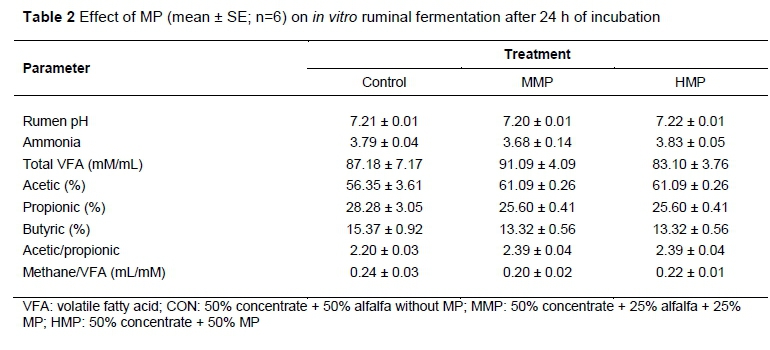
The amounts of VFA ranged from 83.1 to 91.1 mM/mL. There was no significant difference in terms of production of the three measured VFAs (acetic, propionic and butyric acids), acetate/propionate ratio, and amounts of methane per unit of VFA production among the three treatment groups. However, the production of acetic acid was numerically higher for both groups containing MP compared with the control group. However, for propionic and butyric acid production, the control group showed numerically higher amounts than the two groups with MP.
Table 2 shows the results of in vitro ammonia production after 24 h of incubation. The ammonia production in the rumen fluid samples did not show any significant difference among the three treatment groups, and ranged from 3.68 to 3.83 ppm/mL. Hence, MP had no effect on ammonia production. Results of changes in the pH of rumen fluid from the use of MP are presented in Table 3. The pH of rumen fluid, which ranged from 7.20 to 7.22, did not show any significant difference among the three treatment groups.
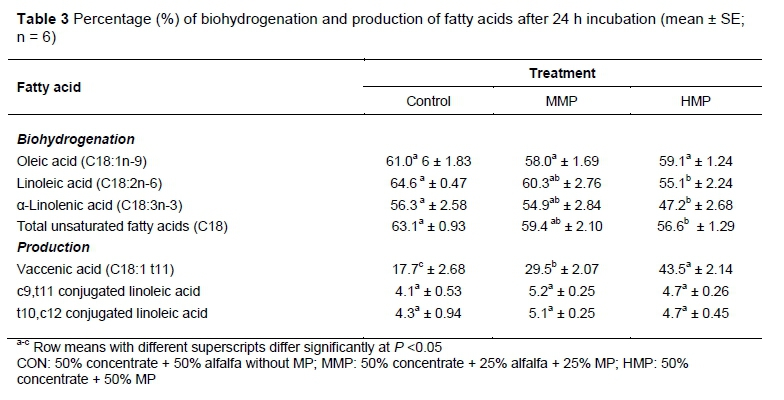
Table 3 shows the percentage of biohydrogenation of oleic acid (C18:1n-9), linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3) and total UFA, percentage of production of vaccenic acid (C18:1 t11), c9,t11 conjugated linoleic acid and t10,c12 conjugated linoleic acid (CLA) at 24 h of incubation. Based on the results, the range of biohydrogenation was from 47.2 % to 64.6 %. Although the biohydrogenation of oleic acid (C18:1n-9) did not show any difference (P >0.05) among the three treatment groups, MP in both levels (25% in MMP group and 50% in HMP group) reduced (P <0.05) the biohydrogenation of linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3) and the total UFA (C18). The amounts of vaccenic acid (C18:1 t11) production were different (P <0.05) among the three treatment groups. Production of the two isomers of CLA, c9,t11 and t10,c12, which are derivatives of linoleic acid, did not show any difference (P >0.05).
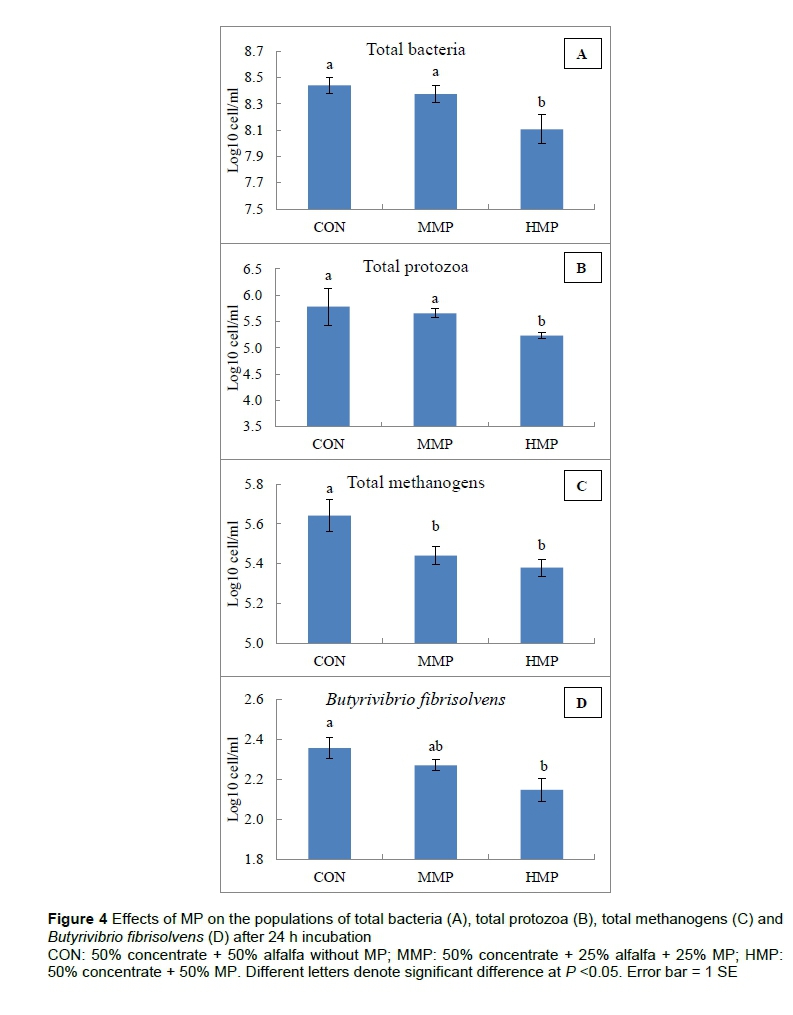
Figure 4 shows the results of the microbial quantification assay. The high level (50%) of MP reduced (P <0.05) the population of total protozoa and total bacteria in comparison with the MMP and control groups. The MMP and control groups did not show any significant difference in terms of population of total protozoa and total bacteria. The population of total methanogens was reduced (P <0.05) in the two treatment groups containing MP compared with the control group. However, there was no significant difference between the populations of total methanogens in the MMP and HMP groups. The population of Butyrivibrio fibrisolvens was in the order of CON > MMP > HMP, with significant difference (P <0.05) between HMP and CON.
Discussion
The rumen contains millions of microbes, namely bacteria, protozoa and fungi. Interaction of these microbes in the rumen environment provides the ruminant with the ability to digest cellulose and carbohydrate to simple sugars and VFA, which can be used by the host (Frandson et al., 2009), and produces certain components, such as carbon dioxide, methane and ammonia (Hobson, 1997). Any change in the rumen environment, which could be because of changes in the diet, might cause an alteration in the population of the rumen resident microbes, which would lead to change in the amounts of fermentation end-products. Among these end-products, methane is one of the most important, and has been studied extensively. According to the US Environmental Protection Agency (2007), ruminants produce huge volumes (about 80) of methane every year. Methane is considered a harmful product, not only because it is a greenhouse gas, but because it is responsible for losing some part (11 % to 13 %) of digestible energy (McDonald et al., 2002). Hence, there is continued research to find ways of mitigating methane production in ruminants without affecting VFA production.
After the ban on the usage of antibiotics in livestock feeds by the European Union (OJEU, 2003) because of increasing risk of passing on antibiotic resistance to human pathogens, plant extracts and plant secondary metabolites, such as essential oils, tannins, saponins and flavonoids, to improve livestock productivity, has been regarded as natural alternatives to antibiotics (Cieslak et al., 2012). This strategy was also intended to reduce environment pollutants, including lowering methane production by ruminants, and decrease phosphorus and nitrogen in manure (Makkar et al., 2009).
As stated, the mangosteen (Garcinia mangostana), which contains secondary metabolites such as tannins and saponins, can affect rumen fermentation and methane production. Since mangosteen production is extensive, leading to high availability of the fruit in Malaysia, mangosteen peel can be regarded as a potential feed source or feed supplement for ruminants. In the present study, MP was compared with alfalfa to evaluate its effects on in vitro ruminal parameters, such as total gas, methane, VFA and ammonia production, and biohydrogenation of fatty acids, pH and microbial population of rumen fluid in vitro.
Based on the results, although MP had no significant effect on total gas production, it reduced the total produced gas numerically. Besides, MP had no adverse effect on the pH of rumen fluid and VFA production. VFA production in the rumen depends on the available substrates and the microbial population of the rumen (Szumacher-Strabel & Cieslak, 2012). In a normal ruminal fermentation process, the gas produced is directly correlated to the level of VFA produced. However, in the present study, the reduction of total gas and the rate of gas production in the treatments containing MP, without decreasing the level of VFA produced, indicated that the fermentation kinetics had been shifted towards more VFA production than the that of gases such as methane, hydrogen and carbon dioxide. The reduced methane production (Figure 3) can be attributed to the lowered effect of the secondary metabolites of MP, such as saponins and condensed tannins on methane production. These results confirmed those of Animut et al. (2008), Tan et al. (2011) and Cieslak et al. (2012), who showed reduced methane emissions with tannin inclusion in cows with rumen cannula, rumen fistulated cattle, and goats fed increasing levels of Kobe lespedeza. In addition, the current results agree with those of Theodoridou et al. (2011), who incubated Sainfoin (containing condensed tannins) with buffered rumen fluid for 3.5 h and 24 h and observed no effect on VFA production by tannins. Cieslak et al. (2012), who supplemented Vaccinium vitisidaea (containing 2 g of tannins/kg dietary DM) in the diet of cows, reported that tannins did not have any significant effect on the VFA production. This could be because of the adaptation of rumen microorganisms to the tannins (Patra & Saxena, 2011). However, another study by Hassanat & Benchaar (2013), using various sources and concentrations of tannins on rumen microbial fermentation in vitro, showed that gas and total VFA production decreased as tannin concentration increased. This discrepancy could be owing to the usage of various types of tannins, because in the results obtained by Hassanat and Benchaar (2013), tannins from Vaíonea were the only tannin source that reduced methane production without affecting VFA concentration.
It has been reported that saponins and condensed tannins may reduce methane production by decreasing the methanogen populations through a reduction in the numbers of protozoa, and lowering methanogenesis activity of protozoa associated methanogens (Hess et al., 2003; Guo et al., 2008; Tan et al., 2011). In addition, Tavendale et al. (2005) proposed that condensed tannins reduce methane emissions from ruminants directly by inhibiting the growth of methanogens, and indirectly by reducing fibre digestion which decreases hydrogen production. Since the suppressing effect of secondary metabolites on the ruminal protozoa and methanogens have been already reported, the effects of MP on methane production in the current study could be similar to those observed in these reports. To confirm this, the present study observed decreased in the population of total methanogens (Figure 4).
Besides, MP reduced the population of total protozoa in the treatment group containing high level (50%) of MP compared with the control group. These results confirm the effect of secondary metabolites of MP on the population of rumen protozoa and methanogens. This is consistent with the results of Animut et al. (2008), who showed that increased levels of tannins (50, 101, 151 g/kg DM) in goat diets lowered the population of rumen protozoa. In another study, Cieslak et al. (2012) reported anti-protozoal effects of tannins (2 g tannins/kg dietary DM) in cows. However, in their experiment (Cieslak et al 2012), tannin had no significant effect on the populations of total bacteria and methanogens, whereas in the present study, the populations of total bacteria and total methanogens were reduced by using 50 % MP. This discrepancy could be attributed to the types of tannins, their origin and supplementation levels (Patra & Saxena, 2011).
Saponin, a secondary metabolite, has been reported to affect ruminal conditions. Hess et al. (2003), who compared three saponin-rich tropical fruits (Sapindus saponaria, Enteroíobium cycíocarpum and Pitheceííobium saman) reported that only S. saponaria significantly lowered protozoal population and methane production, without affecting the population of methanogens, in comparison with the control group. In another study (Guo et al., 2008), the addition of saponins extracted from tea seeds significantly decreased methane production during ruminal fermentation. It also reduced the population of protozoa, but not methanogens.
In the present study, the population of Butyrivibrio fibrisoívens, which are responsible for ruminal biohydrogenation, decreased numerically and significantly in the MMP and HMP groups, respectively, in comparison with the control group, which may be a result of reduced hydrogen production. However, the amounts of hydrogen produced were not determined in the present study.
The reduction in the population of Butyrivibrio fibrisoívens probably caused a decrease in biohydrogenation of linoleic acid (C18:2n-6), α-linolenic acid (C18:3n-3) and total UFA (C18). This result suggests that MP has the potential to prevent the saturation of UFA and thus increase the presence of UFA in the ruminant meat. To the best of the authors' knowledge, this is the first report on the effect of mangosteen peel on rumen biohydrogenation.
Biohydrogenation in the rumen causes production of CLA isomers and vaccenic acid (C18:1 t11). The c9,t11 isomer of CLA, known as rumenic acid (RA), is the main isomer of CLA in products derived from ruminants. It is considered to have many potential human health-promoting effects (Pariza et al., 2001). RA can be produced by desaturating vaccenic acid in the mammary gland and other tissues. However, it is produced mainly in the rumen through biohydrogenation of dietary linoleic acid (C18:2n-6) by rumen microorganisms, especially Butyrivibrio fibrisolvens The RA is then reduced to vaccenic acid and after that to stearic acid (C18:0) (Palmquist et al., 2005). Accumulation of vaccenic acid in the rumen has been considered a means to increase the content of CLA in ruminant products(Pariza et al., 2001; Palmquist et al., 2005). Inhibition of the rumen biohydrogenation of vaccenic acid to stearic acid leads to accumulation of vaccenic acid in the rumen. Recent studies have reported that this step of rumen biohydrogenation could be inhibited with tannins (Khiaosa-Ard et al., 2009; Vasta et al., 2009a; Vasta et al., 2009b). However, reports on the effects of tannins on rumen biohydrogenation are few and controversial. For instance, in vitro experiments (Khiaosa-Ard et al., 2009; Vasta et al., 2009a) indicate positive effects of tannins on rumen vaccenic acid accumulation, but in vivo studies suggest that tannins have negative effects or none (Benchaar & Chouinard, 2009; Cabiddu et al., 2009; Vasta et al., 2009b). In the present study, although the amounts of vaccenic acid significantly increased by raising the levels of MP, which indicated inhibition of biohydrogenation of vaccenic acid to stearic acid (C18:0), there were no significant differences among the three treatment groups in terms of the amounts of CLA isomers produced.
The pH of rumen fluid is a good indicator of overall rumen function (Kay, 1983). The pH of rumen can range from about 8 to less than 5, based on the diet fed to ruminants. Lower pH values are normally observed in animals receiving concentrate diets (Slyter et al., 1964; Cerrato-Sánchez et al., 2008). Rumen microflora is sensitive to the rumen pH (Istasse et al., 1986). Therefore, any change in rumen pH may lead to huge impact on ruminal microbial composition and the digestive functions (Orskov, 2012). At a lower pH, VFA production is often reduced and acetic acid accumulation is increased (Dunlop & Hammond, 1965). In the present study, the MP had no adverse effect on pH of rumen fluid and VFA production.
Ammonia results from amino acid deamination in the rumen, and tannins can bind to the proteins in the rumen and form insoluble complexes, and therefore reduce protein degradation, and decrease ammonia production in rumen fluid (Patra & Saxena, 2011). This would decrease nitrogen excretion in urine (Carulla et al., 2005), which is responsible for emissions of nitrous oxide, a potent greenhouse gas. Reduction of ammonia production by tannins have been reported for example by Hassanat and Benchaar (2013), who showed that tannin treatments reduced ammonia concentrations, indicating a reduction in ruminal protein degradation. However, in the present study, MP had no significant effect on ammonia production. This may be because different plant sources provide different types, structure and concentration of secondary metabolites.
Conclusion
In terms of total gas and VFA production, MP was comparable with alfalfa, with the advantage that MP can reduce gas production numerically per unit of VFA produced. In addition, MP reduced methane production, possibly by lowering the populations of total protozoa and total methanogens in comparison with alfalfa. Furthermore, MP reduced the percentage of biohydrogenation of UFA, probably by decreasing the population of Butyrivibrio fibrisolvens in comparison with alfalfa. Hence, the results suggest that the MP has great potential to be used in animal feed to reduce ruminal methane production and biohydrogenation, without adverse effects on ruminal pH and VFA production. The effects of MP on ruminal parameters of various ruminant hosts in vivo needs to be investigated in future.
Acknowledgements
The authors are grateful to the Faculty of Veterinary Medicine and Institute of Tropical Agriculture, Universiti Putra Malaysia. This research was supported by Malaysian Government E-Science Grant No. 05-01-04-SF0200.
Authors' Contributions
PS, MAR, YMG, II, MFR, ME and MFJ designed and carried out the studies, performed the statistical analysis, interpreted the results and drafted the manuscript. MAR, ME and PS supervised the study, designed the experiments and drafted the manuscript. MFJ, II, MFR, PS and ME participated in the biochemical and microbial data analysis. ME, II and MFR carried out the in vitro experiment. ME and YMG performed the investigation of fatty acid changes and statistical analysis. All authors read and approved the final manuscript.
Conflict of Interest Declaration
Authors declare that there is no conflict of interest for this study.
References
Animut, G., Puchala, R., Goetsch, A., Patra, A., Sahlu, T., Varel, V. & Wells, J., 2008. Methane emission by goats consuming diets with different levels of condensed tannins from lespedeza. Anim. Feed Sci. Tech. 144, 212-227. [ Links ]
Benchaar, C. & Chouinard, P., 2009. Assessment of the potential of cinnamaldehyde, condensed tannins, and saponins to modify milk fatty acid composition of dairy cows. J. Dairy Sci. 92, 3392-3396. [ Links ]
Bhatta, R., Malik, P.K. & Prasad, C.S., 2015. Enteric Methane Emission: status, mitigation and future challenges-an Indian perspective. In: Livestock Production and Climate Change. Eds Malik, P.K., Bhatta, R., Takahashi, J., Kohn, R. & Prasad, C.S., CABI International, pp. 229-244. [ Links ]
Buccioni, A., Cappucci, A. & Mele, M., 2015. Methane emission from enteric fermentation: methanogenesis and fermentation. In: Climate Change Impact on Livestock: Adaptation and Mitigation. Eds Sejian, V., Gaughan, J., Baumgard, L. & Prasad, C., Springer India, pp. 171-186. [ Links ]
Cabiddu, A., Molle, G., Decandia, M., Spada, S., Fiori, M., Piredda, G. & Addis, M., 2009. Responses to condensed tannins of flowering sulla (Hedysarum coronarium L.) grazed by dairy sheep: Part 2: Effects on milk fatty acid profile. Livest. Sci. 123, 230-240. [ Links ]
Carulla, J., Kreuzer, M., Machmüller, A. & Hess, H., 2005. Supplementation of Acacia mearnsii tannins decreases methanogenesis and urinary nitrogen in forage-fed sheep. Crop Pasture Sci. 56, 961 -970. [ Links ]
Cerrato-Sánchez, M., Calsamiglia, S. & Ferret, A., 2008. Effect of the magnitude of the decrease of rumen pH on rumen fermentation in a dual-flow continuous culture system. J. Anim. Sci. 86, 378-383. [ Links ]
Cieslak, A., Zmora, P., Pers-Kamczyc, E. & Szumacher-Strabel, M., 2012. Effects of tannins source (Vaccinium vitis idaea L.) on rumen microbial fermentation in vivo. Anim. Feed Sci. Tech. 176, 102106. [ Links ]
Crutzen, P.J., Aselmann, I. & Seiler, W., 1986. Methane production by domestic animals, wild ruminants, other herbivorous fauna, and humans. Tellus B. 38, 271 -284. [ Links ]
Dunlop, R.H. & Hammond, P.B., 1965. D-Lactic acidosis of ruminants. Ann. N. Y. Acad. Sci. 119, 1109-1132. [ Links ]
Ebrahimi, M., Rajion, M., Goh, Y. & Sazili, A., 2012. Impact of different inclusion levels of oil palm (Elaeis guineensis Jacq.) fronds on fatty acid profiles of goat muscles. J. Anim. Physiol. Anim. Nutr. 96, 962969. [ Links ]
ˇEbrahimi, M., Rajion, M.A., Goh, Y.M., Farjam, A.S., Sazili, A.Q. & Schonewille, J.T., 2014. The effects of adding lactic acid bacteria and cellulase in oil palm (Elais guineensis Jacq.) frond silages on fermentation quality, chemical composition and in vitro digestibility. Ital. J. Anim. Sci. 13, 557-562. [ Links ]
Ebrahimi, M., Rajion, M.A., Goh, Y.M., Shokryzadan, P., Awis, Q.S. & Jahromi, M.F., 2015. Feeding oil palm (Elaeis guineensis, Jacq.) fronds alters rumen protozoal population and ruminal fermentation pattern in goats Ital. J. Anim. Sci. 14, 403-409. [ Links ]
FAO, 2010. Greenhouse gas emissions from the dairy sector. Food and Agriculture Organization. http://www.fao.org/docrep/012/k7930e/k7930e00.pdf. [ Links ]
Faseleh Jahromi, M., Liang, J.B., Mohamad, R., Goh, Y.M., Shokryazdan, P. & Ho, Y.W., 2013. Lovastatin- enriched rice straw enhances biomass quality and suppresses ruminal methanogenesis. Biomed Res. Int. [serial online]. Available from http://dx.doi.org/10.1155/2013/397934, [ Links ]
Fievez, V., Babayemi, O. & Demeyer, D., 2005. Estimation of direct and indirect gas production in syringes: A tool to estimate short chain fatty acid production that requires minimal laboratory facilities. Anim. Feed Sci. Tech. 123, 197-210. [ Links ]
Folch, J., Lees, M. & Sloane-Stanley, G.H., 1957. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226, 497-509. [ Links ]
Frandson, R.D., Wilke, W.L. & Fails, A.D. 2009: Anatomy and physiology of farm animals. John Wiley & Sons, Colorado, USA. [ Links ]
Getachew, G., Makkar, H. & Becker, K., 2000. Tannins in tropical browses: Effects on in vitro microbial fermentation and microbial protein synthesis in media containing different amounts of nitrogen. J. Agr. Food Chem. 48, 3581-3588. [ Links ]
Guo, Y., Liu, J.X., Lu, Y., Zhu, W., Denman, S. & McSweeney, C., 2008. Effect of tea saponin on methanogenesis, microbial community structure and expression of mcrA gene, in cultures of rumen micro-organisms. Lett. Appl. Microbiol. 47, 421-426. [ Links ]
Hassanat, F. & Benchaar, C., 2013. Assessment of the effect of condensed (acacia and quebracho) and hydrolysable (chestnut and valonea) tannins on rumen fermentation and methane production in vitro. J. Sci. Food Agr. 93, 332-339. [ Links ]
Hess, H.D., Kreuzer, M., Diaz, T.E., Lascano, C.E., Carulla, J.E., Soliva, C.R. & Machmüller, A., 2003. Saponin rich tropical fruits affect fermentation and methanogenesis in faunated and defaunated rumen fluid. Anim. Feed Sci. Tech. 109, 79-94. [ Links ]
Hobson, P.N. (1997): The Rumen Microbial Ecosystem. Chapman and Hall, London, UK. [ Links ]
Istasse, L., Reid, G., Tait, C. & Orskov, E., 1986. Concentrates for dairy cows: effects of feeding method, proportion in diet and type. Anim. Feed Sci. Tech. 15, 167-182. [ Links ]
Jayanegara, A., Goel, G., Makkar, H.P.S. & Becker, K. 2010. Reduction in methane emissions from ruminants by plant secondary metabolites: effects of polyphenols and saponins, In: Odongo N.E., M. garcia & G.J. Viljoen (ed.) Sustainable improvement of animal production and health. Food and Agriculture Organization, Rome, Italy, 151 -157. [ Links ]
Jayanegara, A., Kreuzer, M., Wina, E. & Leiber, F., 2011. Significance of phenolic compounds in tropical forages for the ruminal bypass of polyunsaturated fatty acids and the appearance of biohydrogenation intermediates as examined in vitro. Animal Production Science. 51, 1127-1136. [ Links ]
Kay, R., 1983. Rumen function and physiology. Vet. Rec. 113, 6-9. [ Links ]
Khiaosa-Ard, R., Bryner, S., Scheeder, M., Wettstein, H.-R., Leiber, F., Kreuzer, M. & Soliva, C., 2009. Evidence for the inhibition of the terminal step of ruminal α-linolenic acid biohydrogenation by condensed tannins. J. Dairy Sci. 92, 177-188. [ Links ]
Lu, C.D. & Jorgensen, N.A., 1987. Alfalfa saponins affect site and extent of nutrient digestion in ruminants. J. Nutr. 117, 919-927. [ Links ]
Makkar, H., Norvsambuu, T., Lkhagvatseren, S. & Becker, K., 2009. Plant secondary metabolites in some medicinal plants of Mongolia used for enhancing animal health and production. Tropicultura. 27, 159167. [ Links ]
Makkar, H.P.S., Siddhuraju, S., Siddhuraju, P. & Becker, K. 200: Plant secondary metabolites. Humana Press: Totowa, New Jersey, USA, [ Links ]
McDonald, P., Edwards, R.A., Greenhalgh, J.F.D. & Morgan, C.A. 2002: Animal nutrition. Pearson Education Ltd, Harlow, UK. [ Links ]
Morimoto, K.C., Van Eenennaam, A.L., DePeters, E.J. & Medrano, J.F., 2005. Hot topic: Endogenous production of n-3 and n-6 fatty acids in mammalian cells. J. Dairy Sci. 88, 1142-1146. [ Links ]
Navidshad, B., Liang, J.B. & Jahromi, M.F., 2012. Correlation coefficients between different methods of expressing bacterial quantification using real-time PCR. Int. J. Mol. Sci. 13, 2119-2132. [ Links ]
Ngamsaeng, A. & Wanapat, M., 2005. Effects of mangosteen peel (Garcinia mangostana) supplementation on rumen ecology, microbial protein synthesis, digestibility and voluntary feed intake in beef steers. http://www.mekarn.org/msc2003-05/theses05/mod_p2.pdf. [ Links ]
OJEU, 2003. Regulation (EC) No. 1831/2003 of European Parliament and the Council of 22 September 2003 on additives for use in animal nutrition. Off. J. Eur. Union. Page L268/36 in OJEU of 10/18/2003. Brussels, Belgium. [ Links ]
Orskov, E.R. (2012): Energy nutrition in ruminants. Springer Science & Business Media, New York, USA. [ Links ]
Palmquist, D.L., Lock, A.L., Shingfield, K.J. & Bauman, D.E., 2005. Biosynthesis of conjugated linoleic acid in ruminants and humans. Adv Food Nutr Res. 50, 179-217. [ Links ]
Pariza, M.W., Park, Y. & Cook, M.E., 2001. The biologically active isomers of conjugated linoleic acid. Prog. Lipid Res. 40, 283-298. [ Links ]
Patra, A.K. & Saxena, J., 2011. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food Agr. 91, 24-37. [ Links ]
Porter, L.J., Hrstich, L.N. & Chan, B.G., 1985. The conversion of procyanidins and prodelphinidins to cyanidin and delphinidin. Phytochemistry. 25, 223-230. [ Links ]
Rajion, M.A., McLean, J.G. & Cahill, R.N.P., 1985. Essential fatty acids in the fetal and newborn lamb. Aust. J. Biol. Sci. 38, 33-40. [ Links ]
Rajion, M., Goh, Y., Dahlan, I. & Salam Abdullah, A., 2001. Dietary manipulation and increase in plasma unsaturated fatty acids in sheep. Asian Australas. J. Anim. Sci. 14, 1073-1077. [ Links ]
SAS, 2008. Statistical Analysis System. SAS online documentation 9.2. SAS Institute Incorporation. [ Links ]
Slyter, L., Nelson, W. & Wolin, M., 1964. Modifications of a device for maintenance of the rumen microbial population in continuous culture. Appl. Microbiol. 12, 374-377. [ Links ]
Solorzano, L., 1969. Determination of ammonia in natural waters by the phenolhypochlorite method. Limnol. Oceanogr. 14, 799-801. [ Links ]
Szumacher-Strabel, M. & Cieslak, A. 2012. Dietary possibilities to mitigate rumen methane and ammonia production, In: Liu G. (ed.) Greenhouse Gases Capturing, Utilization and Reduction. Intech, Rijeka, Croatia. Intech, Rijeka, Croatia, 199-238. [ Links ]
Tan, H.Y., Sieo, C.C., Abdullah, N., Liang, J.B., Huang, X.D. & Ho, Y.W., 2011. Effects of condensed tannins from Leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro. Anim. Feed Sci. Tech. 169, 185-193. [ Links ]
Tavendale, M.H., Meagher, L.P., Pacheco, D., Walker, N., Attwood, G.T. & Sivakumaran, S., 2005. Methane production from in vitro rumen incubations with Lotus pedunculatus and Medicago sativa, and effects of extractable condensed tannin fractions on methanogenesis. Anim. Feed Sci. Tech. 123, 403-419. [ Links ]
Theodoridou, K., Aufrère, J., Niderkorn, V., Andueza, D., Le Morvan, A., Picard, F. & Baumont, R., 2011. In vitro study of the effects of condensed tannins in sainfoin on the digestive process in the rumen at two vegetation cycles. Anim. Feed Sci. Tech. 170, 147-159. [ Links ]
US Environmental Protection Agency, 2007. Ruminant livestock. How much methane is produced by livestock. http://www.epa.gov/rlep/faq.html. [ Links ]
Vasta, V., Makkar, H.P.S., Mele, M. & Priolo, A., 2009a. Ruminal biohydrogenation as affected by tannins in vitro. Brit. J. Nutr. 102, 82-92. [ Links ]
Vasta, V., Mele, M., Serra, A., Scerra, M., Luciano, G., Lanza, M. & Priolo, A., 2009b. Metabolic fate of fatty acids involved in ruminal biohydrogenation in sheep fed concentrate or herbage with or without tannins. J. Anim. Sci. 87, 2674. [ Links ]
Wang, Y., McAllister, T., Yanke, L. & Cheeke, P., 2000. Effect of steroidal saponin from Yucca schidigera extract on ruminal microbes. J. Appl. Microbiol. 88, 887-896. [ Links ]
Received 10 September 2015
Accepted 11 September 2016
First published online 24 November 2016
# Corresponding authors: mebrahimii@gmail.com; ymgoh@upm.edu.my


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}