










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.45 no.5 Pretoria 2015
http://dx.doi.org/10.4314/sajas.v45i5.10
Effect of diets differing in rumen soluble nitrogen on utilization of poor-quality roughage by sheep
A.M. Mentz; W.A. van Niekerk; A. Hassen#; R.J. Coertze; B.S.Gemeda
Department of Animal & Wildlife Sciences, University of Pretoria, Pretoria 0002, South Africa
ABSTRACT
This study investigated the effects of replacing rapid-release nitrogen (N) from urea with a graded level of slow-release N (Optigen® II) source on intake, digestibility, rumen fermentation and microbial protein synthesis, when sheep were fed a poor-quality roughage diet. Five rumen cannulated wethers were used in a 5 x 5 Latin square experimental design. The treatments had various proportions of urea to Optigen® II (0 : 100, 25 : 75, 50 : 50, 75 : 25 and 100 : 0), with the same inclusion level of starch and a mineral premix on an iso-nitrogen basis. The 25% urea : 75% Optigen® II treatment showed significantly higher intakes of dry matter, organic matter, neutral detergent fibre and digestible organic matter than in the other treatments. No differences were recorded for apparent dry matter digestibility, organic matter digestibility and neutral detergent fibre digestibility among the treatments. However, sheep on the 100% Optigen® II treatment had a significantly lower apparent nitrogen digestibility. No differences were observed for ruminal pH and volatile fatty acid concentrations among the treatments, except for butyrate and isobutyrate concentrations. The butyrate concentration of the 100% Optigen® II treatment was significantly lower than the other treatments, whereas the isobutyrate concentration was significantly lower than in Treatments 2 (75% urea : 25% Optigen® II) and 5 (100% Optigen® II). The rumen NH3-N concentration of the 100% Optigen® II treatment was significantly lower than the 100% urea treatment at two and four hours after infusion. Based on biological responses, results suggest that up to 75% of urea could be replaced with Optigen® II in supplements.
Keywords: Digestibility, intake, Optigen® II, non-protein nitrogen, rumen ammonia nitrogen
Introduction
Most sheep production in South Africa is based on extensive grazing systems, where the primary feed source is poor-quality roughage. Protein is usually the first limiting nutrient, with poor-quality roughage containing less than 60 g crude protein (CP)/kg on a dry matter (DM) basis during the winter season, especially in the sourveld regions of South Africa (Currier et al., 2004). Under such conditions, ruminants are often unable to meet their nutrient requirements, because of the low nitrogen (N) and variable nutritional qualities of such feeds. Additional nutrients should therefore be supplemented strategically to minimize production losses in winter, when roughage quality and quantity are often major constraints (Köster et al., 1996; Wahrmund et al., 2007). In winter, supplementation of N could improve the intake and digestibility of poor-quality roughage feeds (Köster et al., 1996; Ferrell et al., 1999; Bohnert et al., 2002).
Urea is most commonly used in supplements as a non-protein nitrogen (NPN) source for ruminants, and is more affordable than true protein sources per unit N. The utilization of urea as an NPN source is limited because the rapid conversion of urea to ammonia may often exceed the ability of rumen bacteria to utilize the rumen NH3-N, resulting in a loss of N for synthesis of microbial protein (Löest et al., 2001; Galo et al., 2003; Golombeski et al., 2006). To overcome this limitation and reduce risks of urea toxicity in ruminants, various options have been evaluated over the years, such as creating slow-release N sources. These technologies are used to improve synchrony between available energy and N in the rumen to maximize microbial protein synthesis, and minimize the risk of NH3-N toxicity (Harrison & Karnezos, 2005; Taylor-Edwards et al., 2009b). The aim of using a slow-release N source is to provide a slow, even release of N over 24 hours, and meet the N requirements of rumen bacteria when rumen NH3-N levels are expected to be low after the ingestion of urea. By providing a sustained level of N, N utilization efficiency and microbial protein production should be increased (Harrison & Karnezos, 2005). Optigen® II (Alltech Inc. Nicholasville, KY, USA) is an example of a slow-release NPN product. It is a blended controlled urea source, coated in polyester polyurethane, which allows the diffusion of urea through micropores, and slows down the rate of N release in the rumen (ICF Consulting, 2004). The objectives of this study were to determine whether urea could be replaced fully or partially with a slow-release N source, Optigen® II, and to identify the optimum ratio of the slow-release N source to urea that synchronizes N supply with an energy source (starch) to obtain maximum intake, rumen fermentation, neutral detergent fibre (NDF) degradability and microbial protein synthesis.
Materials and Methods
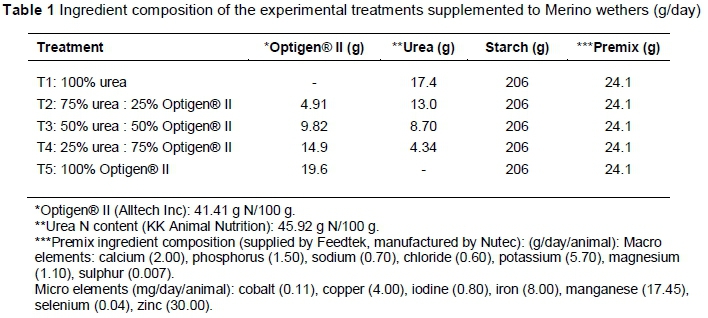
Five rumen cannulated Merino wethers with an average bodyweight (BW) of 59 ± 0.43 kg were randomly assigned to five treatments in a 5 x 5 Latin square design experiment. A preliminary intake trial was conducted to determine voluntary dry matter intake (DMI) of the poor-quality roughage before the experiment. The voluntary intake level of the roughage was used to calculate the amount of supplement (urea, Optigen® II and starch) required to meet the maintenance requirements of a 60 kg wether (NRC, 2007). These wethers had ad libitum access to milled Eragrostis curvula hay and water, while a daily supplement as per treatment was infused directly into the rumen at 08:00 and 16:00 throughout the adaptation (10 days) and experimental (8 days) periods. The daily supplement consisted of five ratios of urea to Optigen® II as experimental treatments with additional fermentable metabolizable energy (starch) and a mineral/premix (Table 1). An additional 1.8 g sulphur per day was included in each treatment to meet the recommended maintenance requirements for a 60 kg wether.
The duration of the trial was 90 days, consisting of five periods in which animals were changed to a different diet. Each experimental period lasted 18 days, of which 10 days were used for adaptation and eight for sample collection. From days 1 to 7 of the adaptation period, the animals were housed in a pen, and then moved to metabolism cages on day 8. Faecal bags were fitted to the wethers, and urine pans were fitted to the metabolism cages before the wethers were individually placed in it on day 8.
During the digestibility trial, total feed intake and total faecal output were measured over the first five days of the collection period. The last three days were used for an in situ trial to determine NDF degradability. Roughage intake was determined daily for each animal by recording the weight of the roughage offered minus the orts. Feed and orts samples were taken daily for each sheep during the five data collection periods to obtain a representative sample per treatment, and frozen at -10 °C until laboratory analysis. Total faecal output was recorded for each individual before the morning and afternoon feeding. A sample of voided faeces was taken daily for each animal, mixed and pooled over the first five days of the eight-day collection period (Köster et al., 1996; Olson et al., 1999). Two separate sub-samples of faeces were taken and frozen at -10 °C. One sub-sample was used for DM analysis at 105 °C and the other was dried at 60 °C for N and NDF analysis. Wethers were weighed before and after each experimental period. Feed, orts and faecal (dried at 60 °C) samples were ground through a 1-mm sieve before analyses. Total tract apparent digestibility for DM, organic matter (OM), CP and NDF were determined by the difference between feed nutrient intake and faecal nutrient output. The DM and ash content of the feed, orts and faecal output were determined according to standard procedures (AOAC, 2000). Crude protein was derived from the N concentration of feed, orts and faecal output, where N was determined according to the Leco instrumental manual with Leco-FP 428. The filter bag technique with ANKOM technology method 9 was used to determine NDF levels. The E. curvula hay samples were analysed for acid detergent lignin (ADL), calcium (Ca) and phosphorus (P). The ADL concentration was measured by the method of Goering & Van Soest (1970). Calcium concentration was done according to AOAC (2000) and the values read with a Perkin Elmer atomic absorption spectrophotometer as described by Giron (1973). The P concentration was analysed according to the photometric method described in AOAC (2000).
Rumen fluid samples were collected to estimate rumen fermentation parameters such as volatile fatty acid (VFA) concentration, rumen NH3-N concentration and rumen pH. These samples were collected with a suction strainer through the rumen cannulae on the first two days of the collection period. Eight samples of rumen fluid were sampled over two days by pushing the sample collection time for the next day at three-hour intervals (03:00, 06:00, 09:00, 12:00, 15:00, 18:00, 21:00, 24:00) to represent the rumen over a 24-hr period. The eight samples were pooled to make a composite sample to analyse rumen fermentation parameters. The pH was measured immediately after collection with a pH meter. To analyse rumen NH3-N, 5 mL 50% H2SO4 preservative was added to 30 mL rumen fluid (Broderick & Kang, 1980) and 4 mL 25% H3PO4 preservative was added to 20 mL ruminal fluid for VFA analysis (Webb, 1994). The rumen NH3-N and VFA samples were pooled separately over the first day of the eight-day collection period to obtain a representative sample for each individual during each collection period. Five rumen fluid samples per individual were also collected at 0, 1, 2, 3 and 4 hours after supplemental infusion to analyse rumen NH3-N concentration on the third day of each collection period. Samples were preserved with 5 mL 50% H2SO4 and frozen at -10 °C, pending further analysis.
An in situ trial was conducted after the digestibility trial (last three days of the eight-day collection period) to determine the NDF and DM disappearance of E. curvula hay, following the procedure described by 0rskov & McDonald (1979). Representative samples of hay were milled through a 2-mm sieve and 5 g was weighed into dacron bags. Dacron bags were incubated in duplicate in the rumen for 0, 2, 4, 8, 16, 24, 48 and 72 hours per sheep (16 bags per sheep). Two bags were taken out at each time interval, washed until the water was clear, and frozen at -10 °C. After incubation, the dacron bags were defrosted and dried at 60 °C (NRC, 2007). The residues were analysed for NDF and DM.
Urine samples were collected to estimate rumen microbial protein (MCP) synthesis by analysing the urine samples from each animal during each period for purine derivatives (PD) (Chen & Gomes, 1992). Two methods were used to estimate the PD. The preferred method for PD analysis (total PD analysis) was used (Chen & Gomes, 1992), as well as the spot urine collection method (Chen et al., 1995). Purine derivatives in the urine are positively related to microbial nitrogen flow, because of the production of exogenous purines by rumen micro-organisms through the degradation and absorption of microbial nucleic acids (Chen & Gomes, 1992; Chen et al., 1995; Moorby et al., 2006). The spot urine method is an alternative to the preferred PD analysis method. It is based on the assumption that creatinine in the urine is excreted at a constant rate and is a function of metabolic body weight (Chen et al., 1995; Cetinkaya et al., 2006; Moorby et al., 2006). The PD : creatinine ratio is highly correlated to PD excretion, and could therefore be used to estimate MCP supply. High-performance liquid chromatography was used to analyse the urine samples for purine derivatives (Chen & Gomes, 1992) and a quantichrome creatinine essay kit and spectrophotometer were used to analyse for creatinine. Urine of each sheep was collected with a urine pan and preserved with 50% H2SO4. A container with 40 mL 50% H2SO4 was used to collect the urine. After each daily collection, the pH of the urine was reduced to 3, by adding more 50% H2SO4 and the amount of urine was recorded. The volume of 50% H2SO4 added depended on the amount of urine collected and the pH of the urine each day. Then the urine was diluted with tap water up to 4000 mL. A sub-sample (50 mL) was taken from each animal's diluted urine (Chen & Gomes, 1992). The urine samples were pooled to obtain five representative samples.
Data from the laboratory were subjected to analysis of variance using the Proc GLM model (Statistical Analysis System, 2006) for a Latin square design model. The model of 0rskov & McDonald (1979) was fitted to determine NDF and DM degradation. The experiment was approved by the Ethics Committee of University of Pretoria (Ref No: EC004-10).
Result and Discussion
The E. curvula hay fed was of poor quality (34.5 g CP/kg), far below 60 g CP/kg and the CP requirements of a 60 kg sheep at maintenance (79 g CP/kg) (NRC, 2007). Apart from low CP, it had high NDF (834.8 g/kg) and ADL (102.8 g/kg) concentrations. The Ca and P concentrations of the E. curvula hay were 2.2 g and 0.9 g/kg, respectively.
No differences were observed between the 100% urea and 100% Optigen® II treatments for intake (Table 2). A combination of urea and Optigen® II (Treatment 4) improved most of the intake variables (P <0.05), because higher DMI, organic matter intake (OMI) and total roughage intake were recorded for Treatment 4 compared with Treatments 1, 3 and 5 (P <0.05). The NDF and digestible organic matter intake (DOMI) of Treatment 4 were higher than in Treatment 5 (P <0.05). The higher intake observed with Treatment 4 could be owing to improved fermentation associated with better synchrony of N release from Optigen® II with the slow-released fermentable energy from fibre, as well as the fast N release from urea at early stages that synchronized with the fast fermentable energy from starch. A similar finding was reported by Ribeiro et al. (2011), who suggested that Optigen® II treatments might have provided sufficient N for rumen fermentation over an extended period. In contrast with this study and that of Ribeiro et al. (2011), Wahrmund et al. (2007) found no difference in DMI between the control, urea and Optigen® II treatments in a study that compared Optigen® II as a NPN source instead of urea in Bahia grass-fed animals.
The apparent digestibility of DM, OM and NDF was not affected by the treatments, except for apparent N digestibility (P <0.05) (Table 2). Apparent N digestibility was lower for Treatment 5 than Treatments 1, 2, 3 and 4 (P <0.05). This might be ascribed to the slow release of NH3-N in the rumen from Optigen® II or it might be due to asynchrony between the available fermentable metabolizable energy (FME) and N from Optigen® II to the rumen microbes. Consequently, some of the Optigen® II might have left the rumen without being hydrolysed or incorporated into rumen microbial protein. Lack of difference in terms of apparent digestibility of DM, OM and NDF among treatments was reported by Taylor-Edwards et al. (2009b). These authors reported a significantly lower N digestibility with the polymer-coated slow-release urea (SRU) treatment than for the urea treatment. In contrast with the present findings (Table 4), Ribeiro et al. (2011) found no differences in N digestibility between urea and Optigen® II treatments. According to the findings of this study, it could be suggested that post-ruminal N concentration would increase when Optigen® II is used to replace urea completely in a supplement, and therefore increased faecal N levels could be expected (Taylor-Edwards et al., 2009a).
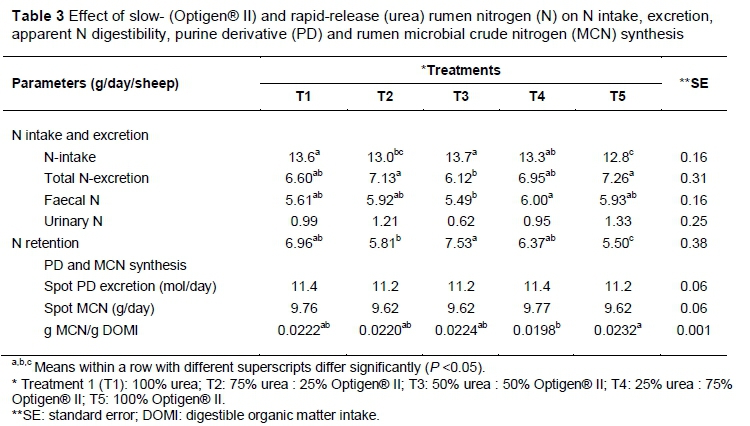
Significant differences were observed for N intake, N excretion, faecal N and N retention (P <0.05) (Table 3). The N intake of Treatment 5 was lower than in the other treatments (P <0.05). However, Treatments 3 and 5 had higher N-excretions (P <0.05) than the other treatments, with no differences in urinary N excretion among treatments. Nitrogen retention was positive for all the treatments, with Treatment 3 having the highest value (Table 3), but differed (P <0.05) only from Treatments 4 and 5. The higher N retention of Treatment 3 could be explained by the observed higher N intake and lower N excretion for this treatment compared with Treatment 5. The observed low N retention for Treatment 5 could be related to high N excretion, low apparent N digestibility and the low N intake. In contrast with the findings of this study, Taylor-Edwards et al. (2009b) found higher faecal N for slow-release urea (SRU) compared with urea. These authors suggested that these findings could be possible only if some of the SRU passed through the digestive tract without being hydrolysed in the rumen. In contradiction, Galo et al. (2003) found that the control diet had a higher faecal N excretion compared with treatments containing SRU, and postulated that some damage to the coating might have occurred. In line with this study, Bourg et al. (2009) recorded no differences between urea and Optigen® II treatments for faecal and urinary N excretion. The present study showed that treatments containing higher levels of Optigen® II had higher N excretion (faecal and urinary) values (Table 4), suggesting that N released from Optigen® II might not have been captured in microbial crude protein (MCP) during the early hours of fermentation (Galo et al., 2003; Taylor-Edwards et al., 2009b).
No differences were found between the treatments in terms of PD excretion, PD absorption and microbial crude nitrogen (MCN) supply with the total PD method (Chen & Gomes, 1992). In addition, analysis of urine with the spot urine method resulted in no differences (P >0.05) for PD and MCN. However, a difference was observed between Treatments 4 and 5 for the efficiency of MCN supply (g MCN/g DOMI). Results from this study were comparable with those of Chen et al. (1995). According to results of this study it seems that Optigen® II might improve the efficiency of MCN supply. According to Chen et al. (1995), DMI is positively correlated to PD excretion. Chen et al. (1995) reported that the diets with the highest DMI had the highest PD excretion. A similar tendency was observed in the present study where Treatment 4 had the highest DMI (P <0.05) and the highest PD excretion (P >0.05).
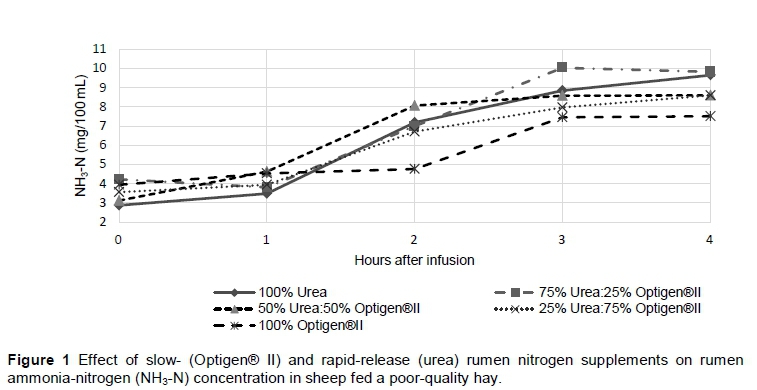
Although the average rumen NH3-N concentrations did not differ (P >0.05) among treatments, a significant treatment x time interaction was recorded for all treatments (P <0.05). Treatments containing higher concentrations of Optigen® II (Treatments 4 and 5) had lower (P <0.05) rumen NH3-N concentrations at two and four hours after feeding (Figure 1). Similar results were recorded by Ribeiro et al. (2011), in which the authors found that urea and urea/Optigen® II treatments had higher rumen NH3-N concentrations than Optigen® II alone. The rumen NH3-N concentrations of Treatments 1, 2, 3 and 4 at two hours were higher than at one hour after feeding (P <0.05). Rumen NH3-N concentration of Treatment 5 started to increase only between two and three hours after feeding (P <0.05), suggesting that 100% Optigen® II indeed has a slower ammonia-release rate than the other treatments containing urea. However, a slower ammonia-release rate might not have been the only reason for a lower rumen NH3-N concentration for treatments containing higher levels of Optigen® II. This might also be partly because of improved utilization of NH3-N by rumen bacteria. A continuous release of NH3-N in the rumen should improve fibre digestion and the utilization of the diet owing to prolonged availability of N to rumen bacteria (Wahrmund et al., 2007). The lowest rumen NH3-N concentrations were observed at 0 hours (just before feeding), with Treatment 1 having the lowest (2.88 mg/100 mL) concentration. However, it was still above the suggested limiting concentration (2 mg NH3-N/100 mL) for microbial growth and digestion to fulfil the maintenance requirements of sheep (Satter & Slyter, 1974). Therefore, rumen NH3-N concentrations should not have had a negative effect on microbial growth and digestion.
There were no differences among treatments in terms of average rumen pH (P <0.05). In the current study there were no definite tendencies that indicated 100% Optigen® II or a combination of Optigen® II/ urea had lower pH levels than 100% urea alone. The average pH over 24 hours was similar among treatments. Differences in pH were observed between Treatments 1 and 2 at 06:00 and between Treatment 1 and Treatments 2, 3 and 5 at 21:00 (P <0.05) (Figure 2). Taylor-Edwards et al. (2009b) found similar results in terms of pH between urea and SRU treatments. In contrast, Wahrmund et al. (2007), reported that rumen pH tended to be affected by urea source, where Optigen® II tended to have a lower pH than the control and urea treatments. The pH of all treatments was never below 6.0 for a prolonged period which would have had a negative influence on fibre digestion (Mould et al., 1983).
No differences (P >0.05) were observed among treatments for the ruminal acetate : propionate ratio and the other individual rumen-produced VFA, except for butyrate and isobutyrate (Table 4). In accordance with previous studies (Taylor-Edwards et al., 2009b), VFA concentration was not affected differently by urea and/or a slow-release N source. It is uncertain what caused the difference (P <0.05) in butyrate concentrations. According to Heldt et al. (1999) and Olson et al. (1999), supplemental energy is responsible for increased butyrate concentration in the rumen, and not the RDP source.
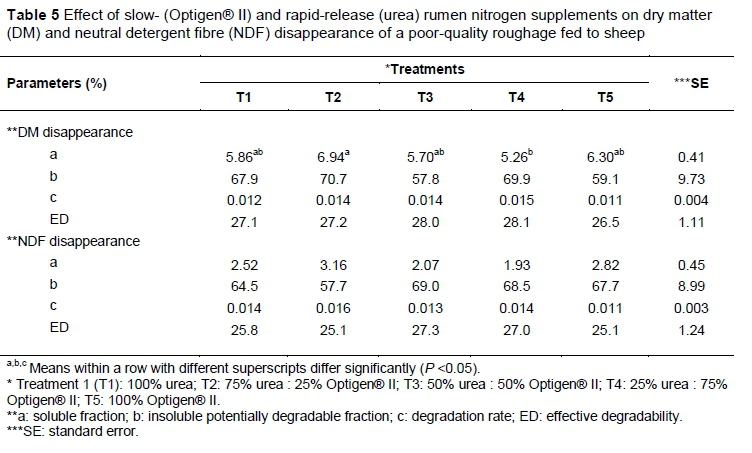
The DM and NDF in situ disappearance of the E. curvula hay did not differ among treatments (Table 5). The only difference (P <0.05) was in the soluble fraction of DM disappearance of Treatments 2 and 4. Similar results were reported by Ribeiro et al. (2011) for the insoluble potentially degradable fraction (b), degradation rate (c) and effective degradability (ED) of both the DM and NDF disappearances of the poor-quality hay (Bracharia humidicola cv Llanero). The lack of treatment effect on NDF disappearance of poor-quality E. curvula hay in the present study might be due to the optimization of ruminal NDF degradation. The average rumen NH3-N was below 8 mg/100 mL for all the treatments over a 24-hour period, but after three hours of infusion, the rumen NH3-N concentrations increased above the 8 mg/100 mL level that was suggested for fibrolytic enzymes to optimize NDF degradation (Detmann et al., 2009). It is suggested that the degradability of DM and NDF is dependent not only on the rumen NH3-N concentration, but also on the fermentability of the diet and the chemical and physical structures of the diet (Boucher et al., 2007). No lag time was observed for DM and NDF degradability. Similarly, Ribeiro et al. (2011) found no lag time for various treatments (urea, urea + Optigen® II and Optigen® II) when Llanero hay was fed.
Conclusion
In this study, a 100% Optigen® II treatment gave lower N retention, lower N intake, higher N-excretion and lower rumen NH3-N concentrations than the other treatments, However, the 100% Optigen® II treatment was more efficiently utilized by ruminal micro-organisms for the synthesis of MCP. A level of up to 75% of urea could be replaced with Optigen® II as a NPN source without negative effects on intake, digestibility, rumen fermentation and microbial protein synthesis. The primary advantage of replacing feed grade urea with Optigen® II would be an improvement in roughage intake in addition to a reduction in potential hazards such as ammonia toxicity, which are associated with the use of urea.
Acknowledgement
The authors are grateful to the Protein Research Foundation and Feedtek for funding the first author in her research study. Feedtek also provided the mineral premix and deserves our acknowledgment. Additional research funding was made available from an NRF and IFS grant.
References
AOAC, 2000. Official Methods of Analysis (15th ed.). Association of Official Analytical Chemists, Arlington, VA., USA. [ Links ]
Bohnert, D.W., Schauer, C.S. & Delcurto, T., 2002. Influence of rumen protein degradability and supplementation frequency on performance and nitrogen use in ruminants consuming low-quality forage: cow performance and efficiency of nitrogen use in wethers. J. Anim. Sci. 80, 1629-1637. [ Links ]
Boucher, S.E., Ordway, R.S., Whitehouse, N.L., Lundy, F.P., Kononoff, P.J. & Schwab, C.G., 2007. Effect of incremental urea supplementation of a conventional corn silage based diet on ruminal ammonia concentration and synthesis of microbial protein. J. Dairy Sci. 90, 5619-5633. [ Links ]
Bourg, B.M., Wickersham, T.A., Tedeschi, L.O. & Tricarico, J.M., 2009. Effects of a slow-release urea product on performance, carcass characteristics, and nitrogen balance of steers fed steam-flaked corn. J. Anim. Sci. 90, 3914-3923. [ Links ]
Broderick, G.A. & Kang, J.H., 1980. Automated simultaneous determination of ammonia and total amino acids in ruminal fluid and in vitro media. J. Dairy. Sci. 63, 64-75. [ Links ]
Cetinkaya, N., Yaman, S. & Baber, N.H.O., 2006. The use of purine derivatives/creatinine ratio in spot urine samples as index of microbial protein supply in Yerli Kara crossbred cattle. Livest. Sci. 100, 91-98. [ Links ]
Chen, X.B. & Gomes, M.J., 1992. Estimation of microbial protein supply to sheep and cattle based on urinary excretion of purine derivatives: An overview of the technical details. Technical report, Rowett Research Institute, Bucksburn, Aberdeen AB2 9SB, UK. [ Links ]
Chen, X.B., Mejia, A.T., Kyle, D.J. & 0rskov, E., 1995. Evaluation of the use of the purine derivative: creatinine ratio in spot urine and plasma samples as an index of microbial protein supply in ruminants: studies in sheep. J. Agric. Sci., Camb. 125, 137-143. [ Links ]
Currier, T.A., Bohnert, D.W., Falck, S.J., Schauer, C.S. & Bartle, S.J., 2004. Daily and alternate-day supplementation of urea and biuret to ruminants consuming low-quality forage: II. Effects on site of digestion and microbial efficiency in steers. J. Anim. Sci. 82, 1518-1527. [ Links ]
Detmann, E., Paulino, M.F., Mantovani, H.C., Valadares Filho, S.C., Sampaio, C.B., De Souza, M.A., Lazzarini, I. & Detmann, K.S.C, 2009. Parameterization of ruminal degradation in low-quality tropical forage using Michaelis-menten kinetics. Livest. Sci. 126, 136-146. [ Links ]
Ferrell, C.L., Kreikemeier, K.K. & Freetly, H.C., 1999. The effect of supplemental energy, nitrogen and protein on feed intake, digestibility and nitrogen flux across the gut and liver in sheep fed low quality forage. J. Anim. Sci. 77, 3353-3364. [ Links ]
Galo, E., Emanuele, S.M., Sniffen, C.J., White, J.H. & Knapp, J.R., 2003. Effects of a polymer-coated urea product on nitrogen metabolism in lactating Holstein dairy cattle. J. Dairy Sci. 86, 2154-2162. [ Links ]
Giron, H.C., 1973. Comparison between dry ashing and wet digestion in the preparation of plant material for atomic absorption analysis. Atomic Absorption Newsletter 12, 28. [ Links ]
Goering, H.K. & Van Soest, P.J., 1970. Forage fibre analysis (Apparatus, reagents, procedures and some applications). Number Agricultural handbook no.379. US Dept. of Agriculture, Washington DC., USA. [ Links ]
Golombeski, G.L., Kalscheur, K.F., Hippen, A.R. & Schingoethe, D.J., 2006. Slow-release urea and highly fermentable sugars in diets fed to lactating dairy cows. J. Dairy Sci. 89, 4395-4403. [ Links ]
Harrison, G.A. & Karnezos, T.P., 2005. Can we improve the efficiency of nitrogen utilization in the lactating dairy cow? In: Recent Advances in Animal Nutrition in Australia. School of Rural Science and Agriculture, University of New England. Vol. 15. pp. 146-154. [ Links ]
Heldt, J.S., Cochran, R.C., Mathis, C.P., Woods, B.C., Olson, K.C., Titgemeyer, E.C., Nagaraja, T.G., Vanzant, E.S. & Johnson, D.E., 1999. Effects of level and source of carbohydrate and level of degradable intake protein on intake and digestion of low-quality tallgrass-prairie hay by beef steers. J. Anim. Sci. 77, 2846-2854. [ Links ]
ICF Consulting, 2004. Potential concentrations of optigen residues in the environment and a comparison to similar residues from other sources. Technical report, Alltech Inc., Catnip Hill Pike, Nicholasville, KY. [ Links ]
Köster, H.H., Cochran, R.C., Titgemeyer, E.C., Vanzant, E.S., Abdelgadir, I. & St-Jean, G., 1996. Effect of increasing degradable intake protein on intake and digestion of low-quality, tallgrass-prairie forage by beef cows. J. Anim. Sci. 74, 2473-2481. [ Links ]
Löest, C.A., Titgemeyer, E.C., Drouillard, J.S., Lambert, B.D. & Trater, A.M., 2001. Urea and biuret as nonprotein nitrogen sources in cooked molasses blocks for steers fed prairie hay. Anim. Feed Sci. Technol. 94, 115-126. [ Links ]
Moorby, J.M., Dewhurst, R.J., Evans, R.T. & Danelon, J.L., 2006. Effects of dairy cow diet forage proportion on duodenal nutrient supply and urinary purine derivative excretion. J. Dairy Sci. 89, 3552-3562. [ Links ]
Mould, F.L., Ørskov, E.R. & Mann, S.O., 1983. Associative effects of mixed feeds. Effects of type and level of supplementation and the influence of the rumen fluid pH on cellulolysis in vivo and dry matter digestion of various roughages. Anim. Feed Sci. Technol. 10, 15-30. [ Links ]
NRC, 2007. National Research Council. In: Nutrient Requirements of Sheep. National Academy Press, Washington D.C., USA. [ Links ]
Olson, K.C., Crochan, R.C., Jones, T.J., Vanzant, E.S., Titgemeyer, E.C. & Johnson, D.E., 1999. Effects of ruminal administration of supplemental degradable intake protein and starch on utilization of poor quality warm-season grass hay by beef steers. J. Anim. Sci. 77, 1016-1025. [ Links ]
Ørskov, E.R. & McDonald, P., 1979. The estimate of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J. Agric. Sci., Camb. 92, 499-507. [ Links ]
Ribeiro, S.S., Vasconcelos, J.T., Morais, M.G., Itavo, C.B.C.F. & Franco, G.L., 2011. Effects of ruminal infusion of a slow-release polymer-coated urea or conventional urea on apparent nutrient digestibility, in situ degradability, and rumen parameters in cattle fed low-quality hay. Anim. Feed Sci. Technol. 164, 53-61. [ Links ]
SAS, 2006. Statistical Analysis System user's guide: Statistics version 9.1.3. SAS institude Inc, Cary, N.C., USA. [ Links ]
Satter, L.D. & Slyter, L.L., 1974. Effect of ammonia concentration on rumen microbial production in vitro. Br. J. Nutr. 32, 199-208. [ Links ]
Taylor-Edwards, C.C., Elam, N.A., Kitts, S.E., Mcleod, K.R., Axe, D.E., Vanzant, E.S., Kristensen, N.B. & Harmon, D.L., 2009a. Influence of slow-release urea on nitrogen balance and portal-drained visceral nutrient flux in beef steers. J. Anim. Sci. 87, 209-221. [ Links ]
Taylor-Edwards, C.C., Hibberd, G., Kitts, S.E., Mcleod, K.R., Axe, D.E., Vanzant, E.S., Kristensen, N.B. & Harmon, D.L., 2009b. Effects of slow-release urea on ruminal digesta characteristics and growth performance in beef steers. J. Anim. Sci. 87, 200-208. [ Links ]
Wahrmund, J., De Araujo, D.V., Hersom, M. & Arthington, J., 2007. Evaluation of Optigen® II as a source of rumen degradable protein for mature beef cows. Technical report, Florida beef report, Florida, USA. [ Links ]
Webb, E.C., 1994. Synthesis of long chain fatty acids in ruminants and their effects on meat quality. PhD thesis, University of Pretoria, Pretoria. [ Links ]
Received 16 September 2014
Accepted 15 October 2015
First published online 14 December 2015
# Corresponding author: Abubeker.hassen@up.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}