










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.45 n.5 Pretoria 2015
http://dx.doi.org/10.4314/sajas.v45i5.4
Genetic analysis of pre-weaning survival and inbreeding in the Boxer dog breed of South Africa
B.E. MostertI, #; E. van Marle-KösterI; C. VisserI; M. OosthuizenII
IDepartment of Animal and Wildlife Sciences, University of Pretoria, Pretoria, 0002, South Africa
IIDepartment of Veterinary Tropical Diseases, University of Pretoria, Onderstepoort, 0110, South Africa
ABSTRACT
Members of the Federation of Boxer Clubs in Southern Africa (FBCSA) have participated in official recording for many years. The aim of the study was to estimate genetic parameters and trends for pre-weaning survival and to assess inbreeding in the South African Boxer, based on performance and pedigree data. Fitness records were included from 2138 litters recorded between 1988 and 2012 and pedigree information of 32 029 Boxer dogs recorded from 1950 to 2012. The average litter size was 6.14 ± 2.43. More male puppies were born per litter (3.23 ± 1.79) than female puppies (2.91 ± 1.72), with an increase in the proportion of males in larger litter sizes. Heritability values of 0.23 ± 0.08 and 0.25 ± 0.08 were estimated for litter size (LS) and number of puppies alive at two weeks after birth (NA), respectively. Litters larger than five tended to have more puppies born dead or with low survivability. Number of puppies born dead or that died within two weeks after birth increased from 0.43/litter at first parity to 0.82/litter at fifth parity, with a substantial increase of 2.25/litter with low survivability at seventh parity. Average inbreeding levels of the Boxer breed were above 10%, with an inbreeding rate of 0.14%/year since 1982. These results indicate that FBCSA should focus on lowering the levels of inbreeding by avoiding mating closely related animals. The complete recording of fitness traits is encouraged for effective genetic management of the breed.
Keywords: Fitness, heritability, litter size, number born alive, survivability
Introduction
The Boxer dog is descended from the ancient eastern Molossor dogs, which were used for fighting and protection. The modern Boxer dates back to 1890 when the German Bullbeiser (now extinct) and the English Bulldog were crossed in Germany (Grandjean & Vaisarre, 2004). Breed standards for the Boxer were formalized in the early nineteenth century. Since then it has become a popular breed worldwide for protection and companionship.
The Federation of Boxer Clubs in Southern Africa (FBCSA) was founded in 1960 and became responsible for organizing the breed, setting breed standards and promoting genetic improvement of the breed (http://www.fbcsa.net/). FBCSA is a recognized breed society for dogs under the Animal Improvement Act No 62 (1998) and takes part in official recording of traits important to the breed. The Southern African Boxer population currently comprises about 2650 recorded dogs (Zeiler et al., 2013).
Recording in most dog breeds is limited to pedigrees. Breeding objectives focus on conformation, subjective appearance and breed standards relating to the function of the breed (Asher et al., 2009; Stock et al., 2012). Because of the importance of conformation, body measurements have been studied in breeds, for instance the German shepherd, where traits such as wither height and bodyweight have been found to be moderately heritable (Stock et al., 2012). In a number of studies, conformation traits have been examined pertaining to hip and elbow dysplasia (Malm et al., 2008; Krontveit et al., 2010; Lewis et al., 2010; Meyer et al., 2012), as these are common disorders in several dog breeds.
Breeding and mating strategies practised by breeders influence the structure of the breed and genetic diversity directly. Canine breeding is usually characterized by small population sizes and mating decisions based on desired phenotypes (appearance and behaviour) without considering relatedness (Glazewska, 2008). This results in inbreeding depression that adversely affects fitness and fertility traits (Leroy et al., 2006; Urfer, 2009; Cecchi et al., 2013). Unfortunately, because puppies are sold at a young age, breeders are often unaware of these consequences. Genetic diversity studies that assess inbreeding have been conducted for a number of dog breeds based on pedigree data (Leroy et al., 2006; Glazewska, 2008; Urfer, 2009; Voges & Distl, 2009). These studies emphasise the challenges that dog breeds face in genetic diversity because of their historical development, with a limited number of founder animals and subsequent inbreeding that accrues partly because of incomplete or inconsistent pedigrees.
Phenotypic measurements of fertility and growth traits are not routinely recorded by dog breeders and estimates of genetic parameters are limited. FBCSA has been recording litter and pedigree information of the Boxer breed for many years. In 1999, they joined SA Stud Book as their service provider, enabling the first genetic analysis of fitness traits as recorded by FBCSA and their first assessment of inbreeding. The aim of the study was therefore to estimate genetic parameters and trends for survival and to assess inbreeding in the Southern African Boxer breed based on performance and pedigree data as captured on the Livestock Operational and Genetic Information Exchange database system of SA Stud Book (Logix).
Material and Methods
Litter and pedigree information from FBCSA members that participate in official recording were downloaded from Logix. For this study, fitness traits were considered, based on 2138 litters recorded between 1988 and 2012 and pedigree information of 32 029 Boxer dogs recorded from 1950 to 2012. These criteria were applied for editing litters that consisted of up to 10 puppies from the first seven parities: litters with no birth date; litters with no puppies; litters with unknown breeders; litters where the birth date of the dam was later than the birth date of her litter; litters where more puppies died than were born alive and the age of the dam at the birth of her litter was below 300 days. Therefore 1, 18, 30, 11, 8 and 4 records, respectively, were discarded. The edited data file consisted of 2096 litters (some litters were discarded based on more than one criteria) from 1096 dams, representing 717 sires and 423 breeders. Animals with unknown birth dates (25% of animals in the pedigree) were allocated dates of birth later than those of their parents and/or earlier than those of their progeny, assuming a generation interval of two years. For the animals with known birth dates, 0.5% of the data were discarded due to faulty birth dates that resulted in dogs being born before their parents. The edited pedigree file consisted of 31 877 animals, the progeny of 2133 sires and 3631 dams.
Descriptive statistics were calculated for traits that indicated pre-weaning survival, which included average litter size, number of puppies born per litter and born alive, and the distribution of litter size over parities and seasons.
Heritability estimates for litter size (LS) and number of puppies alive at two weeks after birth (NA) were estimated as a trait of the dam with the genetic software package VCE6 (Groeneveld et al., 2010), using the edited dataset and a pedigree file traced back for three generations. The best linear unbiased prediction (BLUP) animal model was used to estimate heritability for LS and NA:

where yijklmn = LS or NA analysed as a trait of the nth dam
cgi = fixed effect of contemporary group (breeder x birth year; 1118 levels)
agej = linear regression of age of dam on LS or NA
ssirek = fixed effect of service sire (717)
parl = fixed effect of parity of the dam (7 levels)
pem = permanent environmental effect to account for repeated records of the trait in the different parities
an = additive genetic effect of the dam
eijklmn = random residual error
Assumptions of the variances in the model were V(a) = Aa2a, V(pe) = Ia2pe, V(e) = Ia2e, where A is the numerator relationship matrix, I the identity matrix and σ2a, σ2pe and σ2ethe direct additive genetic variance, permanent environmental variance and residual error variance. Because the age of the dam showed wide variations for all parities and an overlap of age of the dam among parities occurred, a linear regression of age of the dam on the traits and a parity effect were fitted.
Breeding values for LS and NA were estimated using PEST2 (Groeneveld et al., 2001) by iteration on data, with a relaxation factor and a stopping criterion set at 0.64 and 0.00001, respectively. Genetic trends were calculated by averaging breeding values per year of birth for dams with litters. Linear regressions were fitted on the genetic trends using SAS (2012) to describe the rate of genetic change for birth year periods 1983 - 2000 and 2000 - 2010.
Inbreeding coefficients were calculated for all animals in the edited pedigree file using the algorithm of Aguilar & Mistzal (2008). This algorithm allocates an average inbreeding level (according to the generation involved) to unknown parents for more reliable estimates of inbreeding compared with conventional methods, which assume that unknown parents are unrelated. Both parents were known for 23 386 dogs (73%), 200 dogs had only a known sire, 118 dogs had only a known dam, and 8325 dogs (26%) had no parental information. Rate of inbreeding per year was determined as:
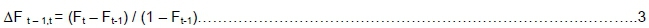
where ΔF t - 1t is the rate of inbreeding between years t-1 and t, Ft-1 and Ft are the average inbreeding levels at years t-1 and t, respectively.
The effect of inbreeding on LS and NA was estimated by fitting individual inbreeding coefficients as a linear covariate in the above model, using PEST2 (Groeneveld et al., 2001).
Mortalities of Boxer puppies are recorded up to 14 days after birth. The effects of litter size and parity on puppies born dead or dead at an early age after birth were investigated separately by acquiring least squares means (LS means) from this fixed effects model, using Proc GLM of SAS Version 4.3 (2012):
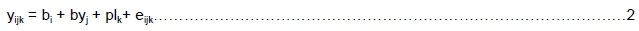
where yijk= number of dead puppies for breederi, year of birthjand parity or litter sizek
bi= fixed effect of breeder (423 levels)
byj= fixed effect of birth year of litter (25 levels)
plk= fixed effect of litter size (10 levels) (when investigating parity) or parity (7 levels) (when investigating litter size)
eijk= random residual error
The LS means were averaged for every level of parity from the analysis where litter size was included as a fixed effect and plotted to assess the influence of parity on number of dead puppies. The same was done for LS means of litter size, where parity was included as fixed effect. This effect of litter size on number of dead puppies per litter was plotted separately for each parity to assess a parity x litter size effect on number of dead puppies/litter.
Results
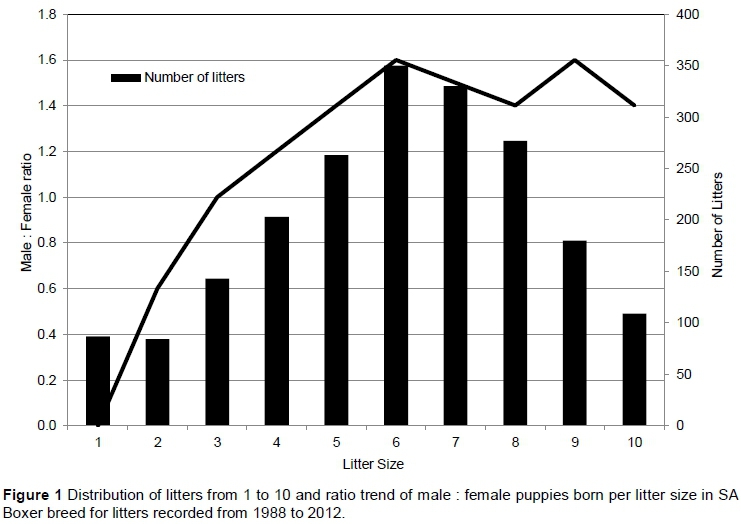
Litter size in the Boxer breed ranged between 1 and 14 puppies per litter (evaluated in the unedited dataset), with an average litter size of 6.14 ± 2.43. On average, more male puppies were born per litter (3.23 ± 1.79) than female puppies (2.91 ± 1.72), with a significant (P = 0.0001) increase in number of males relative to number of females born in larger litter sizes. Interestingly, all litters consisting of only one puppy (87) contained only males. In Figure 1, the ratio of males to females born increased (line) up to the average litter size of 6, after which it stabilized at a ratio of 1.4 - 1.6.
Litters were distributed over 10 parities, with 53% of all litters being from parity 1, and only 5% from parities greater than 4. The average number of parities for the breed is 1.9 ± 1.3. Most litters were born in 1993 (9%), with 56% of all litters being born from 1992 to 1999, 42% since 2000, and 2% of litters in 2012. However, at the time of data withdrawal, all information for litters born in 2012 had not been captured.
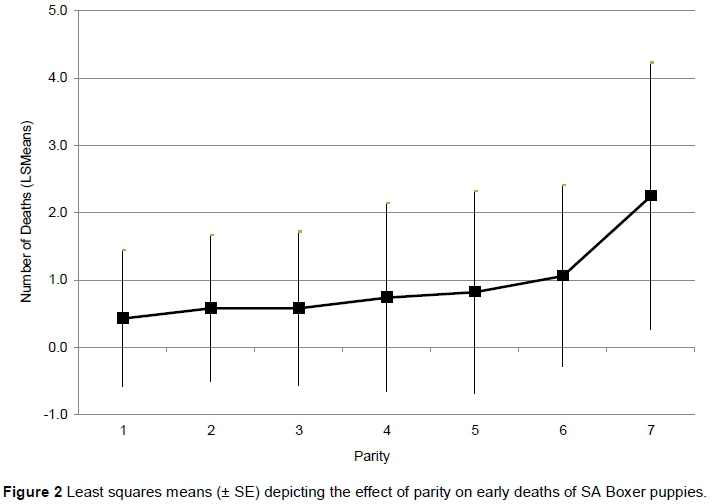
Most litters were born in the winter months (May - July), and the fewest in December. Although month did not have a significant effect (P = 0.055) on the average number of puppies/litter that died within the first two weeks of birth, there is a tendency for more deaths in colder months (May - August: 0.53 ± 1.10 puppies/litter), with the lowest average deaths/litter (0.39 ± 0.87 puppies/litter) occurring in February. Figure 2 presents the effect of parity on early deaths of puppies for the first seven parities. This effect is corrected for differences among breeders, years and litter sizes, and therefore indicates the effect of parity only on the early deaths of puppies. The relationship of parity with early deaths per litter can be described by a curvilinear function. The number of puppies born dead or that died at an early stage after birth increases gradually up to the fifth parity. A much steeper increase can be observed from the fifth parity onwards. LS means of number of dead puppies/litter were 0.43 ± 1.01, 0.82 ± 1.09 and 2.25 ± 1.98 puppies/litter for parities 1, 5 and 7, respectively.
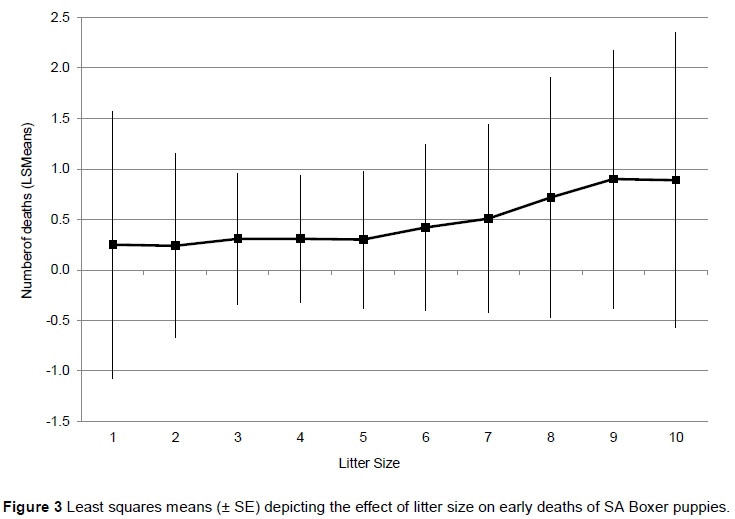
The effect of litter size on the number of puppies born dead or that died shortly after birth is indicated in Figure 3. This effect was corrected for differences among breeders, years and parities, and therefore indicates the effect of litter size only on early deaths. The number of dead puppies rises only slightly with increasing litter size up to a litter size of 5, followed by a much steeper escalation with litter sizes larger than 5. LS means for early deaths were 0.24 ± 0.91, 0.30 ± 0.68 and 0.90 ± 1.28 puppies/litter for litter sizes of 2, 5 and 9, respectively.
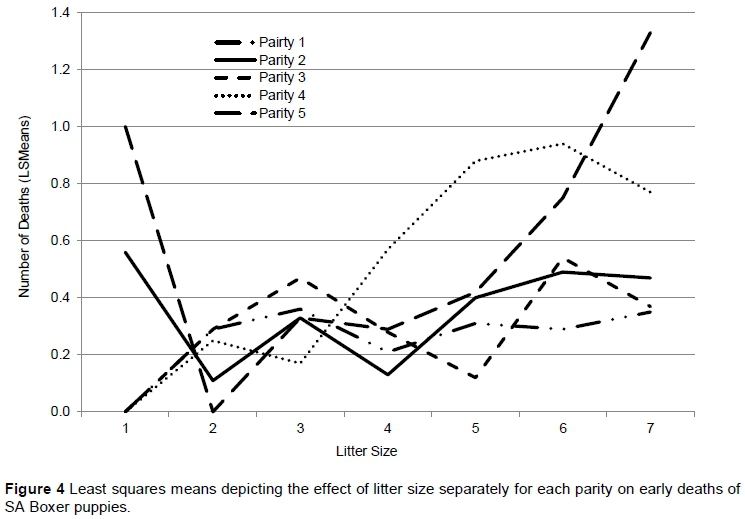
The effect of litter size on early deaths is indicated separately for each parity in Figure 4. The first parity had a smoother curve than subsequent parities, and shows the lowest incidence in early deaths with increasing litter sizes. The first three parities have a relatively similar effect on early deaths with increasing litter size, compared with later parities. Curves for the fourth and fifth parities in particular show fluctuations because few records are available for estimation. Early deaths in parities 4 and 5 increase at a much higher rate for litter sizes of four and five, compared with the earlier parities. The unparallel behaviour of the lines is an indication of litter size x parity interactions for the early deaths of puppies/litter.
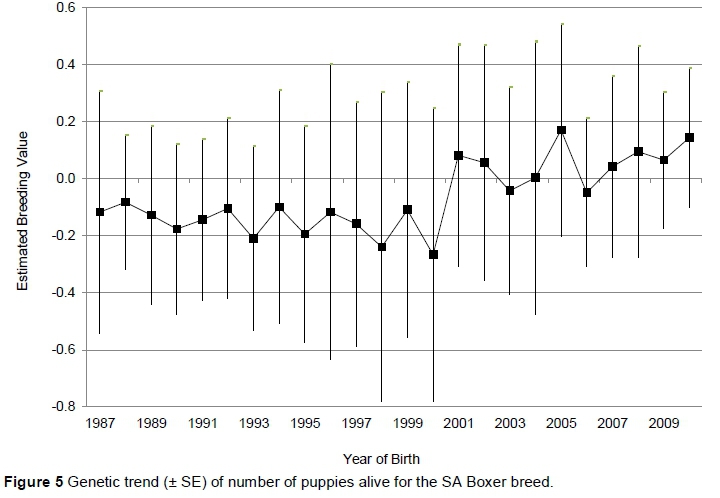
Heritability estimates of 0.23 ± 0.08 (additive variance 1.255 ± 0.540) and 0.25 ± 0.08 (additive variance 1.350 ± 0.549) were estimated for LS and NA, respectively. The permanent environmental effect was insignificant for both traits, probably because only 24% of the records originated from dams with more than one parity, since 54% of the dams in the dataset had only a single parity record. Convergence could not be obtained when including both traits in a bivariate genetic evaluation. A Pearson correlation coefficient of 99.94 (P <0.0001) between the breeding values of the two traits from separate univariate analyses illustrates the strength of the relationship between the two traits. In Figure 5 the genetic trend for NA of the Boxer breed is indicated.
Figure 5 shows a slightly downward trend (b = -0.013; R2 = 0.19) up to 2000, followed by an increase in the number of puppies alive since then (b = 0.021; R2 = 0.24). This puts the breed at a higher genetic level for this trait in the 2000s.
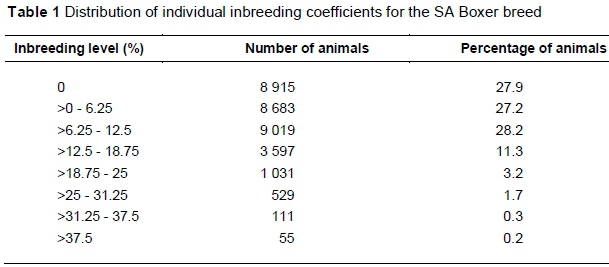
Table 1 presents the distribution of individual inbreeding coefficients for the SA Boxer breed. A total of 27.9% of dogs (8 915) were not inbred (0% inbreeding coefficients). Of these, only 5% had parent information and therefore were not inbred because of unrelatedness. The base consisted of 23% of the animals in the pedigree, with the assumption that they were not inbred. The largest proportion of the population (28.2%) had an inbreeding coefficient of 6.25% - 12.5% (Table 1), with 45% of the population having an inbreeding coefficient above 6.25%. The highest inbreeding coefficient observed in this study was 42.7% for a litter of two male and two female puppies born in 1992.
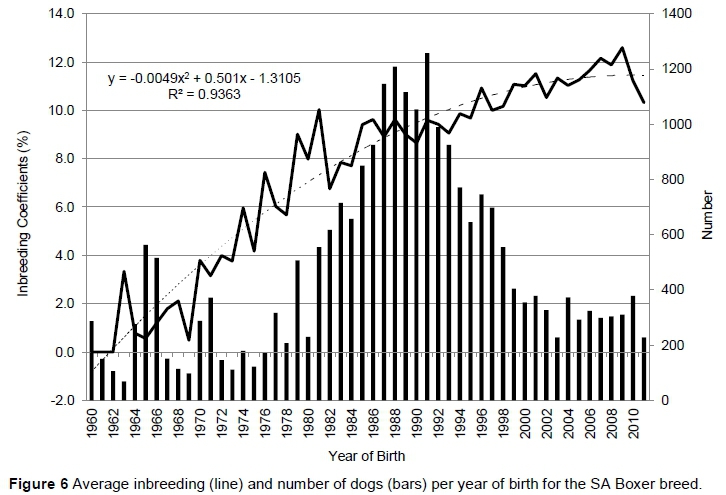
Figure 6 indicates the rate of inbreeding per year of birth for the SA Boxer breed. Inbreeding has increased steadily since 1960, with the average inbreeding coefficient for 228 dogs born in 2012 as 10.33%. The rate of increase was 0.42% (R2 = 0.84) per year from 1960 to 1982, and 0.14% (R2 = 0.77) per year afterwards. It was found that for every 1% increase in inbreeding, litter size of the SA Boxer breed decreased by 0.85 puppies and the number or puppies alive decreased by 1.4 puppies.
Discussion
The Boxer in South Africa is one of the few dog breeds that has kept pedigrees and recorded information on traits related to fitness. Fitness traits in this study included litter size and number of puppies alive at two weeks of age with heritability estimates of 0.23 ± 0.08 and 0.25 ± 0.08, respectively. These estimates compare favourably with those reported for Golden Retrievers for number of puppies born and number of puppies alive at 14 days (0.24 ± 0.07 and 0.28 ± 0.06) and German Shepherds (0.19 ± 0.05 and 0.21 ± 0.05) for the same traits (Hare & Leighton, 2006). Heritability estimates for similar traits in farm animal species, such as number of piglets born alive, tend to be lower with a mean heritability of 0.08 (Akanno, 2013).
Litters larger than five had higher mortalities at birth and/or low survivability. The effect of parity was more severe on early deaths of puppies compared with the effect of litter size, where the number of puppies born dead or that died within two weeks of birth increased from 0.43/litter at first parity to 0.82/litter at the fifth parity. A marked increase of 2.25 early deaths/litter was observed at the seventh parity. The interaction between litter size and parity should be considered in selecting and breeding dams. Litter size should be treated as an optimum trait, because maximum litter size would not necessarily result in a higher number of puppies at weaning, while the focus should be on improving the number of puppies born alive and number of puppies weaned.
The detrimental effects of inbreeding on fitness traits are well documented (Urfer, 2009; Johnson et al., 2011; Shariflou et al., 2011). In this study, litter size of the SA Boxer breed decreased by 0.85 puppies and number of puppies alive with 1.4 puppies for every 1% increase in inbreeding. A similar trend was reported by Gresky et al. (2005), in which a reduction in litter size and increase in percentage puppies born dead were found with increased inbreeding levels in Dachshunds. Based on the pedigrees analysed, 45% of the dogs had inbreeding levels above the recommended 6.25% (Van der Westhuizen & Mostert, 1998). In a French study (Leroy et al., 2006), nine breeds were studied with average coefficients of inbreeding exceeding 3.125%. Glazewska (2008) estimated that 69.3% of Polish hound litters had inbreeding coefficients larger than 25%, with 20% of litters exceeding inbreeding coefficients of 40%. In Germany the mean inbreeding coefficients of three scenthound breeds varied between 4.51% and 9.47%, with mean increases of 0.69%, 0.98% and 4.08%, respectively (Voges & Distl, 2009).
The rate of inbreeding for the SA Boxer breed up to 1982 was 0.42%/year, and thus was higher than the recommended rate of 0.1% - 0.2% per year to ensure adequate genetic diversity in a breed (Weigel, 2001). Since 1982, the rate of inbreeding has decreased to more acceptable levels of 0.14%/year, but this is still higher than the level of 0.043%/year reported for the Boxer breed in Australia (Shariflou et al., 2011). The levels in this study, however, correspond with a French study (Leroy et al., 2006), which reported rates of inbreeding of 0.1% - 0.5%/year in nine French dog breeds.
A study based on Australian dogs registered with the Australian National Kennel Council reported effective population sizes (Ne) for 32 dog breeds, in which the Boxer in Australia was classified as a breed with a large registry and a Ne of 113 with a decrease in inbreeding from 2000 to 2009 (Shariflou et al., 2011). In studies of Hanoverian hounds, inbreeding decreased rapidly when breeders engaged in pedigree recording and encouraged mating based on low relatedness (Voges & Distl, 2009).
This is the first study to evaluate fitness traits and inbreeding in the SA Boxer breed. Although the data structure was challenging, with small contemporary groups and large standard errors involved in the analyses, valuable information and trends could be indicated for the breed. It can therefore be recommended that inbreeding levels should be reduced, as they would influence the genetic diversity of the breed adversely in the long term. Heritability for fitness traits was found to be moderate and breeders can use this in selecting animals as parents for the next generation. The willingness and participation of breeders in recording pedigrees and fitness traits are positive steps towards effective genetic management of the breed. Assessments of fitness traits, inbreeding levels, rate of inbreeding and effect of inbreeding on fitness traits should be done regularly to assist in the survival and vitality of the SA boxer breed.
Conclusion
This study is the first genetic evaluation of the SA Boxer breed. The inbreeding results reported in this study follow international trends. Despite relatively high inbreeding levels, the rate of inbreeding has shown a decrease since 1982. There is room for improvement, with the focus on pedigree completeness, recording fitness traits and avoiding mating closely related animals.
References
Aguilar, I. & Misztal, I., 2008. INBUPGF90 V1.14. University of Georgia, USA. [ Links ]
Akanno, E.C., 2013. Meta-analysis of genetic parameter estimates for reproduction, growth and carcass traits of pigs in the tropics. Livest. Sci. 152, 101-113. [ Links ]
Asher, L., Diesel, G., Summers, J.F., McGreevy, P.D. & Collins, L.M., 2009. Inherited defects in pedigree dogs. Part 1: Disorders related to breed standards. Vet. J. 182, 402-411. [ Links ]
Cecchi, F., Paci, G., Spaterna, A. & Ciampolini, R., 2013. Genetic variability in Bracco Italiano dog breed assessed by pedigree data. Ital. J. Anim. Sci. 12, 348-352. [ Links ]
Glazewska, I., 2008. Genetic diversity in Polish hounds estimated by pedigree analysis. Livest. Sci. 113, 296-301. [ Links ]
Grandjean, D. & Vaisarre, J.J., 2004. The Royal Canine Dog Encyclopaedia. Guy Roland, Aniwa 10 Rue du Coliseé, Paris. [ Links ]
Gresky, C., Hamann, H. & Distl, O., 2005. Influence of inbreeding on litter size and the proportion of stillborn puppies in dachshunds. Berl. Munch. Tierarztl. 118, 134-139. [ Links ]
Groeneveld, E., Kovak, M. & Wang, T., 2001. PEST Version 4.2.3. University of Illinois, USA. [ Links ]
Groeneveld, E., Kovak, M. & Mielenz, N., 2010. VCE Users Guide and Reference Manual - Version 6.0. Institute of Farm Animal Genetics, FAL, Germany. [ Links ]
Hare, E. & Leighton, E.A., 2006. Estimation of heritability of litter size in Labrador Retrievers and German Shepherd dogs. J. Vet. Behav. 1, 62-66. [ Links ]
Johnson, H.E., Mills, L.S., Wehausen, J.D., Stephenson, T.R. & Luikart, G., 2011. Translating effects of inbreeding depression on component vital rates to overall population growth in endangered Bighorn sheep. Conserv. Biol. 25, 1240-1249. [ Links ]
Krontveit, R.I., Nødtvedt, A., Sævik, B.K., Ropstad, E., Skogmo, H.K. & Trangerud, C., 2010. A prospective study on Canine hip dysplasia and growth in cohort of four large breeds in Norway. Prev. Vet. Med. 97, 252-263. [ Links ]
Leroy, G., Rognon, X., Varlet, A., Joffrin, C. & Verrier, E., 2006. Genetic variability in French dog breeds assessed by pedigree data. J. Anim. Breed. Genet. 123, 1-9. [ Links ]
Lewis, T.W., Blott, S.C. & Woolliams, J.A., 2010. Genetic evaluation of hip score in UK Labrador. PlosOne, 5, e12797. [ Links ]
Malm, S., Fikse, W.F., Danell, B. & Strandberg, E., 2008. Genetic variation and genetic trends in hip and elbow dysplasia in Swedish Rottweiler and Bernese Mountain dog. J. Anim. Breed. Genet. 125, 403-412. [ Links ]
Meyer, F., Schawalder, P., Gaillard, C. & Dolf, G., 2012. Estimation of genetic parameters for behaviour based on results of German Shepherd dogs in Switzerland. Appl. Anim. Behav. Sci. 140, 53-61. [ Links ]
SAS, 2012. Statistical Analysis Systems, Enterprise Guide, Version 4.3. SAS Institute Inc., Cary, N.C., USA. [ Links ]
Shariflou, M.R., James, J.W., Nicholas, F.W. & Wade, C.M., 2011. A genealogical survey of Australian registered dog breeds. Vet. J. 189, 203-210. [ Links ]
Stock, K.F., Dammann, M. & Distl, O., 2012. Selection for conformation and conformational homogeneity of litters in the German shepherd dog. J. Anim. Sci. 90, 1088-1096. [ Links ]
Urfer, S.R., 2009. Inbreeding and fertility in Irish Wolfhounds in Sweden: 1976 to 2007. Acta. Vet. Scand. 51, 21. doi:10.1186/1751-0147-51-21 [ Links ]
Van der Westhuizen, J. & Mostert, B.E., 1998. Inbreeding and the Stud Breeder. Charolais J. 23-24. [ Links ]
Weigel, K.A., 2001. Controlling inbreeding in modern breeding programs. J. Dairy Sci. 84, 177-184. [ Links ]
Voges, S. & Distl, O., 2009. Inbreeding trends and pedigree analysis of Bavarian mountain hounds, Hanoverian hounds and Tyrolean hounds. J. Anim. Breed. Genet. 126, 357-365. [ Links ]
Zeiler, G.E., Van der Zwan, H. & Oosthuizen, M., 2013. Genetic testing of canine degenerative myelopathy in the South African Boxer dog population. J. S. Afr. Vet. Assoc. 84 (1), Art#1005. Available at: <http://www.jsava.co.za/index.php/jsava/article/view/1005/1243> [ Links ]
Received 12 August 2014
Accepted 2 November 2015
First published online 1 December 2015
# Corresponding author: bernice@studbook.co.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}