










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.46 n.4 Pretoria 2016
http://dx.doi.org/10.4314/sajas.v46i4.2
Comparison of shear force tenderness, drip and cooking loss, and ultimate muscle pH of the loin muscle among grass-fed steers of four major beef crosses slaughtered in Namibia
P. StrydomI, II; J. LühlI; C. KahlI; L.C. HoffmanI, #
IDepartment of Animal Sciences, Stellenbosch University, P Bag X1 Matieland, 7602. South Africa
IIAgricultural Research Council - Animal Production Institute, Private Bag X2, IRENE, 0062. South Africa
ABSTRACT
The authors investigated the effects of breed (Brahman, Bonsmara, Simbrah and Simmental crosses) and post-mortem ageing on meat tenderness, purge, cooking loss and ultimate pH of the longissimus muscle under commercial production and slaughter conditions of the Namibian beef export industry. Fifty steers of each breed cross, at an average age of 2½ years and with an average subcutaneous fat cover of 2 mm over the 9th-10th thoracic vertebrae, were included in this study. The Brahman (BRX) differed significantly (P < 0.05) from all other breed crosses in all ageing treatments, recording higher Warner-Bratzler shear force values. The Bonsmara (BNX) steers showed the highest rate of tenderization and maintained this advantage up to day 30 post mortem. Purge and cooking loss increased initially and then tapered off over ageing time. BNX steers recorded the highest cooking loss.
Keywords: beef, breeds, cooking loss, grass-fed, purge, tenderness
Introduction
The eating quality of beef is best described as a combination of flavour, juiciness and tenderness (Shackelford et al., 1995; Koohmaraie et al., 2003; Verbeke et al., 2010). However, various studies have found that inconsistent tenderness is the most important factor in determining consumer dissatisfaction (Brooks et al., 2000; Koohmaraie et al., 2003; Chambaz et al. 2003; Hanzelkoá et al., 2011).
Free-range beef production in Namibia poses multiple challenges in terms of climate, parasites and predators, to name a few. For this reason, resilient breeds and breed crosses that are able to produce and reproduce under these conditions are used extensively. The four main breed crosses (Bonsmara, Brahman, Simbrah, and Simmental) that are slaughtered at Windhoek Export Abattoir of the Meat Corporation of Namibia (MEATCO) generally reflect these requirements. The Bonsmara, as a locally developed composite breed (composite B. taurus africanus), Brahman (B. indicus) and Simbrah (B. indicus taurus composite) breeds are well adapted to the sub-tropical conditions in Namibia. Although the Simmental breed (B. taurus) is considered less adapted in the tropics, it is the oldest farmed cattle breed in Namibia after the indigenous Sanga animals (B. taurus africanus). The first Simmental animals were introduced to Namibia in 1893 (Anonymous, 2009a) and have been adapted to local environmental challenges for more than a century.
Although meat tenderness and other meat quality characteristics depend on a number of inherent factors (e.g. breed, age, sex, muscle type, marbling) and environmental factors (e.g. nutrition, ante-mortem stress conditions, slaughter processes and chilling conditions, and ageing), research has shown that the production advantages of B. indicus and B. indicus x B. taurus composite breeds in harsher environments are partially negated by problems with beef tenderness and other quality characteristics (Strydom, 2008; Ferguson et al., 2000; Jerez-Timaure & Huerta-Leidenz, 2009).
Namibia is a net exporter of beef, with South Africa, European Union, and Japan being its major markets (Bowles et al., 2005), while Norway was recently added to the list. All these markets are discerning about the quality of the beef that they import. Because Namibia has a small feedlot industry, most of the export beef originates from older (2-3 years) animals that are finished on natural pastures, which may result in tougher beef than that of younger feedlot finished animals. In addition, because of variations in the duration of shipping and cold storage for the various markets and particular client specifications, meat may be exposed to post-mortem ageing periods of five weeks or more. All these factors may affect the quality of the final product.
The aim of this investigation was to measure and compare the meat quality characteristics of the loin muscle of the four most prominent beef breeds farmed extensively on natural pasture for the Namibian export market. In addition, the study investigated the effect of extended post mortem ageing on tenderness and other quality parameters in order to overcome possible quality defects caused by breed and production system conditions.
Materials and Methods
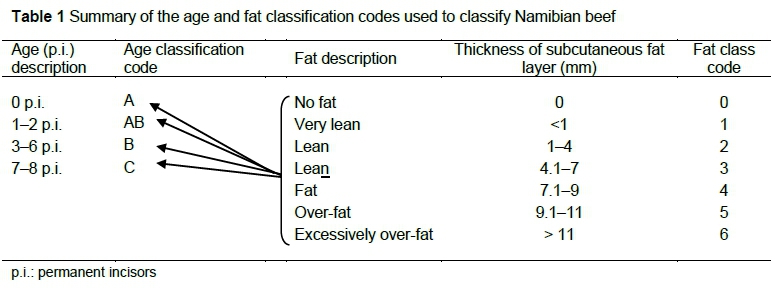
Namibian carcass classification is similar to the South African classification system and is based mainly on differentiation of age, according to dentition, carcass fatness and conformation, determined subjectively (Anon 1999b). Table 1 presents a summary of the South African and Namibian beef classification system with regards to age and fatness. Fifty steers of each of the four major breed crosses slaughtered in Namibia, Bonsmara (BNX), Brahman (BRX), Simbrah (SBX), and Simmental (SX), were selected in the lairage of the export Windhoek MEATCO Abattoir. After slaughter only carcasses of age class B (2-2½ years old) (Lawrence et al., 2001) with a fat code of 2 (1-4 mm subcutaneous fat) were included in this study.
Animals originated from various production areas in Namibia as well as different producers. All animals in this study were raised on natural pasture.
The animals were killed according to the standard slaughter procedures of the export abattoir, that is, stunning by captive bolt, hoisting by the hind leg, dead weight determination, exsanguination, dressing, evisceration, carcass classification, hot weight determination and being placed in the cooler (4 °C). No electrical stimulation was applied.
After 48 hours the carcasses were removed from the coolers and deboned. A portion of the longissimus muscle (LD) was removed from the three-rib cut of the left side of identified carcasses, cut into six 15-mm thick steaks, which were then vacuum-packed individually. The weight of each steak was recorded as well as the pH at 48 hours post mortem (pHu). The pH was determined with a hand-held pH meter (Testo-205, Scientific Laboratories Supplies, UK). Steaks were randomly allocated to various ageing treatments (2, 9, 16, 23, 30 and 37 days of ageing) packed in boxes and stored at 4 °C until they were evaluated.
These physical measurements were carried out on the individual steaks:
• Purge: Purge refers to weep/water accumulating in vacuum bag of packed meat over time. The individually packed steaks were removed from their bags after the relevant ageing period, gently dried between absorbent tissues, and weighed. Percentage purge was calculated as the weight loss expressed as a percentage of the original sample weight as demonstrated by Strydom & Hope-Jones (2014).
• Cooking loss: The steaks used to determine purge were weighed and then heated in sealed plastic bags in a water bath at 80 °C for 60 min. The cooked samples were cooled down under running water to room temperature (ca. 25 °C), dried and weighed again. Cooking loss was calculated as total fluid lost and collected in the bag during cooking, expressed as percentage of the uncooked sample weight (adapted from Honikel, 1998).
• Warner Bratzler shear force (WBSF) (mechanical measurement of tenderness) was performed on the steaks that had been used to determine cooking loss. Five cylindrical samples (12.7-mm core diameter) were cored parallel to the grain of the meat, making sure that the whole steak area was covered. Each of the five cores were sheared perpendicular to the fibre direction using a Warner-Bratzler shear attachment (with a circular cross-section of 1.27 cm 0 blade) fitted to an electrical scale programmed to measure maximum weight/resistance. The reported value in kilograms represented the average of the peak force measurements of each of the five cores (Hoffman et al., 2009). For further analysis, samples were categorized into three tenderness groups based on their WBSF values according to threshold levels reported by Shackelford et al. (1991), Voisinet et al. (1997), and Shackelford et al. (1999): 'tender' (< 3.9 kg), 'intermediate' (3.9-4.6 kg) and 'tough' (> 4.6 kg).
Statistical analysis was done with Proc GLM of SAS Enterprise Guide 3.0 (SAS, 2006). The data of purge loss, cooking loss, pH and Warner Bratzler shear force (WBSF) were subjected to analysis of variance (ANOVA), with post-mortem ageing (2, 9, 16, 23, 30 and 37 days) and breed (BNX, BRX, SBX, and SX) as the main effects. Means were separated with Bonferroni's multiple comparison method to determine least significant differences (LSD) at 5% level (Snedecor & Cochran, 1980).
Results
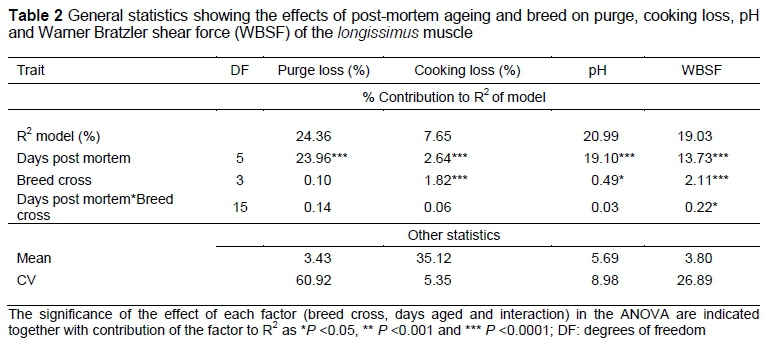
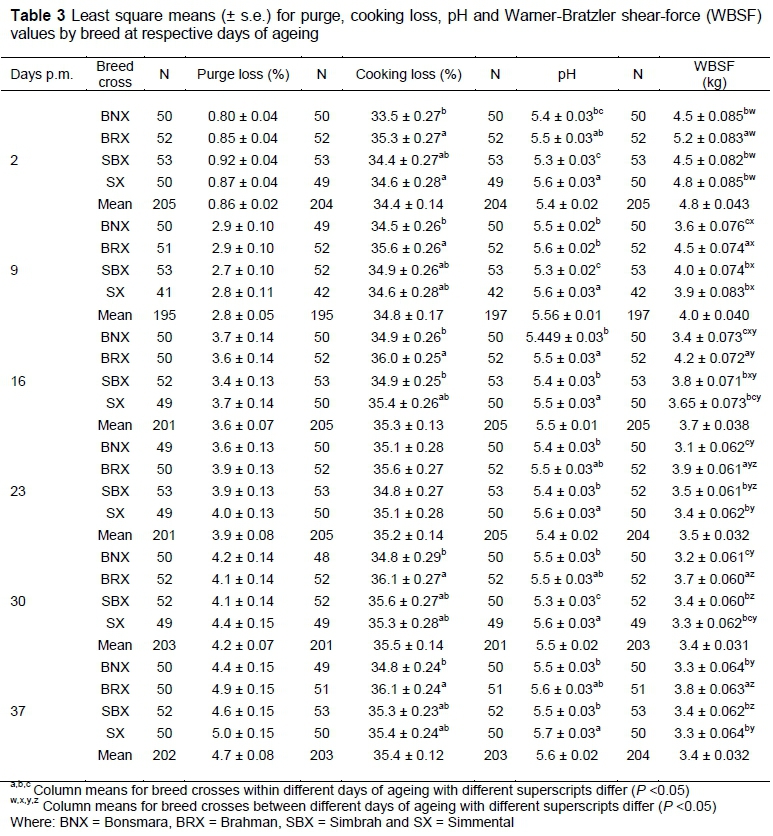
General statistics for the four measured quality traits are presented in Table 2. Least square means for these traits are summarized in Table 3 with superscripts identifying means, which differed significantly among breed crosses within and between individual days of ageing.
Although breed cross had no influence on purge, it had a significant influence on pH and a highly significant influence on cooking loss and WBSF. Days post mortem had a highly significant influence on all four quality traits.
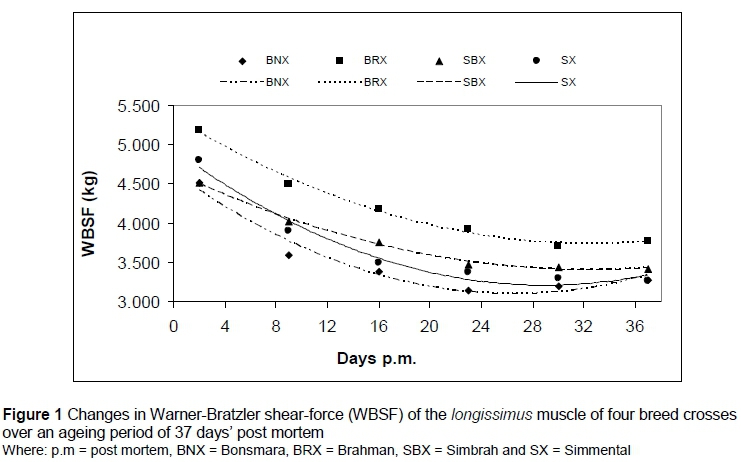
A significant interaction was recorded for WBSF (P <0.05) between breed cross and days post mortem. As expected, WBSF values decreased over the ageing period, reaching equilibrium around 23 days (Figure 1). BRX recorded consistently higher WBSF values compared with the other three breed crosses for the duration of the ageing period. At two days, no differences were found between BNX, SBX and SX, but between 9 and 30 days, BNX recorded lower WBSF values than the other groups (Table 3). In addition, SBX recorded numerically lower WBSF values on day 2 post mortem compared with SX. This was reversed at 16 days' post mortem, indicating that the rate of post-mortem tenderization was higher in the SX.
The amount of purge increased significantly in a curvilinear way over ageing time (Table 3), with most of the losses occurring between 2 and 9 days. A similar trend was observed for cooking loss, although the effect was less pronounced and no differences occurred between 16 and 36 days. However, in contrast to purge, there were consistent breed differences for cooking loss throughout the ageing period, although the magnitude of the differences were small. BNX recorded lower cooking losses (P <0.05) than BRX throughout the ageing period, except on day 23, while the other two breeds showed values intermediate to BRX and BNX. BRX recorded higher pHu values (i.e. after rigor mortis) compared with the other three breeds at two days' post mortem. This difference was maintained throughout the ageing period. pH values initially declined slightly and increased again between 30 and 37 days' post mortem.
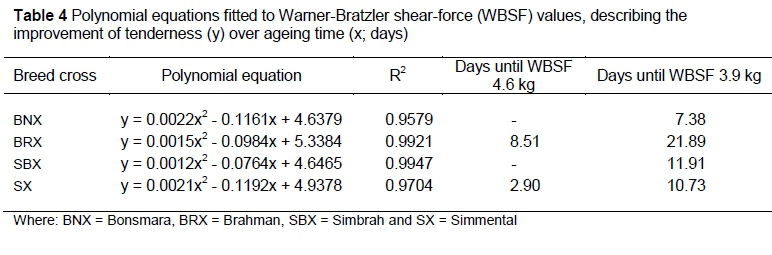
The data depicted in Figure 1 were fitted with polynomial models. The resulting equations (Table 4) were used to calculate the days post mortem to reach 'intermediate' (4.6 kg) and 'tender' (3.9 kg) WBSF values on average. BNX and SBX reached 'intermediate' WBSF values before day 2, while SX reached these values around 3 days' post mortem and the BRX around 9 days. It took the BNX, BRX, SBX and SX 7, 22, 12 and 11 days of ageing, respectively, to reach 'tender' WBSF values.
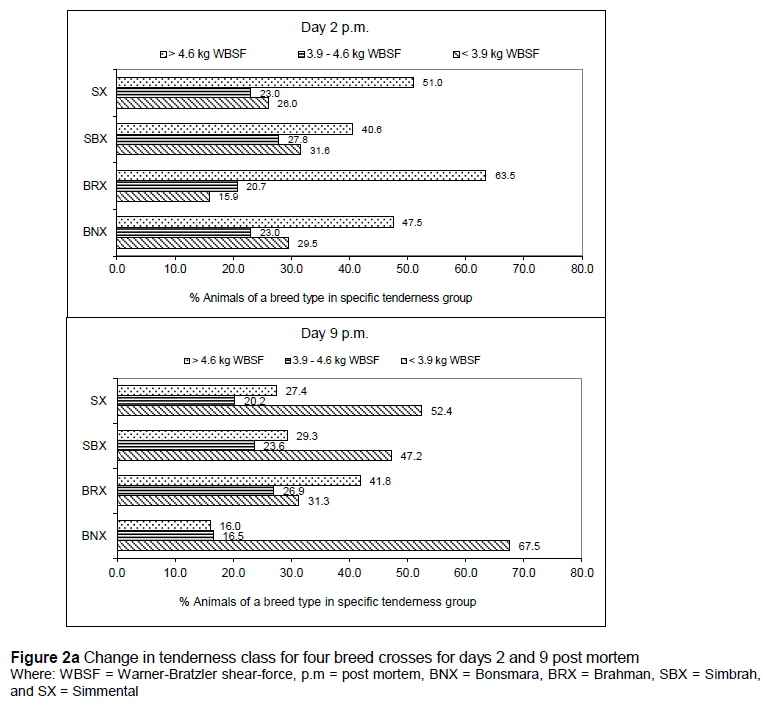
The changes in tenderness class within breed crosses over the ageing period of 37 days are shown in Figure 2a and b. The most noticeable change took place between days 2 and 9 post mortem with the percentage of animals moving from 'tough' (> 4.6 kg) to 'intermediate' (3.9-4.6 kg) and 'tender' (< 3.9 kg). Furthermore, the rate of tenderizing of the BNX group was much higher compared with the other breed crosses. Tenderness improved up to 30 days' post mortem and then stabilised.
Discussion
Juiciness is one of the three sensory attributes that determine overall eating satisfaction of meat (Shackelford et al., 1995; Aaslyng et al., 2003), the others being tenderness and flavour. The amount of water captured and retained in meat before and after cooking influences its juiciness and therefore palatability, as well as the saleable weight of the product. Water is lost from the meat in the form of purge during storage and in thawing of frozen meat, and during cooking through evaporation and drip loss (stock) (Den Hertog-Meischke et al., 1998; Aaslyng et al., 2003). Water exists in muscle in three forms, namely water bound to proteins (this fraction is small, is not affected by freezing and is not lost during conventional cooking); entrapped water (held by steric effects and/or by attraction to bound water), and free water (Huff-Lonergan & Lonergan, 2005). The fraction that is most affected by the conversion of muscle to meat or rigor is the entrapped water. During the rigor process, muscle pH decreases, mainly because of the anaerobic conversion of glycogen to lactic acid. The pH ultimately reaches the iso-electric point of the main muscle proteins (myosin: pH = 5.4), reducing the net charge of these proteins to zero. This means that the attraction of water is reduced, and the repulsion of structures within myofibrils is diminished, allowing the components to move nearer to one another (lateral shrinkage) (Huff-Lonergan & Lonergan, 2005; Pearce et al., 2011). This effectively reduces the space available for water, and the water manifests as drip or purge (purge loss describes the water that is visible in vacuum-packed meat). Low pH leads to partial denaturation of the myosin head, which could contribute to shrinkage of the lateral space. These changes take place soon after death, leading to relatively high levels of purge over the first days post mortem.
There were differences in pHu (pH after rigor mortis was reached) among the breed groups, with pHu of SX being 0.3 units (day 2) above the iso-electric point of meat proteins. That of SBX is closest to the isoelectronic point. However, since the values were in the range for normal meat (pH 5.4 to 5.7) (Renerre, 1990), there was probably little effect on water-holding capacity and therefore on purge, which agrees with the results of Muchenje, et al. (2008) and Du Plessis & Hoffman (2007) for pasture-raised animals. Contradictory results were reported by Jama et al. (2008), who found a decrease in drip loss over an ageing time of 21 days. Despite significant breed cross effects being recorded for cooking loss over the duration of ageing, the actual differences were at the most 1.5 percentage units. BRX cuts showed the highest cooking losses for the duration of the ageing period. According to Huff-Lonergan & Lonergan (2005), impaired postmortem ageing contributes to lateral shrinkage of the muscle cells and creates drip channels, in which more moisture flows into extracellular space and increases moisture losses. Because the BRX group recorded the toughest meat throughout the ageing period, it is plausible that this led to increased moisture losses during cooking, although it is surprising that this effect did not increase the amount of purge as well. Increased purge and cooking loss over time of ageing were expected (Table 3). Kristensen & Purslow (2001) and Straadt et al. (2007) noted that there is initially a continuous release of water from meat over time during ageing and this may stabilise, as in our study. Since cytoskeletal protein forms 10% of muscle protein (Labeit & Kolmerer, 1995), water is released continuously as the meat ages as a result of cytoskeletal structure degradation.
Connective tissue content, sarcomere length and myofibrilar breakdown are the primary sources of meat tenderness variation. Connective tissue components are believed to be responsible for the variation observed in background toughness, in particular the organization of the perimysium (Strandine et al.,1949; Koohmaraie & Geesink, 2006). Background toughness was defined by Marsh & Leet (1966) as 'the resistance to shearing of the muscle before shortening occurred' and is a factor that exists at slaughter and does not change during ageing (Koohmaraie & Geesink, 2006). These effects are more prominent in high connective tissue cuts, and probably less so in the longissimus muscle (Rhee et al., 2004)
The toughening phase starts with rigor mortis development and is caused by sarcomere shortening (Koohmaraie et al., 1996; Hanzelková et al., 2011). This phase is similar in carcasses handled under similar processing conditions (Luciano et al., 2007), as in the current study. The tenderizing phase starts directly after slaughter and may vary greatly in rate and extent among breeds, in particular where B. indicus is involved. This variation causes the inconsistency in tenderness observed at consumer level (Koohmaraie & Geesink, 2006). Proteolysis of key myofibrilar and myofibrilar-associated proteins forms the basis of the tenderizing phase (Koohmaraie & Geesink, 2006; Muchenje et al., 2009). The primary proteolytic enzyme system involved in post mortem tenderization of aged beef is the calpain system, which consists of two calcium requiring enzymes, μ-calpain and m-calpain, as well as an inhibitor, calpastatin (Koohmaraie, 1996; Ibrahim et al., 2008). Other factors such as temperature, pH and calcium ion concentration can influence this process (Takahashi, 1996).
The difference in tenderness between B. indicus and B. taurus is well documented. It appears that it is mainly the result of calpastatin activity in post-mortem muscle. The lower tenderness observed in the BRX (Figure 1; Table.2) is in accordance with the findings of Crouse et al. (1989), Whipple et al. (1990), Johnson et al. (1990a,b), O'Connor et al. (1997), Pringle et al., 1997), Voisinet et al. (1997), Ferguson et al. (2000) and Ibrahim et al. (2008). O'Connor et al. (1997) and Pringle et al. (1997) measured calpastatin activity and concluded that the increased calpastatin activity found in B. indicus breeds would be the major reason for the higher WBSF of BRX group at three days and the lower extent of ageing over the subsequent 34 days (Figure 1). Similar effects between breed x ageing time were reported by Johnson et al. (1990b). Since the animals in this study were of commercial origin, the percentage of indicus blood was not known and identification of crosses was based on phenotypes. O'Connor et al. (1997) and Ibrahim et al. (2008) found that biological type interacted with ageing and reported that as the percentage Brahman inheritance increased, the rate and extent of tenderization decreased. Johnson et al. (1990b) showed that differences in tenderness became significant only when the percentage Brahman breeding reached fifty per cent, while Stolowski et al. (2006) found that calpastatin levels were highest in the longissimus muscle in animals with 50% B. indicus breeding. As well as from the significant effect of indicus influence on WBSF of BRX group, SBX cuts were less tender than BNX at each stage of ageing, except for the final WBSF on day 37. The magnitude of the differences however, was less than between BNX and BRX. Of further interest is the higher WBSF of SX animals compared with BNX at 2, 9 and 23 days (Table 3). SX and BNX did not differ at any stage during ageing. In agreement with these trends, Frylinck & Heinze (2003) reported higher WBSF values for pure Brahman and Simmental compared with Hereford, Afrikaner, Bonsmara and Nguni animals. These differences were supported by higher calpastatin levels in Brahman cuts compared with the other more tender breeds, but higher WBSF was not reflected in the calpastatin levels of the Simmental cuts.
The Bonsmara breed is made up of 5/8 Afrikaner and 3/8 Hereford and Shorthorn. It might be because of its B. taurus inheritance that this breed cross showed the lowest shear values throughout the trial period. Strydom et al. (2000) reported favourable tenderness results for B. taurus africanus breeds, such as the Afrikaner, and therefore the influence of this breed contributed to the tenderness results of the BNX group. Most comparisons of tenderness between B. indicus and B. taurus breeds named here were done on grain-fed younger cattle, while the current study showed similar trends for older grass-fed animals. Du Plessis & Hoffman (2007) reported numerically higher WBSF values for loin cuts of grass-fed Simmental crosses of between 0.6 and 0.9 kg compared with Nguni and Bonsmara crosses.
The display of WBSF distribution in the three tenderness categories in Figures 2a and b confirms that the BNX produced the best quality meat of all the breed crosses investigated. Although the SX and SBX groups lagged behind the BNX in terms of number of samples in the 'tender' class during the initial stages of ageing, the differences were diluted as ageing progressed. However, considering the cost involved in storage of meat, using the BNX group means that meat develops an acceptable level of tenderness earlier than meat from SBX and SX breed crosses and much earlier than meat from BRX. SBX will probably lag behind SX in this regard, while both groups will take fewer ageing days to produce acceptable levels of tenderness than the BRX group. Even after 37 days, between 7% and 22% (i.e. 4 to 11) of the 50 or so samples within a group still measured more than 4.6 kg WBSF.
Conclusion
Clear breed differences were found for cooking loss and tenderness among the four main breed crosses slaughtered in Namibia. Tenderness differences between breeds are in accordance with previous research, performed mostly under grain-fed systems. Ageing clearly improved tenderness in all breeds. Under the conditions of this study, it seems that BNX animals produce loin muscle cuts that age faster and therefore reach acceptable levels of tenderness sooner than the other breeds, while the loins of BRX animals take the longest.
Acknowledgments
The help of the staff from the abattoir in Windhoek is appreciated. This work is based on research supported by the South African Research Chairs Initiative of the Department of Science and Technology and the National Research Foundation of South Africa (Grant: 84633). Opinions, findings and conclusions or recommendations expressed in this material are those of the author(s) and the National Research Foundation does not accept liability.
Authors' Contributions
Conception and design - LCH; Data collection, analyses and drafting of paper - JL; Critical revision - PS & CK; critical revision and final approval of version to be published - LCH.
Conflict of Interest Declaration
The authors certify that they have no affiliations with or involvement in any organization or entity with any financial or non-financial interest in the subject matter or materials discussed in this manuscript.
References
Anonymous, 2009a. History. The Simmentaler/Simbra Cattle Breeders' Society of Southern Africa. http://simmentaler.org/Simmentaler-History.htm. [ Links ]
Anonymous, 2009b. Government notice No. R.342 of March 1999. Regulations regarding the classification and marketing of meat. Government Gazzette of the Republic of South Africa. http://www.gov.za/sites/www.gov.za/files/38431_reg10358_gon55.pdf. [ Links ]
Aaslyng, M.D., Bejerholm, C., Ertbjerg, P., Bertram, H.C. & Andersen, H.J., 2003. Cooking loss and juiciness of pork in relation to raw meat quality and cooking procedures. Fd Qual. Pref. 14, 277-288. [ Links ]
Bowles, D., Paskin, R., Gutierrez, M. & Kasterine, A., 2005. Animal welfare and developing countries: opportunities for trade in high welfare products from developing countries. Rev. Scient. Techn. Off. Intern. Épizo. 24, 783-790. [ Links ]
Brooks, J.C., Belew, J.B., Griffin, D.B., Gwartney, B.L., Hale., D.S., Henning, W.R., Johnson, D.D., Morgan, J.B., Parrish, Jr.,F.C., Reagan J.O. & Savell, J.W., 2000. National beef tenderness survey - 1998. J. Anim. Sci. 78, 1852-1860. [ Links ]
Chambaz, A., Scheeder, M., Kreuzer, M. & Dufey, P., 2003. Meat quality of Angus, Simmental, Charolais and Limousin steers compared at the same intramuscular fat content. Meat Sci. 63, 491-500. [ Links ]
Crouse, J.D., Cundiff, L. V., Koch, R.M., Koohmaraie, M. & Seideman, S.C., 1989. Comparisons of Bos indicus and Bos taurus inheritance for carcass beef characteristics and meat palatability. J. Anim. Sci. 67, 2661-2668. [ Links ]
Den Hertog-Meischke, M.J.A., Smulders, F.J.M. & van Logtestijn, J.G., 1998. The effect of storage temperature on drip loss from fresh beef. J. Sci. Fd Agric. 78, 522-526. [ Links ]
Du Plessis, I., Hoffman, L.C., 2007. Effect of slaughter age and breed on the carcass traits and meat quality of beef steers finished on natural pastures in the arid subtropics of South Africa. S. Afr. J. Anim. Sci. 37, 143-153. [ Links ]
Ferguson, D.M., Jiang, S.-T., Hearnshaw, H., Rymill, S.R. & Thompson, J.M., 2000. Effect of electrical stimulation on protease activity and tenderness of M. longissimus from cattle with different proportions of Bos indicus content. Meat Sci. 55, 265-272. [ Links ]
Frylinck, L. & Heinze, P.H. 2003. Evaluation of meat tenderness of indigenous South African and other beef breeds. In Consistency of quality. Proceedings of the 11th International Meat Symposium, Centurion, South Africa. (pp. 3-13). [ Links ]
Hanzelková, Š, Simeonovová, J., Hampel, D., Dufek, A. & Šubrt, J., 2011. The effect of breed, sex and ageing time on tenderness of beef meat. Acta Veter. Brno 80, 191-196. [ Links ]
Hoffman, L.C., Mostert, A.C., Kidd, M. & Laubscher, L.L., 2009. Meat quality of kudu (Tragelaphus strepsiceros) and impala (Aepyceros melampus): carcass yield, physical quality and chemical composition of kudu and impala Longissimus dorsi muscle as affected by gender and age. Meat Sci. 83, 788-795. [ Links ]
Honikel, K.O., 1998. Reference methods for the assessment of physical characteristics of meat. Meat Sci. 49, 447-457. [ Links ]
Huff-Lonergan, E & Lonergan, S.M., 2005. Mechanisms of water-holding capacity of meat: The role of postmortem biochemical and structural changes. Meat Sci. 71, 194-204. [ Links ]
Ibrahim, R., Goll, D., Marchello, J., Duff, G., Thompson, V., Mares, S. & Ahmad, H., 2008. Effect of two dietary concentrate levels on tenderness, calpain and calpastatin activities, and carcass merit in Waguli and Brahman steers. J. Anim. Sci. 86, 1426-1433. [ Links ]
Jama, N., Muchunje, V., Chimonyo, M., Strydom, P.E., Dzama, K. & Raats, J.G., 2008. Cooking loss components of beef from Nguni, Bonsmara and Angus steers. Afr. J. Agric. Res. 3, 416-420. [ Links ]
Jerez-Timaure, N. & Huerta-Leidenz, N., 2009. Effects of breed type and supplementation during grazing on carcass traits and meat quality of bulls fattened on improved savannah. Livest. Sci. 121, 219-226. [ Links ]
Johnson, M.H., Calkins, C.R., Huffman, R.D., Johnson, D.D. & Hargrove, D.D., 1990-b. Differences in cathepsin B + L and calcium-dependent protease activities among breed type and their relationship to beef tenderness. J. Anim. Sci. 68, 2371-2379. [ Links ]
Johnson, D.D., Huffman, R.D., Williams, S.E. & Hargrove, D.D., 1990-a. Effects of percentage Brahman and Angus breeding, age-season of feeding and slaughter end point on meat palatability and muscle characteristics. J. Anim. Sci. 68, 1980-1986. [ Links ]
Koohmaraie, M. & Geesink, G.H., 2006. Contribution of post-mortem muscle biochemistry to the delivery of consistent meat quality with particular focus on the calpain system. Meat Sci. 74, 34-43. [ Links ]
Koohmaraie, M., 1996. Biochemical factors regulating the toughening and tenderization processes of meat. Meat Sci. 43, 193-201. [ Links ]
Koohmaraie, M., Doumit, M.E. & Wheeler, T.L., 1996. Meat toughening does not occur when rigor shortening is prevented. J. Anim. Sci. 74, 2935-2942. [ Links ]
Koohmaraie, M., Veiseth, E., Kent, M., Shackleford, S. & Wheeler, T., 2003. Understanding and managing variation in tenderness. In: Consistency of quality: abstracts and proceedings of the 11th International Meat Symposium, Centurion, South Africa, ARC, 3-13. [ Links ]
Kristensen, L. & Purslow, P., 2001. The effect of ageing on the water-holding capacity of pork: the role of cytoskeletal proteins. Meat Sci. 58, 17-23. [ Links ]
Labeit, S. & Kolmerer, B., 1995. Titins: giant proteins in charge of muscle ultrastructure and elasticity. Science, 270, 236. [ Links ]
Lawrence, T.E., Whatley, J.D., Montgomery, T.H. & Perino, L.J., 2001. A comparison of the USDA ossification-based maturity system to a system based on dentition. J. Anim. Sci. 79, 1683-1690. [ Links ]
Luciano, F.B., Antón, A.A. & Rosa, C.F., 2007. Biochemical aspects of meat tenderness: a brief review. Rev. Arch. Zoot. 56 (R), 1-8. [ Links ]
Marsh, B.B. & Leet, N.G., 1966. Studies in meat tenderness. III. The effect of cold shortening on tenderness. J. Fd Sci.31, 450-459. [ Links ]
Muchenje, V., Dzama, K., Chimonyo, M., Raats, J.G. & Strydom, P.E., 2008. Meat quality of Nguni, Bonsmara and Aberdeen Angus steers raised on natural pasture in the Eastern Cape, South Africa. Meat Sci. 79, 20-28. [ Links ]
Muchenje, V., Dzama, K., Chimonyo, M., Strydom, P.E., Hugo, A. & Raats, J.G., 2009. Some biochemical aspects pertaining to beef eating quality and consumer health: A review. Fd Chem. 112, 279-289. [ Links ]
O'Connor, S.F., Tatum, J.D., Wulf, D.M., Green, R.D. & Smith, G.C., 1997. Genetic effects on beef tenderness in Bos indicus composite and Bos taurus cattle. J. Anim. Sci. 75, 1822-1830. [ Links ]
Pearce, K.L., Rosenvold, K., Andersen, H.J. & Hopkins, D.L., 2011. Water distribution and mobility in meat during the conversion of muscle to meat and ageing and the impacts on fresh meat quality attributes-A review. Meat Sci. 89, 111-124. [ Links ]
Pringle, T.D., Williams, S.E., Lamb, B.S., Johnson, D.D. & West, R.L., 1997. Carcass characteristics, the calpain proteinase system, and aged tenderness of Angus and Brahman crossbred steers. J. Anim. Sci. 75, 2955-2961. [ Links ]
Renerre, M. 1990. Factors involved in the discoloration of beef meat. Int. J. Fd Sci. Techn., 25, 613-630. [ Links ]
Rhee, M.S., Wheeler, T.L., Shackelford, S.D. & Khoomaraie, M. 2004. Variation in palatability and biochemical traits within and among eleven beef muscles. J. Anim. Sci. 82, 534-550. [ Links ]
SAS, 2006. SAS for Windows version 9.1.3. [ Links ]
Shackelford, S.D., Morgan, J.B., Cross, H.R. & Savell, J.W., 1991. Identification of threshold levels for Warner-Bratzler shear force in beef top loin steaks. J. Musc. Fd. 2, 289-296. [ Links ]
Shackelford, S.D., Wheeler, T.L. & Koohmaraie, M., 1995. Relationship between shear force and trained sensory panel tenderness ratings of ten major muscles from Bos indicus and Bos taurus cattle. J. Anim. Sci. 73, 3333-3340. [ Links ]
Shackelford, S., Wheeler, T. & Koohmaraie, M., 1999. Tenderness classification of beef: II. Design and analysis of a system to measure beef longissimus shear force under commercial processing conditions. J. Anim. Sci. 77, 1474-1481. [ Links ]
Snedecor, G. W. & Cochran, W. G. 1980. Statistical methods (7th ed.). Iowa State University Press, 507. [ Links ]
Stolowski, G.D., Baird, B.E., Miller, R.K., Savell, J.W., Sams, A.R., Taylor. J.F., Sanders, J.O., Smith, S.B., 2006. Factors influencing the variation in tenderness of seven major beef muscles from three Angus and Brahman breed crosses. Meat Sci. 73, 475-483. [ Links ]
Straadt, I. K., Rasmussen, M., Andersen, H. J. & Bertram, H. C., 2007. Aging-induced changes in microstructure and water distribution in fresh and cooked pork in relation to water-holding capacity and cooking loss - A combined confocal laser scanning microscopy (CLSM) and low-field nuclear magnetic resonance relaxation study. Meat Sci. 75, 687-695. [ Links ]
Strandine, E.J., Koonz, C.H. & Ramsbottom, J.M., 1949. A study of variations in muscles of beef and chicken. J. Anim. Sci. 8, 483-494. [ Links ]
Strydom, P.E., Naude, R.T., Smith, M.F., Scholtz, M.M. & van Wyk, J.B., 2000. Characterisation of indigenous African cattle breeds in relation to meat quality traits. Meat Sci. 55, 79-88. [ Links ]
Strydom, P.E., 2008. Do indigenous Southern African cattle breeds have the right genetics for commercial production of quality meat. Meat Sci. 80, 86-93. [ Links ]
Strydom, P.E. & Hope-Jones, M. 2014. Evaluation of three vacuum packaging methods for retail beef loin cuts. Meat Sci. 98, 689-694 [ Links ]
Takahashi, K., 1996. Structural weakening of skeletal muscle tissue during post-mortem ageing of meat: the non-enzymatic mechanisms of meat tenderization. Meat Sci. 43, 67-80. [ Links ]
Verbeke, W., Pérez-Cueto, F.J., de Barcellos, M.D., Krystallis, A. & Grunert, K.G., 2010. European citizen and consumer attitudes and preferences regarding beef and pork. Meat Sci. 84, 284-292. [ Links ]
Voisinet, B.D., Grandin, T., O'Connor, S.F., Tatum, J.D. & Deesing, M.J., 1997. Bos indicus-cross feedlot cattle with excitable temperaments have tougher meat and a higher incidence of borderline dark cutters. Meat Sci. 46, 367-377. [ Links ]
Whipple, G., Koohmaraie, M., Dikeman, M.E., Crouse, J.D., Hunt, M.C. & Klemm, R.D., 1990. Evaluation of attributes that affect longissimus muscle tenderness in Bos taurus and Bos indicus cattle. J. Anim. Sci. 68, 2716-2728. [ Links ]
Received 8 May 2015
Accepted 30 August 2016
Published online 27 October 2016
#Corresponding author: lch@sun.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}