










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.46 n.3 Pretoria 2016
http://dx.doi.org/10.4314/sajas.v46i3.3
Fatty acid profile and health lipid indices in the raw milk of ewes grazing part-time and hemp seed supplementation of lactating ewes
D. Mierlita#
University of Oradea, Department of Animal Science, Oradea City, Romania
ABSTRACT
This experiment was designed to evaluate the effects of the feeding system (indoor or part-time grazing) of ewes and dietary supplementation with hemp seed (non-drug varieties of Cannabis sativa L.) on milk production and the composition of milk fatty acids (FAs). The experimental diets were constituted as a 2 x 2 factorial arrangement of feeding system (I: indoors and receiving 1400 g dry matter (DM) grass hay/d or pasture grazing (PG)-4 h/day + 700 g DM grass hay/d), and with or without hemp seed (Hs) supplementation (-Hs = no hemp seed vs. +Hs = 175 g/d hemp seed). All animals received 700 g DM/day of a concentrate for lactation. Feeding the PG diets increased raw milk production and yields of fat, protein and lactose. Feeding +Hs increased fat-corrected milk, milk fat content and fat yield significantly while the combination of +Hs and PG increased fat-corrected milk and fat yield significantly. Milk protein content was not affected by dietary factors. Feeding the I diets increased the proportion of short-chain FAs (C4:0 -C10:0), at the expense of medium-chain FAs (12 to 17 C units). Feeding +Hs decreased the proportion of short-chain FAs, medium-chain FAs and hypercholesterolaemic FA (sum of C12:0, C14:0, C16:0) and increased the trans-vaccenic acid (C18:1 t11, VA), α-linolenic acid (C18:3 n-3, ALA), conjugated linoleic acid (CLA), eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA) concentrations in milk fat significantly. The combination of hemp seed and part-time grazing produced the highest proportion of c9,t11 CLA, t10,c12 CLA and n-3 FA (sum of ALA, EPA, DHA) in milk fat. The polyunsaturated fatty acid (PUFA)/saturated fatty acid (SFA) ratios in the milk from ewes grazing part time were significantly higher, whereas the atherogenicity index (AI), thrombogenic index (TI) and linoleic acid (LA)/ALA ratio were significantly lower compared with the milk of indoor-fed ewes. Supplementing with +Hs has a similar effect of increasing the PUFA/SFA ratio and lowering of the AI, TI and LA/ALA ratio. It was concluded that pasture grazing combined with hemp seed had a synergistic effect on milk production and fat yield without altering the milk protein content in ewes. This combination has favourable effects on milk FA profile from the human consumer's point of view by increasing the nutritional quality of milk.
Keywords: α-linolenic acid, atherogenicitiy index, Cannabis sativa, c9,t11 CLA, n-6/n-3 FA
Introduction
Raising dairy ewes on pasture is justified by the low cost of feeding, and mainly by the nutritional and sanogenetic quality of the milk, compared with that obtained from housed ewes that were given preserved feed (Mierlita, 2012). Pasture-fed ewes can have good milk production if quality grass is abundantly available. Their milk would have a high content in substances beneficial to human health, such as n-3 fatty acids (FAs) and conjugated linoleic acid (CLA). The n-3 FAs, especially eicosapentaenoic acid (EPA, C20:5 n-3) and docosahexaenoic acid (DHA, C22:6 n-3), can reduce the risk of cardiovascular diseases. In experimental animals, c9,t11 CLA (rumenic acid (RA)) has been proved to possess anticancer and anti-atherosclerotic effects, as well as anti-obesity activities (Mills et al., 2011).
In the hot summer months, the nutritional status of sheep, especially lactating ewes, is affected because pastures are available locally mostly from late winter until late spring (Oregui & Falagan-Prieto, 2006). Traditionally, ewes on farms are fed indoor or often on part-time grazing during much of the lactation period. During this period, the decrease of the c9,t11 CLA and n-3 FA contents in milk can be observed (Gomez-Cortes et al., 2009a). Therefore, further research is needed to ascertain the effect of system of feeding (indoor vs. part-time grazing) and oilseed supplementation diet (such as hemp seed) on the CLA isomers and n-3 FA compositions of milk. It has been suggested that hemp seed may be used as a strategy to avoid the decrease in c9,t11 CLA and n-3 FA concentrations in milk. Hemp seed is obtained from industrial hemp (non-drug varieties of Cannabis sativa L., commonly referred to as hemp), and is a traditional plant in Romanian agriculture. The best fibres from the stalk are used in the production for instance of tea bags, paper money and cigarette papers. The ripened seed of hemp is considered a by-product of oil extracted for the cosmetic industry but may be an excellent source of oil and protein in animal nutrition.
Hemp seed was chosen for this study because it is an exceptionally rich source of the two essential FAs, linoleic acid (C18:2 n-6) and α-linolenic acid (C18:3 n-3), and because it has proved efficient in increasing the concentrations of c9,t11 CLA, vaccenic acid (VA) and n-3 FA in the milk fat of cows (Flowers et al., 2008), sheep (Mierlita et al., 2011; Mierlita & Vicas, 2015) and goats (Choi et al., 2013). A further reason is that data about the effects of feeding hemp seed to animals are limited. Hemp seed and hemp seed oil have been used successfully in the diets of laying hens to produce omega-3 enriched eggs (Silversides & Lefrancois, 2005; Gakhar et al., 2012). In no previous study has hemp seed been used as a lipid supplement in ruminant diets.
Therefore, the objective of the present work was to evaluate the effects of a part-time grazing system or indoor feeding and the supplementation of hemp seed in the diet on milk yield, FA profile and health lipid indices in the raw milk of dairy ewes.
Materials and Methods
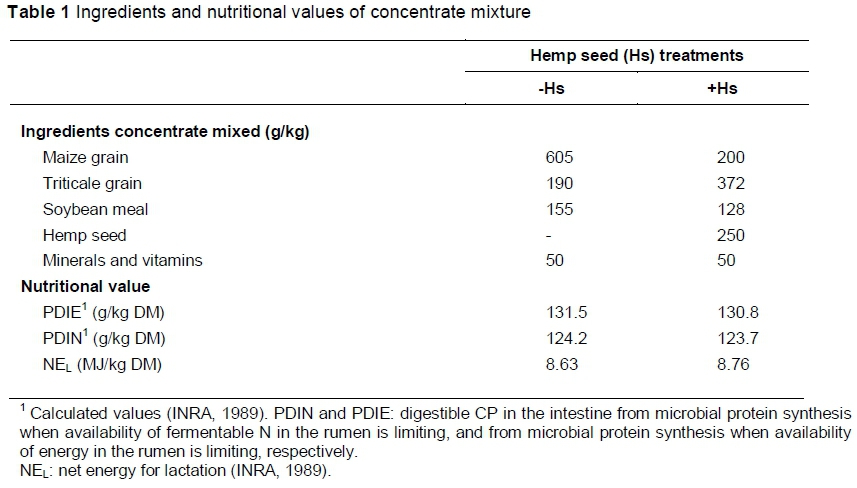
The experiment was conducted at the University of Oradea (Romania) during 10 weeks, from mid June to August 2013. The first three weeks were used as a covariate period (week 1) and adaptation to dietary treatments (weeks 2 and 3). Forty multiparous Turcana ewes (bodyweight (BW) 45.7 ± 1.71 kg) in mid lactation (60 - 80 days of lactation and three months postpartum) were divided into four homogeneous groups (10 ewes/group), balanced for milk yield, BW, days postpartum, number of lactation and number of lambs born. The four groups were assigned randomly to one of four dietary groups arranged in a 2 x 2 factorial experimental design. One of the main treatment factors was the feeding system: indoors during the entire experiment and receiving 1400 g DM grass hay/d; or part-time grazing of 4 h/day + 700 g DM grass hay/d. All animals received 700 g DM concentrate for lactating sheep per day, divided into equal halves and given at the morning and evening milkings. The second main treatment was the supplementation of fat vs. no fat. As a source of fat, hemp seed, was used, which contained 32.7% crude fat and had a high content of linoleic acid (56.5% of total fatty acid methyl esters (FAME)) and linolenic acid (21.2% FAME).
The diets of the four ewe groups were as follows: I/-Hs: indoor feeding with no supplements; I/+Hs: indoor feeding complemented with hemp seed; PG/-Hs: part-time grazing (4 h/d) with no supplements; PG/+Hs: part-time grazing (4 h/d) complemented with hemp seed. Hemp seed was included in the concentrate mixture in a 25% proportion (Table 1), that is, 175 g/d, which provided a fat supplement of approximately 52 g/d for the ewes in the various groups. Ewes in the PG groups grazed on pasture, consisting of a mixture of Festuca rubra, Phleum pratense, Poa pratensis and Trifolium repens. The ewes had ad libitum access to water and vitamin-mineral blocks.
The ewes were milked twice a day (at 07:30 and 20:30), and milk yield was recorded daily. Milk fat, lactose and protein concentrations were recorded on two consecutive days each week. Samples from two consecutive milking sessions were taken in week 6 and week 10 to determine the FA profile of the milk fat.
Samples of diets were collected in weeks 3, 5, 7 and 10 of the experiment (n = 4), stored at -20 °C, and then used for chemical analysis. Samples were analysed for DM (ISO, 1999b), neutral detergent fibre (NDF) and acid detergent fibre (ADF) (Van Soest et al., 1991; on a Fibersac analyzer (Ankom Technology, Fairport, New York, USA), nitrogen (N) (Kjeldahl technique) and ether extract (AOAC, 1996). Starch of the oven-dried materials was determined by a colorimetric method (Dubois et al., 1956). Samples (n = 2) of diets were collected in weeks 6 and 10 to determine the FA profile. These samples were stored immedately at -20 °C, later lyophilized, and kept (about 6 weeks) until analysis.
Milk samples were preserved with two tablets of antimicrobial preservative (Broad Spectrum Microtabs II, Advanced Instruments, Norwood, Massachusetts, USA). The samples were refrigerated at 4 °C before being analysed for fat and protein content by infrared analysis (Milk Analyser System 4000, Foss Electric, Hillerod, Denmark). Monohydrate lactose concentration was measured on these samples using an enzymatic method (FIL, 1991). Samples of milk collected on weeks 6 and 10 for FA analysis were frozen at -20 °C without preservatives.
To determine FA in the diets, FAME were prepared by the one-step extraction-methylation method of Sukhija & Palmquist (1988). To determine the composition of FAs in milk, the fat was extracted according to the international standard, ISO 14156/IDF 172:2001. FAME were prepared according to the method proposed by Christie (1982) and Chouinard et al. (1999); FAME were determined by gas chromatography using a Varian GC 3600 equipped with FID and a fused silica capillary column (SP 2560 Supelco), 100 m x 0.25 mm id, film thickness 0.20 μm. Helium was used as the carrier gas at a flow rate of 1 mL/min. The split ratio was 1 : 100. The oven temperature was programmed at 90 °C and held for 1.50 min, then increased to 210 °C at a rate of 9 °C/min, held at this temperature for 25 min, then increased to 230 °C at 15 °C/min, and held for 7 min. The temperatures of the injector and the detector were set at 270 °C. The identification of FAs was based on external standards, and calculation of the distribution (in weight percentage) was based on the area of each fatty acid ester corrected for the response factors for the individual FAs. Internal standards were used to determine percentage of recovery. The CLA isomer reported is c9,t11 CLA and t10,c12 CLA. The percentage of each fatty acid was calculated by dividing the area under the FA peak by the sum of the areas under total reported FA peaks.
Data obtained were analysed as a 2 χ 2 factorial randomized block design using the PROC MIXED models of SAS (2001). Fixed effects were the feeding system, hemp seeds supplementation and interactions between fixed effects, the time and an appropriate covariate. Random effects of ewes were used as the error term. Variance-covariance structure was first autoregressive [AR(1)] (autoregressive model of order 1). For FA proportion in milk fat, analysis was performed without covariate. Overall differences between treatment means and interaction for feeding system and hemp seeds were considered to be significant for P <0.05.
Results and Discussion
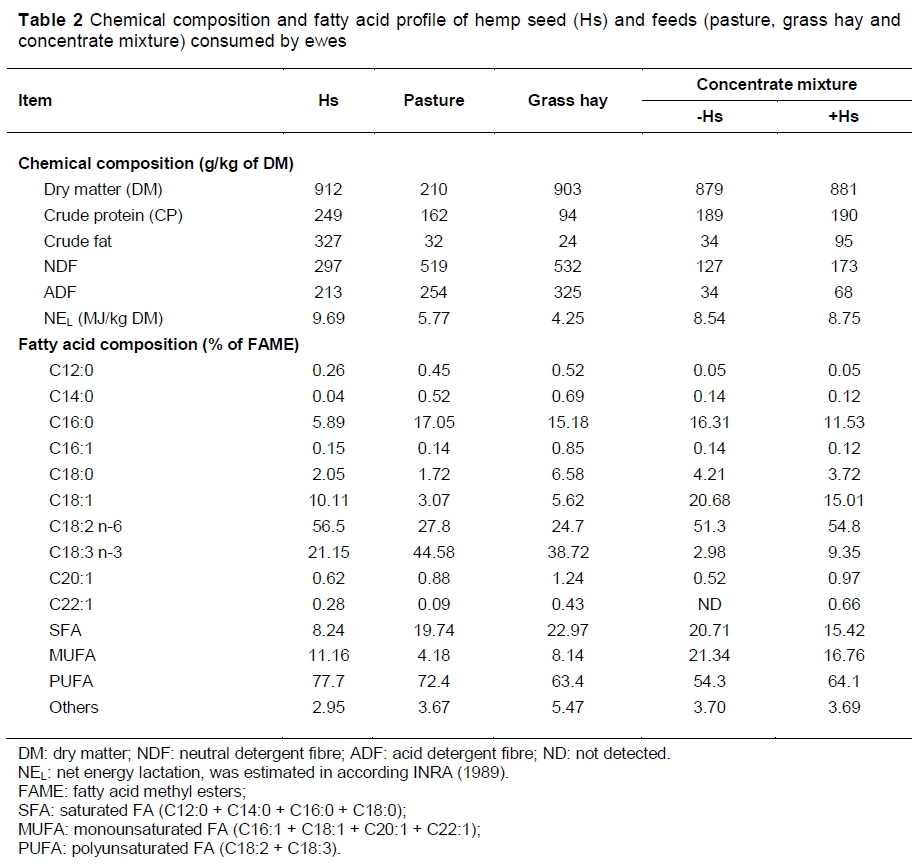
Chemical composition and FA concentrations of feeds (hemp seed, pasture, grass hay and concentrate mixture) are presented in Table 2. As expected, hemp seeds had the highest CP level (249 vs. 94 and 162 g/kg DM in the roughages), while grass hay and pasture had the highest content of cell wall components (NDF and ADF). All feeds, and the concentrate mixture in particular, were good sources of LA, but pasture and hemp seeds were richer in ALA (44.9% and 21.2% of the total FAME, respectively).
The oil content of the hemp seed was 32.7% and the CP was 249 g/kg. Out of the 10 detected FAs, the omega-6 (C18:2 n-6, LA: linoleic acid) was predominant (56.5%), while the omega-3 (C18:3 n-3, ALA: α-linolenic acid) was 21.15% and the omega-9 (C18:1 n-9, oleic acid) was 10.11%. One potential opportunity for hemp seed relates to the relatively high level of ALA (17% - 19%) (Callaway, 2004) as compared with other vegetable oils (<9%) with notable exceptions, such as that of flax. Dietary ALA undergoes a series of elongation and desaturation reactions, leading to the formation of the longer chain n-3 PUFA (EPA, DPA and DHA) that have beneficial effects for human health (Flowers et al., 2008).
Additional hemp seed increased the content of total PUFA, in particular for C18:3 n-3, by more than three times compared with standard concentrate (9.35% vs. 2.98% of FAME). Both C18:2 n-6 and C18:3 n-3 FAs serve as precursors of CLA produced by ruminal biohydrogenation (AbuGhazaleh & Holmes, 2007).
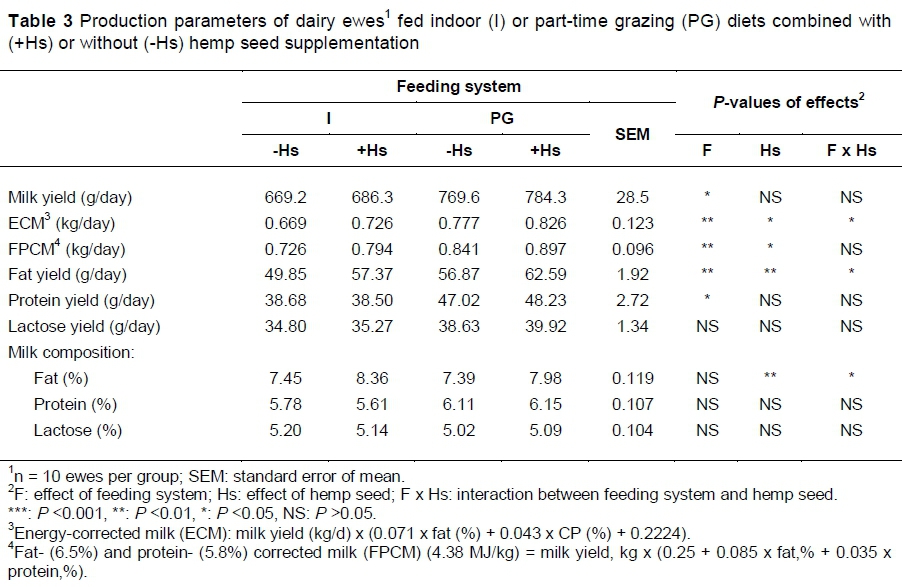
Feeding the PG diets increased raw milk yield and yields of fat, protein and lactose (Table 3). Feeding +Hs increased fat-corrected milk, milk fat content and fat yield and the combination of +Hs and PG increased fat-corrected milk and fat yields. Milk protein content was not affected by dietary factors (Table 3).
The effect of oilseed on sheep milk fat content is controversial. In some studies linseed increased the milk fat concentration in mid-lactation sheep (Caroprese et al., 2011), whereas in others linseed supplementation to early lactation ewes caused milk fat depression (Gómez-Cortés et al., 2014). The rate of PUFA release in rumen is one of the major factors that affect milk fat content. As a result of that, fats with a high ruminal degradability adversely affect milk fat content, unlike some oleaginous seeds (e.g. flaxseeds) that have low ruminal degradability (Mustafa et al., 2003b). Therefore, it can be assumed that the slow release of unsaturated FA from the hemp seed in rumen decreased the amount of trans FAs and in this way milk fat depression was avoided (Zhang et al., 2006).
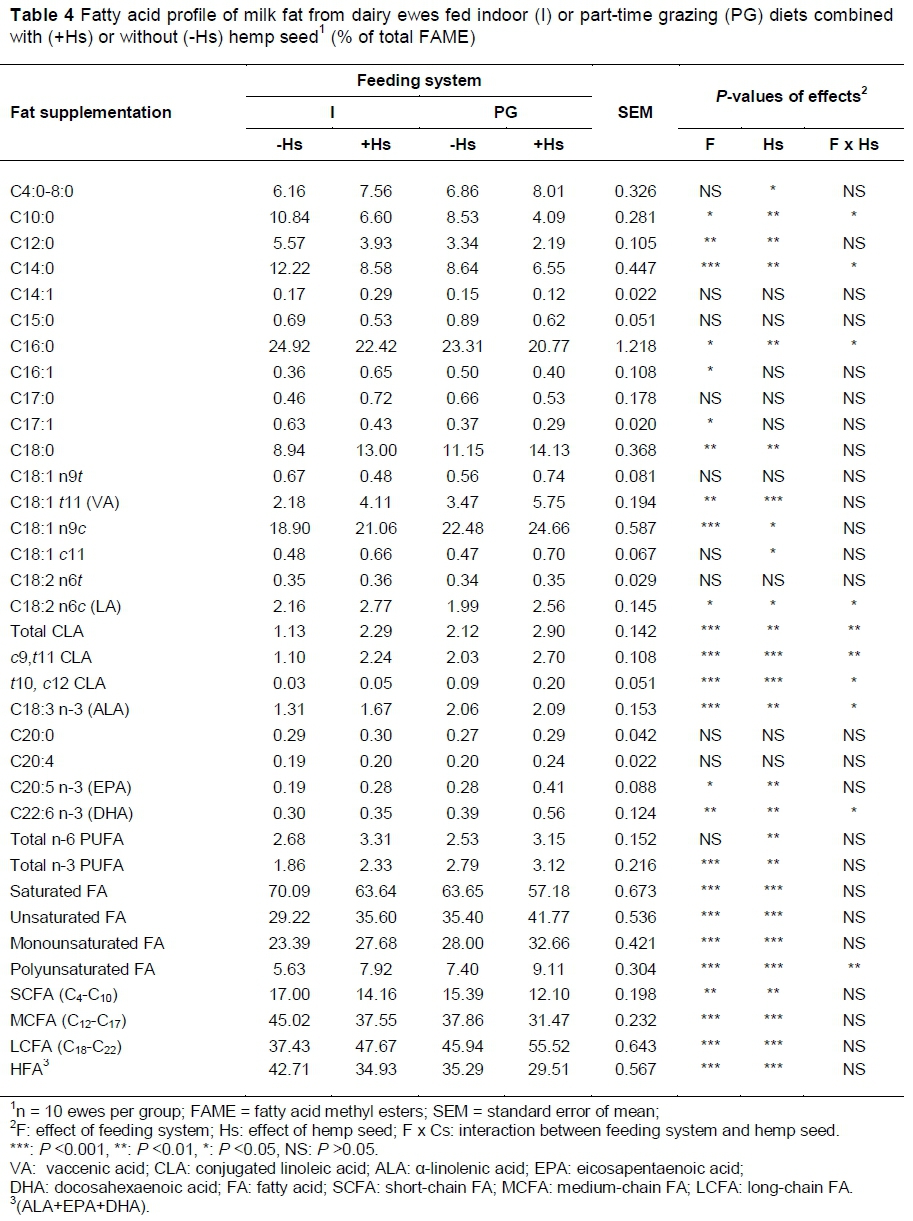
The effect of the treatments on the FA profile of milk fat is presented in Table 4. In dairy ewes fed indoors (I vs. PG), the concentration of saturated FAs in milk fat (C10:0, C12:0, C14:0, C16:0) increased, and the monounsaturated (C16:1, C18:1 t11 and C18:1 c9) and polyunsaturated FA (c9,t11 CLA; t10,c12 CLA; ALA; EPA and DHA) concentrations decreased. For PG ewes the high milk fat content of ALA, VA and CLA are related to the high content of ALA in green pasture (44.58% of FAME). Milk fat from PG diet ewes (supplemented or not with hemp seed) had over 80% more c9,t11 CLA isomer than milk fat from the indoor diet ewes. As pasture intake increased, the concentration of ALA (R2 = 0.69) and CLA (R2 = 0.79) in milk increased (Renobales et al., 2012). Dhiman et al. (2000) reported that cows grazing pasture had three times higher CLA concentrations in milk fat (2.21% of total FA) compared to cows fed a diet containing 50% conserved forage (hay and silages) and 50% grain (0.38% of total FA). In dairy sheep fed on pasture, the concentrations of c9,t11 CLA, VA and ALA in milk fat were the highest in late winter, early spring, when grass availability was the highest, and decreased as lactation progressed and pasture availability and quality decreased (Nudda et al., 2014). c9,t11 CLA is synthesised in the rumen by anaerobic bacteria (B. fibrisolvens) from LA. Furthermore, c9,t11 CLA can be produced in an alternative way in the mammary gland via Δ9-desaturase activity from VA. The importance of VA lies, therefore, in its role as a precursor in the synthesis of c9,t11 CLA in the mammary gland and also in some human tissues (Bauman & Griinari, 2003). The endogenous synthesis of c9,t11 CLA from VA has been proposed as the major pathway of c9,t11 CLA synthesis in milk, accounting for an estimated 80% of the c9,t11 CLA in milk fat (Mosley et al., 2006). The t10,c12 CLA produced in the rumen may be the result of minor bacterial populations or the alternative, biohydrogenation pathway in the rumen (Tsiplakou & Zervas, 2008). The water content of the grass affects microbiological fermentation and pH in the rumen. Rumen pH has an important role in maintaining a viable rumen environment suitable for B. fibrisolvens. It has been shown that a ruminal pH of 6.0 or above has a positive effect on CLA isomer production (Tsiplakou & Zervas, 2008). It is known that cows grazing on pasture have a higher ruminal pH that is favourable for cellulolytic bacteria growth in the rumen, responsible for CLA and VA production (Jenkins et al., 2008).
Part-time grazing significantly increased the concentration of PUFAs, such as ALA, EPA and DHA in milk. Grass contains a low level of fat, but α-linolenic acid accounts for 44.5% of total FAs. Cabiddu et al., 2005 showed that in grass 50% - 75% of the total fat is n-3 FA. ALA is the precursor of such n-3 PUFAs as EPA and DHA, which efficiently prevent coronary heart disease.
Increases in the concentrations of C18:0, C18:1 c9 and VA (P <0.01) in milk fat and a decrease in the concentration of C16:0 were recorded in part-time grazing ewes as compared with those fed indoors (P <0.05). The increase of C18:0 concentration in PG diet ewes' milk fat must have been due to a more complete hydrogenation of PUFA into C18:0 which probably helped the increase in C18:1 c9 in milk fat, as this FA is endogenously synthesized in the mammary gland via Δ9- desaturase of C18:0 (Chilliard et al., 2003). Similar changes of milk FAs profile were found at grazing cows compared to those fed with a standard total mixed ration (Morales-Almaraz, et al., 2010). Couvreur et al. (2006) reported a linear relationship between the proportion of fresh grass in dairy cow diets and milk VA. AbuGhazaleh & Jenkins (2004a) showed that DHA promotes VA accumulations in rumen by inhibiting the reduction of VA to C18:0.
If we look at the impact of dietary fat on human health, the increase in ALA, CLA and C18:1 c9 in milk fat represents an advantage of part-time grazing diets. The effort to increase the c9,t11 CLA and PUFA n-3 concentrations and change the fatty acid profile in milk by dietary manipulation may provide a functional food for human consumption.
The value of hemp seed from the point of view of the primary components is its FA composition, because it contains the two dietary essential FAs, namely LA (50% - 60%) and ALA (20% - 25%) (Callaway, 2004). Dietary hemp seed modified milk FA composition towards a lower level of SFA and higher levels of PUFA and MUFA, confirming how an adequate dietary strategy can improve sheep milk quality. Including hemp seed in the diet resulted in a significant increase in both total CLA (c9,t11 CLA and t10,c12 CLA) and VA, as well as ALA in milk fat (P <0.01). Interactions among feeding system and hemp seed resulted in a significant increase in total CLA, ALA and C18:1 c9 concentrations.
The increases in milk n-3 FA concentrations with the +Hs treatments were relatively small compared with their intake (Table 2). Similar low increases in n-3 FA were reported by others (Flowers et al., 2008; Mierlita et al., 2011) when n-3 lipid supplements were used. This low transfer of n-3 FA from feed to milk fat might be explained by extensive ruminal biohydrogenation of linolenic acid and DHA (AbuGhazaleh & Jenkins, 2004a) or by their partitioning toward other tissues within the body. n-3 FAs are almost totally confined to plasma cholesterol ester and phospholipids, which are poorly taken up by the mammary gland (Kitessa et al., 2003).
Increase in the C18 FA mainly at the expense of C10 to C16, the response to feeding supplements with hemp seed rich in PUFA, is determined probably by long-chain FAs that are powerful inhibitors of de novo lipogenesis in the mammary gland, and this effect is more marked when FAs have a long chain, are more unsaturated and contain more trans double bonds (Chilliard et al., 2003).
The levels of c9,t11 CLA and t10,c12 CLA in milk fat were higher when the ewes were fed hemp seed (P <0.001). Ewes fed +Hs combined with grazing diet had the lowest proportion of C10:0, C14:0 and C16:0 (feeding system x hemp seed: P <0.05) in milk fat. Conversely, they had the highest proportion of LA, c9,t11 CLA, t10,c12 CLA, ALA and DHA in milk fat (feeding system x hemp seed, P <0.05 to <0.01).
Taking into account that ewe milk is generally used for cheese making and that milk transformation does not influence the CLA and n-3 PUFA content of dairy products (Nudda et al., 2014) grazing management and feeding hemp seed could become a useful strategy to naturally manipulate dietetic characteristics of milk and dairy products.
Feeding system part-time grazing of ewes decreased the milk concentrations of short-chain FA and medium-chain FA and increased of long-chain FA (LCFA) (P <0.001). Many authors have noted a reduction in milk short- and medium-chain FAs with an increased fresh pasture supply in the diet (Bargo et al., 2006; Nantapo et al., 2014). Grazed grass generally increases the levels of milk C18:1 c9, PUFA, especially ALA and c9,t11 CLA, and decreases that of saturated medium-chain FA (Chilliard et al., 2003). Hemp seed supplementation decreased milk short-chain FA (SCFA) and medium-chain FA (MCFA) (P <0.01 to <0.001). This decrease could be due to a negative effect of intermediates of rumen biohydrogenation of dietary PUFA on activity of acetyl-CoA carboxylase or fatty acid synthetase (Chilliard et al., 2003).
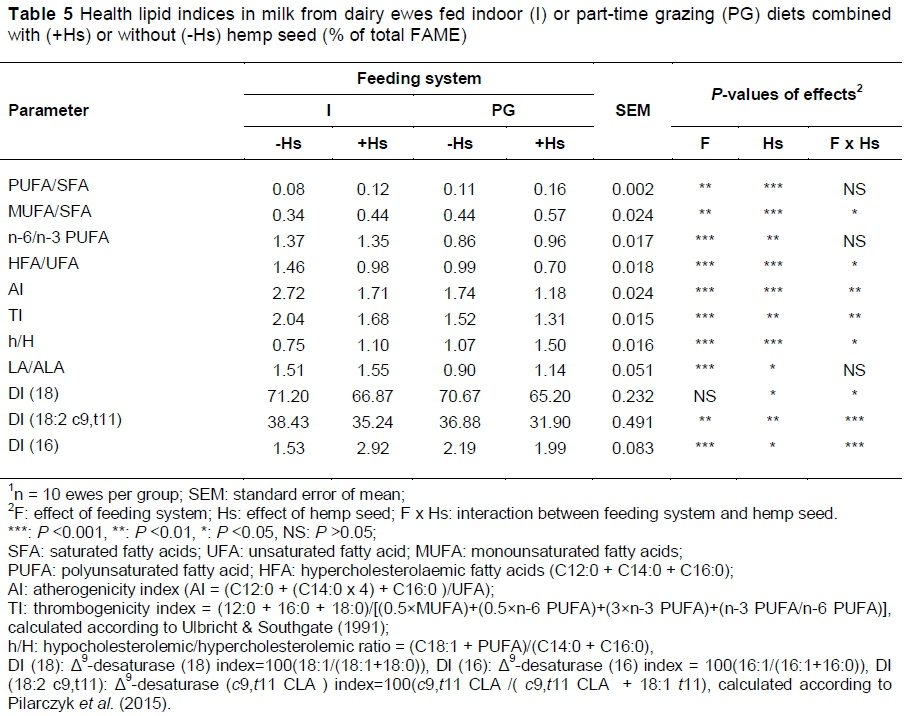
The PUFA/SFA and n-6/n-3 PUFA ratios, atherogenity index (AI) and thrombogenity index (TI) are commonly used to assess the nutritional value and consumer health of animal fat (Pilarczyk et al., 2015). In general, a ratio of PUFA to SFA above 0.45 and a ratio of n-6/n-3 below 4.0 are required in the diet to combat 'lifestyle diseases' such as coronary heart disease and cancers (Simopoulos, 2002). In the present study, the PUFA/SFA ratios (0.08 - 0.16) were considerably lower than the recommended values, whereas the n-6/n-3 ratios (0.86 - 1.37) were within the recommended levels (Table 5). The PUFA/SFA and MUFA/SFA ratios in the milk from ewes feeding part-time grazing were significantly (P <0.01) higher compared with the milk of ewes fed indoor. A low PUFA/SFA ratio in ewes fed indoor resulted from a high level of some SFAs in the milk, especially that of C10:0, C16:0 and C14:0 acids, whose consequence was a very high total content of SFA. The introduction of hemp seeds in ewes' feed of +Hs' groups provided the most beneficial ratios of PUFA/SFA (0.12-0.16) and MUFA/SFA (0.44 - 0.57) in milk fat. The high levels of C16:0 and C14:0 in the milk of ewes +Hs groups is partly countered by the contents of MUFA, in particular C18:1 c9, and PUFA. In the current study the ratio PUFA/SFA in milk were higher that those reported in dairy cows fed with a total mixed ration or under a pasture-based dairy system (Nantapo et al., 2014; Pilarczyk et al., 2015).
In the present study, the milk from PG ewes (part-time grazing groups) had a significantly (P <0.001) higher percentage of ALA and CLA in the total content of FAs, whereas the milk from I ewes (indoor groups) had a significantly (P <0.05) higher percentage of LA. The milk from +Hs ewes (I/+Hs and PG/+Hs groups) had a significantly (P <0.05) higher percentage of LA, ALA and CLA in the total content of FAs. The significantly higher production of ALA and CLA for part-time grazing ewes compared with ewes fed indoors was reported by Renobales et al. (2012), while Zhang et al. (2006) observed the milk from ewes feeding oilseeds rich in linoleic and linolenic FAs had a significantly higher percentage of LA, ALA and CLA. The LA/ALA ratio in the milk from PG ewes was significantly (P <0.001) lower compared with the milk of I ewes, which indicates a more favourable proportion of these acids in the milk from ewes on part-time grazing. Similar findings were recorded in the group supplemented with hemp seeds (+Hs groups), allowing the conclusion that feeding hemp seed and permitting part-time grazing improve the FA profile and health lipid indices in raw milk.
In the current study the ratio CLA/VA (DI 18:2 c9,t11) in milk was much lower in the part-time grazing and +Hs groups, suggesting the important contribution of VA to CLA synthesis in the mammary gland by the epithelial tissue Δ9-desaturase (Griinari et al., 2000).
The atherogenic index (AI) and thrombogenic index (TI) take into account the effects that single FAs might have on human health and, in practice, on the probability of increasing the incidence of pathogenic phenomena such as atheroma and/or thrombus formation (Pilarczyk et al., 2015). In the present study, the AI and TI in the milk from part-time grazing feeding ewes (PG groups) was significantly (P <0.001) lower compared with the milk of ewes fed indoors. Much lower values of AI and TI were obtained by dietary supplementation with hemp seed (+Hs groups).
An increase of c9,t11 CLA, ALA, EPA and DHA in milk could be achieved by part-time grazing and feeding hemp seed to ewes. Grass and hemp seed feeding reduces the n-6/n-3 ratio, atherogenicity index (AI) and thrombogenity index (TI) in milk, which improves human health because of the beneficial effect on the cardiovascular system.
Conclusions
Following a part-time grazing system in lactating ewes was a useful way of improving the FA profile of milk since it had a significantly higher concentration of unsaturated FAs, such as VA, CLA isomers, n-3 FA (ALA, EPA, DHA) and total PUFAs. Based on results from the current study, diet supplementation of hemp seed, to lactating ewes could be an effective way to produce a potentially healthier FA profile high in CLA isomers and n-3 FA in ewes' milk without detrimental effects on the milk production. This feeding strategy is particularly useful when the diet of ewes has a poor content and composition of FAs, that is, when stored forages are used. The PUFA/SFA, n-6/n-3 PUFA, LA/ALA ratios and AI and TI in the milk from feeding hemp seed and part-time grazing ewes were also more favourable.
Acknowledgements
This work was supported by CNCSIS-UEFISCDI, project number PN II - IDEI 679/2008.
Conflict of Interest Declaration
The author declare that there was no conflict of interest.
References
AbuGhazaleh, A.A. & Jenkins, T.C., 2004. Disappearance of docosahexaenoic and eicosapentaenoic acids from cultures of mixed ruminal microorganisms. J. Dairy Sci. 87, 645-651. [ Links ]
AbuGhazaleh, A.A. & Holmes, L.D., 2007. Diet supplementation with fish oil and sunflower oil to increase conjugated linoleic acid levels in milk fat of partially grazing dairy cows. J. Dairy Sci. 90, 2897-2904. [ Links ]
AOAC, 1996. Official Methods of Analysis. Vol. I. 16th ed. Association of Official Analytical Chemists, Arlington, VA., USA. [ Links ]
Bargo, F., Delahoy, J.E., Schroeder, G.F., Baumgard, L.H. & Muller, L.D., 2006. Supplementing total mixed rations with pasture increase the content of conjugated linoleic acid in milk. Anim. Feed Sci. Technol. 131, 226-240. [ Links ]
Bauman, D.E. & Griinari, J.M., 2003. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 23, 203-227. [ Links ]
Cabiddu, A., Decandia, M., Addis, M., Piredda, G., Pirisi, A. & Molle, G., 2005. Managing Mediterranean pastures in order to enhance the level of beneficial fatty acids in sheep milk. Small Rumin. Res. 59, 169-180. [ Links ]
Callaway, J.C., 2004. Hempseed as a nutritional resource: An overview. Euphytica 140, 65-72. [ Links ]
Caroprese, M., Albenzio, M., Bruno, A., Fedele, V., Santillo, A. & Sevi, A., 2011. Effect of solar radiation and flaxseed supplementation on milk production and fatty acid profile of lactating ewes under high ambient temperature. J. Dairy Sci. 94, 3856-3867. [ Links ]
Chilliard, Y., Ferlay, A., Rouel, J. & Lambere, G., 2003. A review of nutritional and physiological factors affecting goat milk synthesis and lipolysis. J. Dairy Sci. 86, 1751-1770. [ Links ]
Choi, S., Kim, J.K., Lee, H.G., Choi, C.W., Choi, Y.I. & Song, M.K., 2013. Cis-9, trans-11-conjugated linoleic acid in dairy goat milk was increased by high linoleic (soybean oil) or linolenic (linseed oil) acid diet. Korean J. Food Sci. An. 33, 487-492. [ Links ]
Chouinard, P.Y., Corneau, L., Barbano, D.M., Metzger, L.E. & Bauman, D.E., 1999. Conjugated linoleic acids alter milk fatty acid composition and inhibit milk fat secretion in dairy cows. J. Nutr. 129, 1579-1584. [ Links ]
Christie, W.W., 1982. A simple procedure of rapid transmethylation of glycerolipids and cholesterol esters. J. Lipid Res. 23, 1072-1075. [ Links ]
Couvreur, S., Hurtaud, C., Lopez, C., Delaby, L. & Peyraud, J.L., 2006. The linear relationship between the proportion of fresh grass in the cow diet, milk fatty acid composition, and butter properties. J. Dairy Sci. 89, 1956-1969. [ Links ]
Cozma A., 2013. The profile of certain liposoluble components in different types of milk. Summary of the PhD thesis. www.umfcluj.ro/ro/sustinute./9-rezumate-2013. [ Links ]
Dhiman, T.R., Satter, L.D., Pariza, M.W., Galli, M.P., Albright, K. & Tolosa, M.X., 2000. Conjugated linoleic acid (CLA) content of milk from cows offered diets rich in linoleic and linolenic acid. J. Dairy Sci. 83, 1016-1027. [ Links ]
Dubois, M., Gilles, K., Hamilton, K., Rebers, A. & Smith, F., 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350-356. [ Links ]
FIL (Federation Internationale Laitie're) 1991. Laits secs, mélanges secs pour cremes glacées et fromages fondus. Determination de la teneur en lactose (méthodes enzymatiques), 79B. IDF-FIL, Brussels, Belgium. (in France). [ Links ]
Flowers, G., Ibrahim, S.A. & AbuGhazaleh, A.A., 2008. Milk fatty acid composition of grazing dairy cows when supplemented with linseed oil. J. Dairy Sci. 91, 722-730. [ Links ]
Gakhar, N., Goldberg, E., Jing, M., Gibson, R. & House, J.D., 2012. Effect of feeding hemp seed and hemp seed oil on laying hen performance and egg yolk fatty acid content: Evidence of their safety and efficacy for laying hen diets. Poult. Sci. 91, 701-711. [ Links ]
Gómez-Cortés, P., Frutos, P., Mantecón, A.R., Juárez, M., de La Fuente, M.A. & Hervás, G., 2009. Effect of supplementation on grazing dairy ewes with a cereal concentrate on animal performance and milk fatty acid profile. J. Dairy Sci. 92, 3964-3972. [ Links ]
Gómez-Cortés, P., Gallardo, B., Mantecón, A.R., Juárez, M., de la Fuente, M.A. & Manso T., 2014. Effects of different sources of fat (calcium soap of palm oil vs. extruded linseed) in lactating ewes' diet on the fatty acid profile of their suckling lambs. Meat Sci. 96, 1304-1312. [ Links ]
Griinari, J.M., Corl, B.A., Lacy, S.H., Chouinard, P.Y., Nurmela, K.V. & Bauman, D.E., 2000. Conjugated linoleic acid is synthesized endogenously in lactating dairy cows by Delta(9)-desaturase. J. Nutr. 130, 2285-2291. [ Links ]
INRA, 1989. Ruminant Nutrition. Recommended Allowances and Feed Tables. Ed: Jarrige, R., John Libbrey & INRA, London & Paris. pp. 193-212. [ Links ]
ISO, 1999. Animal Feeding Stuffs. Determination of Moisture and Other Volatile Matter Content. International Organization for Standardization, Geneva, Switzerland. [ Links ]
Jenkins, T.C., Wallace, R.J., Moate, P.J. & Mosley, E.E., 2008. Recent advances in biohydrogenation of unsaturated fatty acids within the rumen microbial ecosystem. J. Anim. Sci. 86, 397-412. [ Links ]
Kitessa, S.M., Peake, D., Bencini, R. & Williams, A.J., 2003. Fish oil metabolism in ruminants: III. Transfer of n-3 polyunsaturated fatty acids (PUFA) from tuna oil into sheep's milk. Anim. Feed Sci. Technol. 108, 1-14. [ Links ]
Mierlita, D., 2012. Effect of feeding type (pasture vs. total mixed rations) of Turcana ewes on animal performance and milk fatty acid profile. J. Food Agric. Environ. 10, 815-818. [ Links ]
Mierlita, D. & Vicas, S., 2015. Dietary effect of silage type and combination with camelina seed on milk fatty acid profile and antioxidant capacity of sheep milk. S. Afr. J. Anim. Sci. 45, 1-11. [ Links ]
Mierlita, D., Daraban, St., Lup, F. & Chereji, A., 2011. The effect of grazing management and camelina seed supplementation in the diet on milk performance and milk fatty acid composition of dairy ewes. J. Food Agric. Environ. 9, 368-373. [ Links ]
Mills, S., Ross, R.P., Hill, C., Fitzgerald, G.F. & Stanton, C., 2011. Milk intelligence: mining milk for bioactive substances associated with human health. Int. Dairy J. 21, 377-401. [ Links ]
Morales-Almaraz, E., Soldado, A., Gonzalez, A., Martinez-Fernandez, A., Dominiquez-Vara, I., de la Roza-Delgado, B. & Vicente, F., 2010. Improving the fatty acid profile of dairy cow milk by combining grazing with feeding of total mixed ration. J. Dairy Res. 77, 225-230. [ Links ]
Mosley, E.E., Shafii, B., Moate, P.J. & McGuire, M.A., 2006. Cis-9, trans-11 conjugated linoleic acid is synthesized directly from vaccenic acid in lactating dairy cattle. J. Nutr. 136, 570-575. [ Links ]
Mustafa, A.F., McKinnon, J.J. & Christensen, D.A., 1999. The nutritive value of hemp meal for ruminants. Can. J. Anim. Sci. 79, 91-95. [ Links ]
Mustafa, A.F., Chouinard, P.Y., Ouellet, D.R. & Soita, H., 2003. Effects of moist heat treatment on ruminal nutrient degradation of sunflower seed. J. Sci. Food Agric. 83, 1059-1064. [ Links ]
Nantapo, C.T.W., Muchenje, V. & Hugo, A., 2014. Atherogenicity index and health-related fatty acids in different stages of lactation from Friesian, Jersey and Friesian x Jersey cross cow milk under a pasture-based dairy system. Food Chem. 146, 127-133. [ Links ]
Nudda, A., Battacone, G., Neto, O.B., Cannas, A., Francesconi, A., Atzori, A.S. & Pulina G., 2014. Feeding strategies to design the fatty acid profile of sheep milk and cheese. R. Bras. Zootec. 43, 445-456. [ Links ]
Oregui, L.M. & Falagan-Prieto, A., 2006. Spécificités et diversité des systèmes de production o, vine et caprine dans le Bassin Méditerranéen (Specificity and diversity of sheep and goat production systems in Mediterranean Basin). Options Méditerranéenes A, 70, 77-86. [ Links ]
Pilarczyk, R., Wójcik, J., Sablik, P. & Czerniak, P., 2015. Fatty acid profile and health lipid indices in the raw milk of Simmental and Holstein-Friesian cows from an organic farm. S. Afr. J. Anim. Sci. 45, 30-38. [ Links ]
Renobales, M., Amores, G., Arranz, J., Virto, M., Barrón, L.J.R., Bustamante, M.A., Ruiz de Gordoa, J.C., Nájera, A.I., Valdivielso, I., Abilleira, E., Beltrán de Heredia, I., Pérez-Elortondo, F.J., Ruiz, R., Albisu, M. & Mandaluniz, N., 2012. Part-time grazing improves sheep milk production and its nutritional characteristics. Food Chem. 130, 90-96. [ Links ]
SAS, 2001. SAS System for Microsoft Windows. Release 8.2. SAS Institute Inc., Cary, NC., USA. [ Links ]
Silversides, F.G. & Lefrancois, M.R., 2005. The effect of feeding hemp seed meal to laying hens. Br. Poult. Sci. 46, 231-235. [ Links ]
Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharmacother. 56, 365-379. [ Links ]
Sukhija, P.S. & Palmquist, D.L., 1988. Rapid method for determination of total fatty acid content and composition of feedstuffs and faeces. J. Agric. Food Chem. 36, 1202-1206. [ Links ]
Tsiplakou, E. & Zervas, G., 2008. The effect of dietary inclusion of olive tree leaves and grape marc on the content of conjugated linoleic acid and vaccenic acid in the milk of dairy sheep and goats. J. Dairy Res. 75, 270-278. [ Links ]
Ulbricht, T.L.V. & Southgate, D.A.T., 1991. Coronary heart disease: seven dietary factors. Lancet 338, 985-992. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Zhang, R.H., Mustafa, A.F. & Zhao, X., 2006. Effects of feeding oilseeds rich in linoleic and linolenic fatty acids to lactating ewes on cheese yield and on fatty acid composition of milk and cheese. Anim. Feed Sci. Technol. 127, 220-233. [ Links ]
Received 7 April 2015
Accepted 8 June 2016
First published online 2 August 2016
# Corresponding author: dadi.mierlita@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}