










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.46 n.1 Pretoria 2016
http://dx.doi.org/10.4314/sajas.v46i1.2
Effects of glucose oxidase on the growth performance, serum parameters and faecal microflora of piglets
H. TangI, III; B. YaoI, #; X. GaoI, #; P. YangI; Z. WangII; G. ZhangI, III
IKey Laboratory for Feed Biotechnology of the Ministry of Agriculture, Feed Research Institute, Chinese Academy of Agricultural Sciences, Beijing 100081, P. R. China
IINational Engineering Research Centre of Biological Feed, Beijing 100081, P. R. China
IIIBeijing Challenge Bio-technology Limited Company, Beijing 100081, P. R. China
ABSTRACT
The experiment was conducted to investigate the effects of diets supplemented with glucose oxidase (GOD) on growth performance, serum parameters and faecal microflora of piglets. One hundred and twelve piglets (35 days old) were randomly assigned to two groups (four replicates per group, half male and half female, and 14 piglets per replicate) and fed a diet with or without 100 U GOD per kg, for 35 days. Feeding GOD caused a higher average daily weight gain and feed intake, and lower feed conversion ratio (FCR) of piglets. No significant difference was observed in the reference values of serum biochemical parameters between the groups fed with or without GOD. The contents of triiodothyronine, thyroxine and growth hormone of piglets fed GOD were higher than those of the control. Moreover, GOD supplementation suppressed the concentration of faecal Salmonella. The results showed that supplementation of GOD to diets promoted growth performance, increased the contents of growth and development-related hormones, and improved the faecal microflora of growing piglets.
Keywords: Glucose oxidase, intestinal health, performance, swine
Introduction
Glucose oxidase (GOD) is a flavoprotein, which catalyses the oxidation of β-D-glucose to D-glucono-δ-lactone and hydrogen peroxide (H2O2), using molecular oxygen as an electron acceptor (Hatzinikolaou et al., 1996; Pluschkell et al., 1996). Glucose oxidase has several commercial applications, for example improving colour, flavour and shelf-life of food materials, removing oxygen from fruit juices and canned beverages, inhibiting the growth of pathogens and preventing rancidity in mayonnaise (Tiina et al., 1989; Malherbe et al., 2003; Seifu et al., 2004; Opwis et al., 2008; Bankar et al., 2009). Glucose oxidase can catalyse the formation of gluconic acid, and has been used as a food additive to regulate the acidity of sterilization solutions and bleaching agents in food manufacturing and as a salt in chemical components for medication (Nakao et al., 1997; Klein et al., 2002). Since 1999, GOD has been supplemented into 12 feed preparations, according to the Catalogue of Feed Additive Varieties released by the Ministry of Agriculture, People's Republic of China. There are broad prospects for GOD application in the feed industry. However, owing to low fermentation capacity and high cost, the application of GOD is limited as a feed additive. Recently, the Laboratory for Food Technology developed a new recombinant GOD with high yield (615 U/mL and 2.5 g protein/L in a 3 L fermenter) and favourable properties (Gao et al., 2012).
Various commensal and pathogenic bacterial species are present in the gastrointestinal tracts of pigs, especially pathogenic, which represent a major source of economic loss to the pig industry worldwide (Pluske et al., 2002; Namroud et al., 2010). Of these, haemolytic enterotoxigenic, Escherichia coli, is the primary infectious agent of post-weaning colibacillosis, which provokes hypersecretory diarrhoea through the release of specific enterotoxins (Argenzio, 1992; Jones et al., 2001). Salmonella is associated with diarrhoeic piglets, with lesions more typical of enterotoxigenic diarrhoeal disease than salmonellosis (Reed et al., 1985). Schwartz (1999) remarked that most diarrhoeic piglets were associated with poor hygiene, concurrent enteric pathogens, inappropriate diets and a poor environment. It is common for haemolytic E. coli and Salmonella to appear in the faeces of pigs in increased numbers in the first week after weaning in both healthy and diarrhoeic pigs (Hampson et al., 1985). Serum biochemical parameters are good indicators of animal health.
Numerous studies have been conducted to determine fundamental values for serum biochemical and haematological parameters of pigs. For example, triiodothyronine (T3) and thyroxine (T4) are tyrosine-based hormones produced by the thyroid gland, which are primarily responsible for regulation of the pig's metabolism (Decuypere et al., 1983; 2005). However, no experiment has been designed to evaluate the interrelationships between blood constituent changes and GOD. Therefore, the present study was designed to evaluate the effects of GOD as a feed additive on growth performance, serum parameters and faecal microflora of piglets.
Materials and Methods
The GOD preparation and millrun were provided by the Challenge Group, Beijing, People's Republic of China. All pigs used in this study were cared for in accordance with the FASS Guide for the Care and Use of Agricultural Animals in Research and Teaching (2012).
The GOD derived from Pénicillium notatum was modified and produced in Pichia pastoris with a high yield (Gao et al., 2012). Recombinant GOD had optimal activity at 35 - 40 °C and pH 6.2, and was stable at pH 3.0 to 7.0 at 50 °C. The optimized recombinant GOD yielded 615 U/mL (2.5 g protein/L) in a 3 L fermenter. One GOD unit is defined as that quantity of enzyme that liberates 1 μmol of H2O2 per minute under the conditions of 37 °C and pH 5.5.
All pigs were fed a commercial starter diet for an adaptation period of one week prior to the start of the experiment. The diets consisted of the control (no enzyme) and a control supplemented with GOD (100 U/kg diet). The control diet was formulated to meet or exceed the requirements of the Feeding Standard of Swine of the People's Republic of China (2004) and was composed of maize, soybean meal, soybean oil, extruded soybean, fermented soybean meal, fish meal, glucose, etc. (Table 1).
One hundred and twelve piglets, weaned at 21 ± 1 d old, were obtained from a local livestock breeding farm in Beijing, China, and housed 14 in a pen with plastic-covered expanded metal floors. The average weight of the piglets was recorded at the end of the adaptation period. The piglets were then randomly assigned in two groups for control and treatment based on sex and bodyweight. Each group contained four pens (half male and half female), and each pen contained 14 pigs. Pigs had unlimited access to feed and water throughout the five-week study. The room temperature was initially maintained at 28 °C on week 1, and gradually decreased at about 1.5 °C per week.
Bodyweight and feed intake were monitored at d 35 to determine average daily gain (ADG), average daily feed intake (ADFI) and feed conversion ratio (FCR). Blood samples were collected from two pigs/pen via jugular venipuncture on d 35. Samples (approximately 6 mL/tube) were collected into plastic tubes without anticoagulant. The blood was allowed to clot at room temperature, followed by centrifugation at 1, 155 χ g for 5 min at room temperature. Serum was collected and stored at -20 °C until further analysis. The contents of albumin, globulin and total protein were determined by automatic biochemical analyser (7600; Hitachi Medical (Guangzhou) Co., Ltd., Guangzhou, China), and the coefficient of serum proteins was calculated. The concentrations of T3, T4, thyroid-stimulating hormone, growth hormone (GH), insulin and glucagon were determined using radioimmunoassay (Hunter, 1986).
Fresh faecal samples were collected immediately after defecation from three pigs per pen on d 14 and d 35, and subjected to microflora analysis right away. The total counts of bacteria and E. coli and Salmonella were determined by the dilution method on LB agar (peptone 10.0 g/L, yeast power 5.0 g/L, NaCl 9.0 g/L, agar 15.0 g/L, pH 7.2 and MacConkey agar plates (peptone 20.0 g/L, lactose 10.0 g/L, bile salt 5.0 g/L, NaCl 5.0 g/L, toluylene red 0.03 g/L, and agar 14.0 g/L, pH 7.1)), respectively. Lactobacilli were enumerated by using MRS agar plates (peptone 10.0 g/L, beef extract 10.0 g/L, yeast powder 5.0 g/L, diammonium citrate 2.0 g/L, sodium acetate 5.0 g/L, glucose 20.0 g/L, twain 80 1.0 mL/L, MgSO4.7H2O 0.58 g/L, K2HPO4 2.0 g/L, MnSO4.7H2O 0.25 g/L, agar 15.0 g/L, pH 6.8). Approximately 1.0 g of fresh faecal sample was suspended in 99.0 mL of sterile 0.9% normal saline solution, vibrated for 10 min (35 χ g, 20 °C), and serially diluted 1.0 mL to 9.0 mL in the same saline solution. Dilutions of 104 to 106 (0.1 mL) were plated on agar plates and incubated at 37 °C for 24 h. The total counts of bacteria E. coli, Salmonella and Lactobacilli were then determined. All plates were done in triplicate, and the results were reported as cfu per gram of fresh faecal sample.
Bacterial enumeration data were transformed into lgcfu/g before statistical analysis. The sex effect was insignificant, and was excluded from the model. A pen of 14 pigs was treated as an experimental unit for growth performance data analysis, and each pig was a unit for the serum parameter analysis. The data were submitted to ANOVA using the GLM procedure of SAS (SAS, 2004). Significant differences (P <0.05) were determined by Duncan's multiple range test.
Results and Discussion
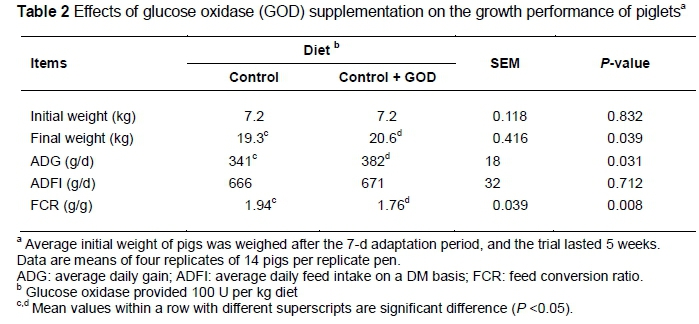
The growth performance of pigs in the various groups is summarized in Table 2. No mortality was observed in this experiment. Piglets on control and treatment groups were similar in ADFI (P >0.05), but showed significant differences in the final weight, ADG and FCR (P <0.05). Pigs fed GOD-supplemented diet had higher ADG (382 g/d vs. 341 g/d) and lower FCR (1.76 vs. 1.94) values than those fed the control diet. The natural function of GOD is closely related to its catalytic activity. It catalyses the oxidation of β-D-glucose to gluconic acid by utilizing molecular oxygen as an electron acceptor with simultaneous production of H2O2(Hatzinikolaou & Macris, 1995). Two products of GOD hydrolysis are H2O2 and gluconic acid, which may act as a bacteriocide (Crueger & Crueger, 1990; Rasiah et al., 2005; Costa et al., 2013) and acidity regulator (Nakao et al., 1997; Klein et al., 2002) to reduce the gastric pH, inhibit harmful bacteria and promote the growth of beneficial bacteria. Biagi et al. (2006) reported that feeding gluconic acid can improve the growth performance of piglets after weaning. With large-scale production of GOD by fermentation, it is possible to achieve commercial application of GOD in the feed industry (Gao et al., 2012). The researchers' previous study showed that the best dosage of GOD for growth performance improvement of pigs was 100 U/kg diet (Tang et al., 2013). Therefore, in the present study 100 U GOD/kg was supplemented into a maize-soybean meal diet. The results confirmed that GOD has positive effects on the weight gain and feed conversion efficiency of piglets.
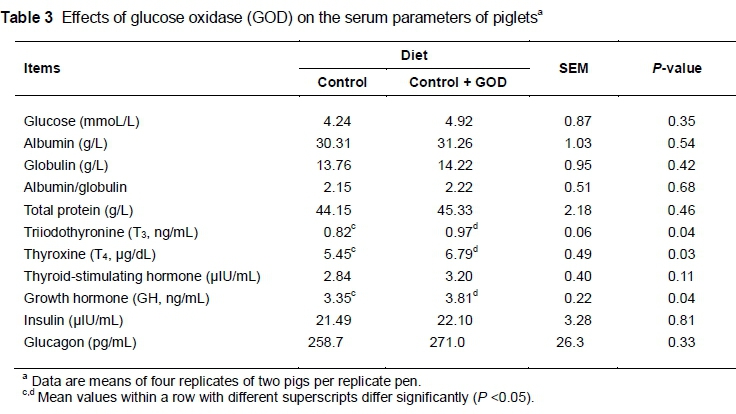
The effects of GOD supplementation in diets on blood parameters are presented in Table 3. The researchers compared 11 parameters between the control and treatment groups and found significant differences in T3, T4 and GH concentrations (P <0.05). Pigs fed GOD-supplemented diets had higher concentrations of T3 (0.97 ng/mL vs. 0.82 ng/mL), T4 (6.79 μg/dL vs. 5.45 μg/dL) and GH (3.81 ng/mL vs. 3.35 ng/mL) in the serum than those fed the control diet. Other blood parameters had a few improvements by supplementing the control diet with GOD. However, no significant differences (P >0.05) were observed.
Triiodothyronine and thyroxine are tyrosine-based hormones produced by the thyroid gland, and plasma T4 can be converted into T3(Sterling et al., 1970). Thyroid hormones have profound effects on important physiological processes such as development and growth (Visser, 1990; Namroud et al., 2010) and regulation of metabolism (Decuypere et al., 1983; 2005). Another key factor is GH, which mainly stimulates growth, cell reproduction and regeneration (Sorensen et al., 1992). Pigs are uniquely sensitive to high levels of GH, which could improve feed conversion and carcass composition (Machlin, 1972). In this experiment, serum T3, T4 and GH concentrations were increased by 13.7% after GOD supplementation. The mechanism of this effect is not clear. It might be due to gluconic acid or H2O2 produced by GOD. Brookes et al. (2005) reported that gluconic acid, as an additive, can be used as an acidity regulator, raising agent, antioxidant and chelating agent in food, feed, etc. In the current study, supplementation of GOD increased the concentrations of T3, T4 and GH in serum and promoted the growth and development of piglets, suggesting the beneficial role of GOD in piglet growth.
The effects of GOD on faecal microflora of piglets are summarized in Table 4. Pigs fed dietary GOD had lower counts of faecal Salmonella (5.3 lgcfu/g vs. 6.0 lgcfu/g, P <0.05) than pigs fed the non-GOD diet on day 35, but not on day 14 (P >0.05). The counts of total bacteria and E. coli in the treatment group were lower than those of the control group, but the differences were not significant (P >0.05). Lactobacilli counts showed no difference between the groups fed with or without GOD. In the current experiment, faecal pathogenic bacteria counts, especially Salmonella, were lower in the GOD-supplemented diet. The reason is probably that the function of GOD is to act as an antibacterial agent through the production of gluconic acid and H2O2. Biagi et al. (2006) demonstrated that feeding gluconic acid positively influenced the composition and activity of the intestinal microflora of piglets after weaning. It has been reported that sustained oxidative stress through the maintenance of low concentration of H2O2 by GOD's continued catalytic activity to be effective against pathogenic bacteria (Dobbenie et al., 1995; Wu et al., 1995). Etemadzadeh et al. (1985) reported that the ability of GOD to kill Streptococcus mutans appears to be enhanced, because the H2O2 produced by GOD acts as a useful bacteriocide. Tiina & Sandholm (1989) reported that the application of the GOD-glucose system in food products inhibited the growth of pathogens such as Salmonella, Bacillus cereus, and Yersinia. Lactobacilli are lactic acid bacteria and commonly considered to have probiotic effects in the gastrointestinal tract of animals (Zhang et al., 2011). Few studies have been performed concerning the effect of GOD on Lactobacilli. The effects of GOD on fermented sausages have shown inhibition of growth of Staphylococcus aureus, whereas GOD did not affect the growth of Lactobacilli. Lactobacilli themselves are known to produce H2O2 and therefore they are rather resistant to H2O2(Klaenhammer, 1982; Attaie et al., 1987). In this study, except for Lactobacilli, pigs fed GOD showed decreased concentrations of bacteria enumerated in contrast to the control. It is likely that GOD and their associated physicochemical effects play a beneficial role in maintaining the intestinal microflora.
Conclusion
The results showed that supplementation of GOD to diets promoted growth performance (ADG and feed efficiency), increased the concentrations of growth and development-related hormone (T3, T4 and GH), and improved the faecal microflora of growing piglets. These changes might be attributed to the functional activities of GOD in the gastrointestinal tract, which utilizes O2 and produces H2O2 and gluconic acid. Further research is needed to determine the effect of dietary GOD supplementation on other parameters of intestinal health.
Acknowledgements
This work was supported by the National High-Tech Research and Development Programme of China (863 Programme) [2013AA102803] and the 948 programme of the Ministry of Agriculture [2014-S1] and the Special Fund for Agro-Scientific Research in the Public Health of China [201403047] and China Modern Agriculture Research System [CARS-42].
References
Argenzio, R.A., 1992. Pathophysiology of diarrhea. In: Veterinary Gastroenterology. Ed: Anderson, N., Lea and Fabiger, Philadelphia. pp.163-172. [ Links ]
Attaie, R., Whalen, P.J., Shahani, K.M. & Amer, M.A., 1987. Inhibition of growth of Staphylococcus aureus during production of acidophilus yogurt. J. Food Prot. 50, 224-228. [ Links ]
Bankar, S.B., Bule, M.V., Singhal, R.S. & Ananthanarayan, L., 2009. Glucose oxidase - An overview. Biotechnol. Adv. 27, 489-501. [ Links ]
Biagi, G., Piva, A., Moschini, M., Vezzali, E. & Roth, F.X., 2006. Effect of gluconic acid on piglet growth performance, intestinal microflora, and intestinal wall morphology. J. Anim. Sci. 84, 370-378. [ Links ]
Brookes, G., Neville, C. & Kniel, B., 2005. The Global GM Market - implications for the European food chain. PG Economics Limited, UK. [ Links ]
Costa, L.B., Luciano, F.B., Miyada, V.S. & Gois, F.D., 2013. Herbal extracts and organic acids as natural feed additives in pig diets. S. Afr. J. Anim. Sci. 43, 181-193. [ Links ]
Crueger, A. & Crueger, W., 1990. Glucose transforming enzymes. In: Microbial Enzymes and Biotechnology. Eds: Fogarty, W.M. & Kelly, C.T., Elsevier, New York. pp. 177-226. [ Links ]
Decuypere, E., Scanes, C.G. & Kuhn, E.R., 1983. Effects of glucocorticoids on circulating concentrations of thyroxine (T4) and triiodothyronine (T3) and on peripheral monodeiodination in pre- and post-hatching chickens. Horm. Metab. Res. 15, 233-236. [ Links ]
Decuypere, E., Van As, P., Van der Geyten, S. & Darras, V.M., 2005. Thyroid hormone availability and activity in avian species: a review. Domest. Anim. Endocrinol. 29, 63-77. [ Links ]
Dobbenie, D., Uyttendaele, M. & Debevere, J., 1995. Antibacterial activity of the glucose-oxidase glucose system in liquid whole egg. J. Food Prot. 58, 273-279. [ Links ]
Etemadzadeh, H., Ainamo, J. & Murtomaa, H., 1985. Plaque growth-inhibiting effects of an abrasive fluoride- chlorhexidine toothpaste and a fluoride toothpaste containing oxidative enzymes. J. Clin. Periodontol. 12, 607-616. [ Links ]
Gao, Z., Li, Z., Zhang, Y, Huang, H., Li, M., Zhou, L., Tang, Y, Yao, B. & Zhang, W., 2012. High-level expression of the Penicillium notatum glucose oxidase gene in Pichia pastoris using codon optimization. Biotechnol. Lett. 34, 507-514. [ Links ]
Hampson, D.J., Hinton, M. & Kidder, D.E., 1985. Coliform numbers in the stomach and small intestine of healthy pigs following weaning at three weeks of age. J. Comp. Pathol. 95, 353-362. [ Links ]
Hatzinikolaou, D.G. & Macris, B.J., 1995. Factors regulating production of glucose-oxidase by Aspergillus-niger. Enzyme Microb. Technol. 17, 530-534. [ Links ]
Hatzinikolaou, D.G., Hansen, O.C., Macris, B.J., Tingey, A., Kekos, D., Goodenough, P. & Stougaard, P., 1996. A new glucose oxidase from Aspergillus niger: Characterization and regulation studies of enzyme and gene. Appl. Microbiol. Biotechnol. 46, 371-381. [ Links ]
Hunter, W.M., 1986. Radioimmunoassay. In: Handbook of Experimental Immunology. Volume 1. Immunochemistry. Ed: Weir, D.M., Blackwell Scientific Publications, London. pp. 1-23. [ Links ]
Jones, PH., Roe, J.M. & Miller, B.G., 2001. Effects of stressors on immune parameters and on the faecal shedding of enterotoxigenic Escherichia coli in piglets following experimental inoculation. Res. Vet. Sci. 70, 9-17. [ Links ]
, Klaenhammer, T.R., 1982. Microbiological considerations in selection and preparation of lactobacillus strains for use as dietary adjuncts. J. Dairy Sci. 65, 1339-1349. [ Links ]
Klein, J., Rosenberg, M., Markos, J., Dolgos, O., Kroslak, M. & Kristofikova, L., 2002. Biotransformation of glucose to gluconic acid by Aspergillus niger - study of mass transfer in an airlift bioreactor. Biochem. Eng. J. 10, 197-205. [ Links ]
Machlin, L.J., 1972. Effect of porcine growth hormone on growth and carcass composition of the pig. J. Anim. Sci. 35, 794-800. [ Links ]
Malherbe, D.F., Du Toit, M., Cordero Otero, R.R., Van Rensburg, P. & Pretorius, I.S., 2003. Expression of the Aspergillus niger glucose oxidase gene in Saccharomyces cerevisiae and its potential applications in wine production. Appl. Microbiol. Biotechnol. 61, 502-511. [ Links ]
Nakao, K., Kiefner, A., Furumoto, K. & Harada, T., 1997. Production of gluconic acid with immobilized glucose oxidase in airlift reactors. Chem. Eng. Sci. 52, 4127-4133. [ Links ]
Namroud, N.F., Shivazad, M., Zaghari, M. & Shahneh, A.Z., 2010. Effects of glycine-and glutamic acid supplementation to low protein diets on performance, thyroid function and fat deposition in chickens. S. Afr. J. Anim. Sci. 40, 238-244. [ Links ]
Opwis, K., Knittel, D., Schollmeyer, E., Hoferichter, P. & Cordes, A., 2008. Simultaneous application of glucose oxidases and peroxidases in bleaching processes. Eng. Life Sci. 8, 175-178. [ Links ]
Pluschkell, S., Hellmuth, K. & Rinas, U., 1996. Kinetics of glucose oxidase excretion by recombinant Aspergillus niger. Biotechnol. Bioeng. 51, 215-220. [ Links ]
Pluske, J.R., Pethick, D.W., Hopwood, D.E. & Hampson, D.J., 2002. Nutritional influences on some major enteric bacterial diseases of pigs. Nutr. Res. Rev. 15, 333-371. [ Links ]
Rasiah, I.A., Sutton, K.H., Low, F.L., Lin, H.M. & Gerrard, J.A., 2005. Crosslinking of wheat dough proteins by glucose oxidase and the resulting effects on bread and croissants. Food Chem. 89, 325-332. [ Links ]
Reed, W.M., Olander, H.J. & Thacker, H.L., 1985. Studies on the pathogenesis of Salmonella heidelberg infection in weanling pigs. Am. J. Vet. Res. 46, 2300-2310. [ Links ]
SAS, 2004. Statistical Analysis Systems User's Guide. SAS Institute Inc., Cary, N.C., USA. [ Links ]
Schwartz, K.J., 1999. Salmonellosis. In: Diseases of Swine (8th ed). Eds: Straw, B.E., Allaire, S.D., Mengeling, W.L. & Taylor, D.J., Blackwell Science, Oxford. pp. 535-551. [ Links ]
Seifu, E., Donkin, E.F. & Buys, E.M., 2004. Application of the lactoperoxidase system to improve the quality of goat milk cheese. S. Afr. J. Anim. Sci. 34, 184-187. [ Links ]
Sorensen, M.T., Chaudhuri, S., Louveau, I., Coleman, M.E. & Etherton, T.D., 1992. Growth hormone binding proteins in pig adipose tissue: number, size and effects of pGH treatment on pGH and bGH binding. Domest. Anim. Endocrinol. 9, 13-24. [ Links ]
Sterling, K., Brenner, M.A. & Newman, E.S., 1970. Conversion of thyroxine to triiodothyronine in normal human subjects. Science 169, 1099-1100. [ Links ]
Tang, H., Gao, X., Yao, B., Li, X. & Wang, X., 2013. Study on feed effects of glucose oxidase applied in piglets. China Feed 19, 21-23. [ Links ]
The Ministry of Agriculture of P R China, 2004. Feeding Standard of Swine of P. R. China. Natl. Stand. NY/T 65-2004. Beijing, P. R. China. [ Links ]
Tiina, M. & Sandholm, M., 1989. Antibacterial effect of the glucose oxidase-glucose system on food-poisoning organisms. Int. J. Food Microbiol. 8, 165-174. [ Links ]
Vaughn, S.E., 2012. Review of the third edition of the guide for the care and use of agricultural animals in research and teaching. J. Am. Assoc. Lab. Anim. Sci. 51, 298-300. [ Links ]
Visser, T.J., 1990. Importance of deiodination and conjugation in the hepatic metabolism of thyroid hormone. In: The Thyroid Gland. Ed: Greer, M.A., Raven Press, New York. pp. 255-283. [ Links ]
Wu, G., Shortt, B.J., Lawrence, E.B., Levine, E.B., Fitzsimmons, K.C. & Shah, D.M., 1995. Disease resistance conferred by expression of a gene encoding H2O2-generating glucose oxidase in transgenic potato plants. Plant Cell 7, 1357-1368. [ Links ]
Zhang, J., Deng, J., Wang, Z., Che, C., Li, Y.F. & Yang, Q., 2011. Modulatory effects of Lactobacillus salivarius on intestinal mucosal immunity of piglets. Curr. Microbiol. 62, 1623-1631. [ Links ]
Received 1 October 2014
Accepted 19 May 2015
First published online 6 February 2016
# Corresponding authors: binyao@caas.cn; xiuhuagao@126.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}