










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.46 n.3 Pretoria 2016
http://dx.doi.org/10.4314/sajas.v46i3.8
Interactions of genotype, housing and dietary calcium in layer performance, eggshell quality and tibia characteristics
E. TůmováI, # ; J. VlčkováI; V. CharvátováI; O. DrábekII; V. TejneckýII; M. KettaI; D. ChodováI
IDepartment of Animal Husbandry, Czech University of Life Sciences Prague, Kamýcká 129, 165 21 Prague 6 Suchdol, Czech Republic
IIDepartment of Soil Science and Soil Protection, Czech University of Life Sciences Prague, 165 21 Prague 6 Suchdol, Czech Republic
ABSTRACT
The study was designed to evaluate the interactions in performance, eggshell quality and tibia traits of two laying hen genotypes, namely a commercial hybrid, Lohmann (LSL), and a traditional breed, the Czech Hen (CH), housed in conventional cages and on litter, and fed two levels of dietary calcium (Ca) (3.5% vs. 3.0%). A significant interaction between genotype, housing and Ca was determined in egg weight. The heaviest eggs were laid by LSL hens housed in cages and fed 3.0% Ca (61.8 g), and the lightest laid by CH, also in cages on 3.0% Ca (47.3 g). Eggshell strength and shell weight were significantly affected by three-way interaction. Czech hens kept in cages and fed 3.0% Ca produced the strongest shells (4480 g/cm2), whereas the CH in cages on 3.5% Ca had the weakest (3665 g/cm2) shells. Hens housed on litter had significantly stronger tibias than hens housed in cages (485 vs. 397 N). With respect to genotype, tibias were stronger in CH. Housing and level Ca in feed had a significant effect on tibia weight. However, none of these factors affected tibia thickness or ash content. Tibia Ca content was higher in caged birds than in those on litter. The results of the study show that interaction between housing, genotype and Ca resulted in large differences in measurements of egg weight and eggshell quality. Tibia characteristics were less affected by the treatments, and no interactions were evident in the variables.
Keywords: Cage, calcium level, Czech Hen, laying hens, litter, Lohmann
Introduction
Commercially available laying hen genotypes have been selected over many decades for egg production and egg size. This selection has resulted in differences between modern commercial hybrids and traditional pure breeds. On the other hand, traditional breeds have adapted to environmental conditions of the region of their origin and may therefore be suitable in alternative housing systems. There are limited studies in which traditional breeds and commercial hybrids have been compared. Hocking et al. (2003) observed that commercial lines displayed greater rates and persistency of lay, and laid larger eggs, which contained more albumen of higher quality than traditional breeds. In contrast, eggshell weight and thickness have not changed over time. Anderson et al. (2004) did not find differences in eggshell thickness, percentage or specific gravity between historic strains and commercial laying stock. This correlates with findings by Hocking et al. (2003), who reported that there were no differences in eggshell percentage between commercial strains and pure breeds, but a greater eggshell weight was found in the commercial lines. Commercial lines had weak bones with relatively little genetic variation for eggshell strength compared with traditional lines.
Genetic improvement in egg production with good eggshell quality has increased Ca requirements (Farmer et al., 1986). However, little attention has been paid to the Ca requirement in traditional breeds that could be used in alternative production systems. Traditional breeds have often been used in organic production (Rizzi & Chiericato, 2010; Rizzi & Marangon, 2012), but little is known about the quality of their product and their requirements.
The effect of housing systems on eggshell quality is not uniform, and there is no clear indication of which production system results in eggs with the best shell quality (Holt et al., 2011). Inconsistent results could be explained through the interaction of housing system and genotype (Leyendecker et al., 2001; Van den Brand et al., 2004; Singh et al., 2009; Tůmová et al., 2011). Housing system affects bone characteristics, with reports that hens kept in cages have weaker bones than those in alternative housing (Abrahamsson & Tauson, 1997; Leyendecker et al., 2001; Lichovníková & Zeman, 2008). Leyendecker et al. (2001) found a negative correlation between eggshell stability and bone strength. Again, there is lack of data on housing system and bone thickness and strength in traditional breeds.
Calcium is the major structural element in the eggshell, and plays an important role in medullary bone formation. The medullary bone is a reserve of Ca for eggshell formation and a common indicator of Ca metabolism in laying hens. The prevalence of broken bones is related to the high requirement for Ca deposition in eggshells (Hocking et al., 2003). The tibia is known to contain marked amounts of medullary bone (Leyendecker et al., 2005). This bone is turned over according to a daily egg cycle and contributes 35% to 40% of the Ca for eggshell formation (Kim et al., 2012). Calcium requirements can also depend on the genotype of laying hens. The NRC (1994) recommends a dietary Ca content of 3.25% for Leghorn type and 3.6% for brown laying hens. However, there are no data for traditional breeds. It is possible to assume that traditional breeds have lower Ca requirements than hybrids owing to their lower egg production and higher feed intake. Feed Ca level affects bone strength. Cheng & Coon (1990) observed a linear relationship between increasing Ca intake from 2.0 g/d to 4.5 g/d and bone breaking strength. Similarly, Nascimento et al. (2014) found a beneficial effect of increasing dietary Ca (2.85% - 5.25%) on tibia breaking strength. Swiatkiewicz et al. (2015) demonstrated a linear decrease in relative Ca retention with increasing dietary Ca, and confirmed that at low dietary Ca concentration, layers are able to maximize their utilization of dietary Ca. Cufadar et al. (2011) and Neijat et al. (2011) suggested that housing system should also be taken into account when determining the correct Ca requirements in the diets of laying hens. Lichovníková & Zeman (2008) observed that Ca intake was similar in enriched cages and on the floor system. However, a lower proportion of Ca was deposited in the eggshells on the floor system and in enriched cages compared with conventional cages. Tibia strength was also higher on the floor system.
The aim of the present study was to evaluate interactions in performance, eggshell quality and tibia traits of two laying hen genotypes (a commercial hybrid, the Lohmann (LSL) and a traditional breed, the Czech Hen (CH)), housed in conventional cages and on litter, and fed two dietary Ca levels (3.5% and 3.0%).
Materials and Methods
The experiment was approved by the Ethics Committee of the Czech University of Life Sciences Prague and the Central Commission for Animal Welfare at the Ministry of Agriculture of the Czech Republic.
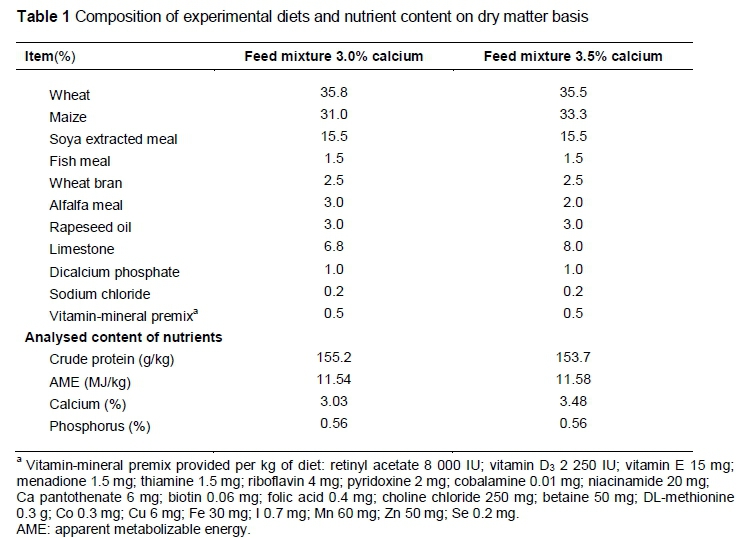
The experiment with 126 laying hens, LSL, and 126 of the traditional breed, CH, was carried out in the second half of the laying cycle from 40 to 60 weeks old. In the first phase of the laying cycle, hens were kept under the same conditions. The CH is the original layer breed that was developed in the middle of the nineteenth century in the Czech Republic. Their live weight is 1.8 kg to 2.2 kg, while egg yield and egg weight (58 g on average) are low, with late sexual maturity. The hens have partridge-coloured plumage, a white skin and white eggshells. The breed is in the conservation programme of genetic resources of the Czech Republic. Laying hens were housed in conventional Eurovent cages (550 cm2/hen, 3 hens in a cage, 22 cages per genotype, 11 cages per group) and in 12 littered pens with floor space of 7 hens/m (six pens for each genotype, three pens per group). Laying hens in both housing systems were fed commercial feed mixtures, differing only in Ca content, namely 3.5% and 3.0%. The composition of the feed mixtures is given in Table 1. Feed and water were available ad libitum throughout the experiment. In the diet, dry matter was determined by drying samples at 105 °C to a constant weight. AOAC International procedures (2005) were used to determine the crude protein (954.01) and ash (942.05) contents. Gross energy was determined by combustion with an adiabatic bomb calorimeter (Laget MS10A, Laget, CZ). Calcium content was analysed using the AOAC method 965.17, based on vanad-molybden reagent and spectrophotometry analysis on Solaar M6 apparatus (TJA Solutions, Cambridge, UK). Environmental conditions were in accordance with laying hen requirements. The daily photoperiod consisted of 15 h light and 9 h darkness (lights were turned on at 4:00 and off at 19:00). During the experiment, egg production and feed consumption were recorded daily. Data of egg collection were used to calculate hen-day egg production (§ekeroglu et al., 2014) and expressed as egg production (%). Based on feed intake, daily Ca consumption was calculated.
Eggs for quality assessment were collected on two consecutive days in a four-week interval. The first collection was at 40 weeks old. All eggs in each cage or littered pen were collected, but defective eggs such as those with double yolks or abnormal shells were excluded (total 1320 eggs). The eggs were weighed individually, and shell strength was determined by the shell-breaking method using a QC-SPA device (TSS York, UK) on the equatorial region. Shell weight was determined after three days of drying at room temperature. The eggshell proportion was calculated from the egg weight and eggshell weight. The eggshell weight was determined after drying. Eggshell thickness was calculated as the mean of three measurements in the equatorial region, after removing the eggshell membranes, using a QCT shell thickness micrometer (TSS York, UK). Eggshell density was calculated according to the formula eggshell density = (shell weight/shell surface) x 100 (Ahmed et al., 2005) and the surface area of each egg was calculated using the equation suggested by Thomson et al. (1985), namely egg surface = 4.67 (egg weight)2/3.
At 50 weeks old, six birds per group were randomly selected to determine tibia characteristics. The selected hens were slaughtered by cervical dislocation after electric stunning. After slaughtering, both tibia were removed completely from the carcass and cleaned of connective tissues. Weight, length, strength and thickness were evaluated on the right tibia. Tibia strength was measured with a QC-SPA device (TSS York, UK) and thickness with a QCT micrometer (TSS York, UK). Ash and Ca were analysed using the left tibia. The tibia bones were incinerated (500 °C) and crushed. From each sample, 0.5 g was weighed and 5 mL 35% HCl (p.a. Penta s.r.o., CZ) was added with 10 gtt 65% HNO3 (p.a. Penta s.r.o., CZ). The mixture was heated for 15 min. and subsequently diluted 200 times, using re-distilled water. Samples prior to analysis were further diluted and acidified with 2 M HNO3 and lanthanized (La). Concentration of La in each sample was adjusted to 10 mg/mL. Ca content was determined by FAAS (AA280FS Varian, Australia) under standard conditions. Data quality was controlled and assured by standard laboratory procedures, including blanks and certified reference material.
The results of the experiment were evaluated with ANOVA, using the GLM procedure of SAS (SAS, 2013). The model included the effects of genotype, housing system and feed Ca level, as well as the interactions among them. A P-value of P <0.05 was considered significant for all measurements.
Results
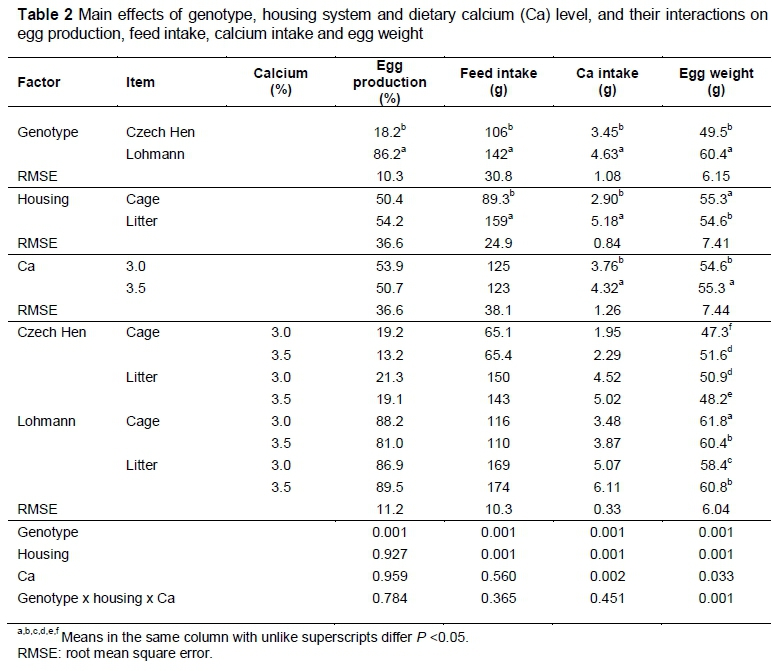
The main effects of laying hen genotype, housing system, feed Ca level, their interactions on egg production, feed intake, Ca intake and egg weight are presented in Table 2. The results show that all production parameters for CH were lower compared with LSL (P <0.001). Neither housing system nor Ca level in the feed had an effect on the rate of lay. In the cage system, feed intake (p <0.001) and Ca intake (P <0.001) were lower, but egg weight was higher (p <0.001). The higher Ca level increased both Ca intake (p <0.002) and egg weight (p <0.033).
There was no interaction between genotype, housing and Ca level (p >0.05) in rate of lay, feed or Ca intake. However, a three-way interaction was observed in egg weight (p <0.001). The heaviest eggs were laid by LSL hens housed in cages and fed 3% Ca, while the lightest eggs were laid by CH, also in cages and on 3% Ca. The two genotypes reacted differently, depending on the housing and Ca level. An increase in dietary Ca resulted in an increase in egg weight in CH housed in cages and in LSL housed on the floor, whereas this increase in Ca caused egg weight to fall in CH housed on the floor and in LSL hens in cages.
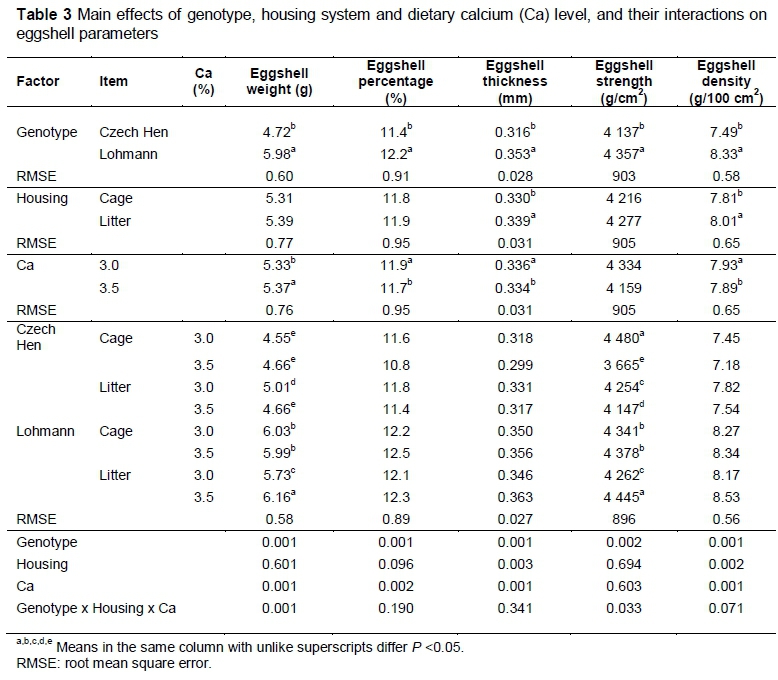
Eggshell parameters for weight, percentage, thickness, strength and density (Table 3) were higher (P <0.001) in eggs laid by the LSL hens. Eggshell thickness (p <0.003) and density (p <0.002) were higher for hens kept on litter. Higher dietary Ca content increased eggshell weight (p <0.001) while decreasing shell percentage (p <0.002), thickness (p <0.002) and shell density (p <0.001). Eggs with the heaviest shells were produced by LSL hens kept on litter and fed 3.5% Ca. There was a significant interaction between genotype, housing and Ca level in shell weight. In contrast, the strongest shells were produced by CH hens housed in cages and fed 3.0% Ca and in LSL hens kept on litter and fed 3.5% Ca. The eggshell strength was affected by an interaction (p <0.033) of genotype, housing and feed Ca level.
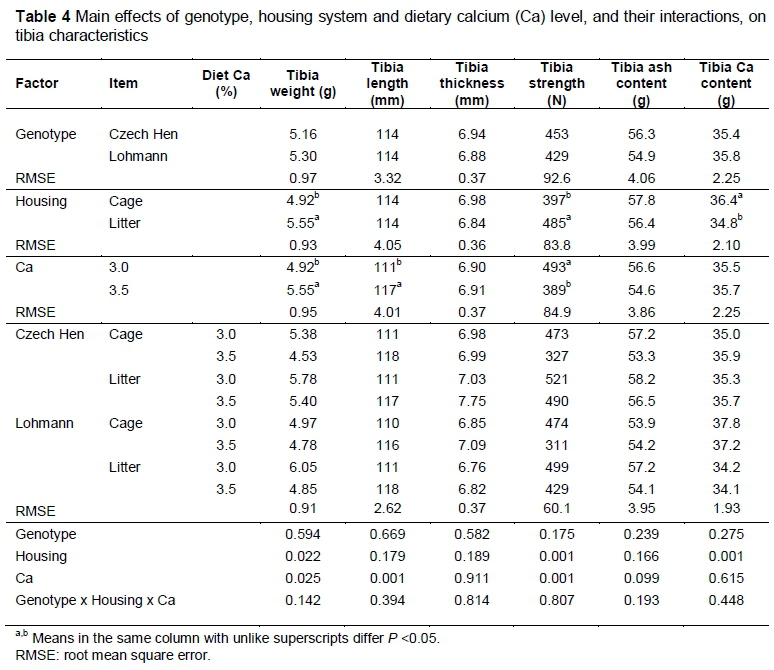
The results of the tibia characteristics indicate that individual factors play an important role in tibia structure (Table 4). Genotype did not affect any of the tibia measurements significantly (P >0.05), whereas laying hens kept on litter had heavier and stronger tibias, but with a lower Ca content. An increased feed Ca level resulted in heavier (P <0.025) and longer (P <0.001) tibias. Tibia strength was lower (P <0.001) on the higher dietary Ca level, whereas tibia ash and Ca content were not affected. There were no significant interactions (P >0.05) among the factors.
Discussion
The results of the study confirmed differences in all production parameters between LSL and CH hens. It is evident that intensive selection for egg production in LSL hens has resulted in higher rates of lay and egg size, which corresponds with the observations of Hocking et al. (2003), Krawczyk (2009) and Rizzi & Chiericato (2010). Housing system had a considerable effect on feed and Ca intake as well as egg weight. In cages, the FI of CH was 57% that consumed by LSL hens in the same housing system, while the difference in FI on litter between genotypes was 8%. The lower FI of CH compared with LSL could be related to their lower productivity. This corresponds with results by Rizzi & Chiericato (2010). The extremely low FI by CH in cages could be the result of this breed's discomfort in cages. Data also indicate that the low FI in CH in cages reduced egg production, but did not influence eggshell or tibia parameters. Feed Ca level did not affect egg production, which is in agreement with the results of Bar et al. (2002), Castillo et al. (2004) and Cufadar et al. (2011), but is in contrast with those of Narváez-Solarte et al. (2006) and Safaa et al. (2008).
These inconsistent results can be related to variable genotypes, age and Ca levels in these studies. Increasing feed Ca level did not affect feed intake, but significantly increased Ca intake. The results indicate that laying hens can tolerate high levels of Ca, as observed by Keshavarz & Nakijama (1993). The higher Ca level resulted in increased egg weight, which is in contrast with the results of Castillo et al. (2004), Valkonen et al. (2010) and Cufadar et al. (2011), who reported that Ca level had no effect on egg weight. The difference in results could be because of the significant interaction between genotype, housing and feed Ca level, all of which appeared to play an important role in this trait. The CH hens produced heavier eggs on 3.5% Ca in cages, whereas on litter the heaviest eggs were produced on 3.0% Ca. Presumably, in the CH genotype housed in cages, a higher dietary Ca level compensated for the low feed intake (3.85 vs. 3.48 g Ca/d). However, the combination of high Ca content and a high feed intake in hens on litter resulted in an additional 0.5 g Ca intake per day (4.52 - 5.02 g). This presumably led to a reduction in the ingestion of other nutrients, resulting in lower egg weight. This theory corresponds with that of Bar et al. (2002), who observed that increasing Ca intake from 3.6 to 5.02 g decreased egg weight. LSL hens in cages compensated for the lower feed Ca level by increasing feed intake, which corresponds with the findings of Narváez-Solarte et al. (2006). On the other hand, on litter, a higher feed intake balanced out lack of other nutrients, which affected egg weight more than rate of lay.
For all the eggshell characteristics, higher values were observed in LSL, which can be attributed to a result of long-term selection for these characteristics. This corresponds with the results of Anderson et al. (2004), Krawczyk (2009), and Rizzi & Marangon (2012), who reported that commercial genotypes have better eggshell quality than traditional breeds. However, Hocking et al. (2003) observed relatively little genetic variation in eggshell strength, and no detectable differences between breeds in hens that were 55 weeks old. Our study evaluated eggshell strength over a longer period, between the 40th and 60th week of age. Housing system did not affect many of the eggshell quality measurements. No differences were found in eggshell weight, eggshell percentage or strength between eggs from the two housing systems, namely cages and litter, which corroborates the work of Tumová et al. (2011) and Englmaierová et al. (2014). However, thicker shells and higher shell density were observed on litter. Yan et al. (2014) observed that eggs with thicker, but less uniform shells resulted in lower shell strength. This relationship is assumed to be the result of correlations between eggshell traits. Low correlations exist between eggshell weight and strength or thickness and strength (Rodriguez-Navarro et al., 2002; Tumová & Ledvinka, 2009). Blanco et al. (2014) reported correlations between eggshell strength and thickness (0.50 - 0.63) and stated that these moderate relationships demonstrate that the strength of an egg does not rely on shell thickness alone, but on the quality and uniformity of eggshell construction. The results in the present study indicate that 3.5% feed Ca resulted in a Ca intake of 4.32 g/d, which is too high to be beneficial to eggshell quality, corresponding with the findings of Safaa et al. (2008). In addition, a significant interaction was found between genotype, housing and Ca in eggshell weight and strength. This interaction plays an important role mainly in eggshell strength, which is a function of shell structural integrity.
The tibia measurements were unaffected by genotype or the interactions between genotype, housing and Ca level. On the contrary, Hocking et al. (2003) observed that 55-week-old commercial layers had lighter, weaker and less stiff tibias than traditional breeds. These results are in contrast with their following study (Hocking et al., 2009), in which no differences in tibia quality were found between commercial layers and traditional breeds. The discrepancy between the two studies could not be explained. The lack of difference in tibia characteristics between genotypes in the present study indicates similar Ca metabolism in the two genotypes. This corresponds with Hocking et al. (2003), who suggested that the absence of differences is likely to be normal developmental consequences of the greater weight of the traditional birds. In hens housed on litter, tibia weight and strength were higher, which corresponds with findings by Abrahamson & Tauson (1997), Leyendecker et al. (2001), Lichovníková & Zeman (2008) that higher values for tibia measurements on litter are associated with greater activity. Lower tibia Ca content indicates that the increased Ca consumption in this housing system presumably resulted in a higher Ca excretion (Swiatkiewicz et al., 2015). The theory can be linked to the Ca intake on litter, which was about 55% higher than in cages. The lower tibia Ca content and higher strength corresponds with results from work by Cufadar et al. (2011), who observed that increasing dietary Ca increased tibia strength, but decreased tibia Ca content. The higher dietary Ca level resulted in higher tibia weight and length, but lower tibia strength, whereas Ca content was not affected. These results support the assumption above because higher dietary Ca increased Ca intake by about 14% compared with lower dietary Ca. Therefore it was not beneficial to tibia strength and did not affect tibia Ca content, which corresponds with the findings of Safaa et al. (2008) and Jiang et al. (2013).
Conclusion
The results of this study show large differences in performance and eggshell quality with respect to genotypes, housing and Ca levels. With regard to the housing system, ultrastructural differences in eggshell structure are assumed. A higher Ca level led to lower bone turnover, which resulted in improved tibia weight, length and thickness, but not in tibia strength. Of particular importance were the significant interactions among these factors in the variables, which need to be considered when determining the way in which these two laying strains are fed.
Acknowledgement
The study was supported by the Ministry of Agriculture of the Czech Republic (Project NAAR No. QJ 1310002).
Authors' Contributions
JV, VCH, MK conducted the experiment as a part of their PhD programme, OD, VT contributed for the tibia analyses, DCH was responsible for the statistical evaluation and supervising of the experiment, ET designed and analysed the experiment and wrote the paper.
Conflict of Interest Declaration
The authors declare that there is no conflict of interests.
References
Abrahamsson, P. & Tauson R., 1997. Effect of group size on performance, health and birds' use of facilities in furnished cages for laying hens. Acta Agric. Scand. Section A-Animal Science. 47, 254-260. [ Links ]
Ahmed, A.M.H., Rodriguez-Navarro, A.B., Vidal, M.L., Gautron, J., Garcia-Ruiz, J.M. & Nys, Y., 2005. Changes in eggshell mechanical properties, crystallographic texture and in matrix proteins induced by moult in hens. Br. Poult. Sci. 46, 268-279. [ Links ]
Anderson, K.E., Tharrington, J.B., Curtis, P.A. & Jones, F.T., 2004. Shell characteristics of eggs from historic strains of single comb white leghorn chickens and the relationship of egg shape to shell strength. Int. J. Poult. Sci. 3, 17-19. [ Links ]
AOAC, 2005. Official Methods of Analysis, 18th ed. Association of Official Analytical. Chemists, Maryland, USA. [ Links ]
Bar, A., Razaphkovsky, V. & Vax, E., 2002. Re-evaluation of calcium and phosphorus requirements in aged laying hens. Br. Poult. Sci. 43, 261-269. [ Links ]
Blanco, A.E., Icken, W., Ould-Ali, D., Cavero, D. & Schmutz, M., 2014. Genetic parameters of egg quality traits on different pedigree layers with special focus on dynamic stiffness. Poult. Sci. 93, 2457-2463. [ Links ]
Castillo, C., Cuca, M., Pro, A., González, M. & Morales, E., 2004. Biological and economic optimum level of calcium in white leghorn laying hens. Poult. Sci. 83, 868-872. [ Links ]
Cheng, T.K. & Coon, C.N., 1990. Effect of calcium source, particle size, limestone solubility in vitro and calcium intake level on layer bone status and performance. Poult. Sci. 69, 2214-2219. [ Links ]
Cufadar, Y., Olgun, O. & Ö. Yildiz, A., 2011. The effect of dietary calcium concentration and particle size on performance, eggshell quality, bone mechanical properties and tibia mineral contents in moulted laying hens. Br. Poult. Sci. 52, 761-768. [ Links ]
Englmaierová, M., Túmová, E., Charvátová, V. & Skřivan, M., 2014. Effects of laying hens housing system on laying performance, egg quality characteristics, and egg microbial contamination. Czech J. Anim. Sci. 59, 345-352. [ Links ]
Farmer, M., Roland, D.A. & Clark, A.J., 1986. Influence of dietary calcium on bone calcium utilization. Poult. Sci. 65, 337-344. [ Links ]
Hocking, P.M., Bain, M., Channing, C.E., Fleming, R. & Wilson, S., 2003. Genetic variation for egg production, egg quality and bone strength in selected and traditional breeds of laying fowl. Br. Poult. Sci. 44, 365-373. [ Links ]
Hocking, P.M., Sandercock, D.A., Wilson, S. & Fleming, R.H., 2009. Quantifying genetic (co) variation and effects of genetic selection on tibial bone morphology and quality in 37 lines of broiler, layer and traditional chickens. Br. Poult. Sci. 50, 443-450. [ Links ]
Holt, P.S., Davies, R.H., Dewulf, J., Gast, R.K., Huwe, J.K., Jones, D.R., Waltman, D. & Willian, K.R., 2011. The impact of different housing systems on egg safety and quality. Poult. Sci. 90, 251-262. [ Links ]
Jiang, S., Cui, L., Shi, CH., Ke, X., Luo, J. & Hou, J., 2013. Effects of dietary energy and calcium levels on performance, eggshell quality and bone metabolism in hens. Vet. J. 198, 252-258. [ Links ]
Keshavarz, K. & Nakajima, S., 1993. Re-evaluation of calcium and phosphorus requirements of laying hens for optimum performance and eggshell quality. Poult. Sci. 72, 144-153. [ Links ]
Kim, W.K., Bloomfield, S.A., Sugiyama, T. & Ricke, S.C., 2012. Concepts and methods for understanding bone metabolism in laying hens. Wrld. Poult. Sci. J. 68, 71-82. [ Links ]
Krawczyk, J., 2009. Effect of layer age and egg production level on changes in quality traits of eggs from hens of conservation breeds and commercial hybrids. Ann. Anim. Sci. 9, 185-193. [ Links ]
Leyendecker, M., Haamann, H., Hartung, J., Kamphues, J., Ring, C., Glünder, G., Ahlers, C., Sander, I., Neumann, U. & Distl, O., 2001. Analysis of genotype-environment interactions between layer lines and hen housing systems for performance traits, egg quality and bone breaking strength -2nd communication: Egg quality traits. Züchtungskunde 73, 290-307. [ Links ]
Leyendecker, M., Haamann, H., Hartung, J., Kamphues, J., Neumann, U., Sürie, C. & Distl, O., 2005. Keeping laying hens in furnished cages and an aviary housing system enhances their bone stability. Br. Poult. Sci. 46, 536-544. [ Links ]
Lichovníková, M. & Zeman, L., 2008. Effect of housing system on the calcium requirement of laying hens and eggshell quality. Czech J. Anim. Sci. 53, 162-168. [ Links ]
Narváez-Solarte, W., Rostagno, H.S., Soares, P.R., Uribe-Velasquez, L.F. & Silva, M.A., 2006. Nutritional requirement of calcium in white laying hens from 46 to 62 wk of age. Int. J. Poult. Sci. 5, 181-184. [ Links ]
Nascimento, G.R., Murakami, A.E., Guerra, A.F.Q.M., Ospinas-Rojas, I.C., Ferreira, M.F.Z. & Fanhani, J.C., 2014. Effect of different vitamin D sources and calcium levels in the diet of layers in the second laying cycle. Braz. J. Poult. Sci. 16, 37-42. [ Links ]
Neijat, M., House, J.D., Guenter, W. & Kebreab, E., 2011. Calcium and phosphorus dynamics in commercial laying hens housed in conventional or enriched cage systems. Poult. Sci. 90, 2383-2396. [ Links ]
NRC, 1994. Nutrient Requirements for Poultry. 9th ed. National Academy Press, Washington, D.C., USA. [ Links ]
Rizzi, C. & Chiericato, G.M., 2010. Chemical composition of meat and egg yolk of hybrid and Italian breed hens reared using an organic production system. Poult. Sci. 89, 1239-1251. [ Links ]
Rizzi, C. & Marangon, A., 2012. Quality of organic eggs of hybrid and Italian breed hens. Poult. Sci. 91, 2330-2340. [ Links ]
Rodriguez-Navarro, A., Kalin, O., Nys, Y. & Garcia-Ruiz, J.M., 2002. Influence of the microstructure on the shell strength of eggs laid by hens of different ages. Br. Poult. Sci. 43, 395-403. [ Links ]
Safaa, H.M., Serrano, M.P., Valencia, D.G., Frikha, M., Jimenéz-Moreno, E. & Mateos, G.G., 2008. Productive performance and egg quality of brown egg-laying hens in the late phase of production as influenced by level and source of calcium in the diet. Poult. Sci. 87, 2043-2051. [ Links ]
SAS, 2013. Statistical Analysis System for Windows. Release 9.4. SAS Institute Inc., Cary, North Carolina, USA. [ Links ]
Şekeroğlu, A., Duman, M., Tahtali, Y., Yildrim, A. & Eleroğlu, H., 2014. Effect of cage tier and age on performance, egg quality and stress parameters of laying hens. S. Afr. J. Anim. Sci. 44, 288-297. [ Links ]
Singh, R., Cheng, K.M. & Silversides, F.G., 2009. Production performance and egg quality of four strains of laying hens kept in conventional cages and floor pens. Poult. Sci. 88, 256-264. [ Links ]
Swiatkiewicz, S., Arczewska-Wlosek, A. & Jozefiak, D., 2015. Bone quality, selected blood variables and mineral retention in laying hens fed with different dietary concentrations and sources of calcium. Livest. Sci. 181, 194-199. [ Links ]
Thomson, B.K., Hamilton, R.M.G. & Grunder, A.A., 1985. The relationship between laboratory measures of eggshell quality and breakage in commercial egg washing and candling equipment. Poult. Sci. 64, 901-909. [ Links ]
Tůmová, E. & Ledvinka, Z., 2009. The effect of time of oviposition and age on egg weight, egg components weight and eggshell quality. Arch. Geflügelkde 72, 110-115. [ Links ]
Tůmová, E., Englmaierová, M., Ledvinka, Z. & Charvátová, V., 2011. Interaction between housing system and genotype in relation to internal and external egg quality parameters. Czech J. Anim. Sci. 56, 490-498. [ Links ]
Valkonen, E., Venäläinen, E., Rossow, L. & Valaja, J., 2010. Effects of calcium diet supplements on egg strength in conventional and furnished cages, and effects of 2 different nest floor materials. Poult. Sci. 89, 2307-2316. [ Links ]
Van den Brand, H., Parmentier, H. & Kemp, K., 2004. Effect of housing system (outdoor vs. cages) and age of laying hens on egg characteristics. Br. Poult. Sci. 45, 745-752. [ Links ]
Yan, Y.Y., Sun, C.J., Lian, L., Zheng, J.X., Xu, G.Y. & Yang, N., 2014. Effect of uniformity of eggshell thickness on eggshell quality in chickens. J. Poult. Sci. 51, 338-342. [ Links ]
Received 3 March 2016
Accepted 31 June 2016
First published online 30 August 2016
# Corresponding author: tumova@af.czu.cz


{kind=link}
{kind=link}
{kind=link}
{kind=link}