










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.45 no.2 Pretoria 2015
http://dx.doi.org/10.4314/SAJAS.V45I2.11
First-time characterization of JY-1-like sequence in goats
R. SharmaI, #; S. AhlawatI; A. MaitraI; M. RoyII; S. MandakmaleIII; M.S. TantiaI
INational Bureau of Animal Genetic Resources, Karnal-132 001, India
IIWest Bengal University of Animal & Fishery Sciences, Kolkata-700037, India
IIIMahatma PhuleKrishiVidyapeeth, Rahuri-413722, Ahmednagar (MS), India
ABSTRACT
The JY-1 protein is oocyte specific, and is associated with folliculogenesis and early embryo development, and thus influences the chance of pregnancy. It was the first protein of maternal origin to be described for a single ovulating species, namely cattle. JY-1-like sequences corresponding to 3' coding and the untranslated region have been reported in other vertebrate species. This is the first description of the partial JY-1 (exon 3 and 3'UTR) in a livestock species other than cattle and buffalo. The sequence was characterized in a panel of nine Indian goat breeds, which differ in reproductive traits (twinning percentage and age of sexual maturity). Forty three variations were recorded in the analysed region of goats JY-1 compared with cattle. Nucleotide variations in the codifying region of goats correspond to seven amino acid changes that could affect the biological function of the protein and possibly reproductive differences between goats (higher proportion of multiparous animals) and cattle (mainly uniparous). A novel single nucleotide polymorphism (C15329T) has been identified in Indian goats, which was genotyped in 272 animals from six breeds. Further studies to investigate other regions of the gene and its expression in goat female reproductive tissues would clarify the role of JY-1 in farm animal species that are not primarily uniparous.
Keywords: Capra hircus, embryogenesis, fertility, folliculogenesis, oocyte, PCR-RFLP, polymorphism, SNP
Introduction
The oocyte plays key roles in regulating female fertility through genes that are important for normal folliculogenesis, oocyte function and early embryogenesis. However, information about the oocyte-expressed genes required for fertility is limited, especially in farm animal species. For the first time an oocyte-specific protein, namely JY-1, which regulates the function of ovarian granulosa cells and early embryogenesis, was described in cattle by Bettegowda et al. (2007). The bovine JY-1 gene is approximately 16 kb in length, has three exons (25, 92, and 1,400 bp) and is located on chromosome 29. The codifying region comprises parts of exons 2 and 3. JY-1-like sequences, corresponding to exon 3 of the gene, were also identified in the genome of additional vertebrate species (human: chr 11; chimpanzee: chr 11; dog: chr 21; mouse: chr 7; and rat: chr 1) (Bettegowda et al., 2007). However, the syntenic loci do not encode for the complete JY-1 gene and lack sequences corresponding to exons 1 and 2, and thus a significant portion of the protein coding region. Therefore, Bettegowda et al. (2007) concluded that the oocyte-specific JY-1 protein is probably species specific. Incidentally, Rajput et al. (2013) recently identified expression of JY-1 in buffaloes and reported polymorphisms, including insertion of a thymine in the codifying region of exon 3 of the buffalo sequence. This insertion causes a frameshift mutation, modifying the 19 terminal amino acids of the buffalo protein and creating a premature stop codon (de Camargo et al., 2013). This finding may explain reproductive differences between cattle and buffaloes in terms of follicle recruitment, embryo development and incidence of twin pregnancies. Therefore, the presence of a functional ortholog that performs similar roles to JY-1 in other farm animal species cannot be ruled out. Thus, studies in farm animal species other than cattle and buffalo (e.g. sheep and goats) are necessary to further determine the specificity in structure and function of the JY-1 gene, as well as its dynamic in evolution.
In developing countries such as India, goats (Capra hircus) are regarded as multi-purpose domestic livestock species owing to their contribution to the economy. Reproductive traits are economically important for the goat production system; therefore, selection for these traits could improve productivity. In view of its biological role in other species, the JY-1 gene could be a candidate gene for reproductive traits in goats.
More than 20 breeds of goat have been reported in India with wide phenotypic variations in body size, coat colour, adaptations to various agro-climatic conditions, prolificacy and sexual maturity (Acharya, 1982). Breeds such as the Black Bengal display significant characteristics of early reproductive maturity and high prolificacy (number per kidding). On the other hand, breeds such as the Sirohi are late maturing with lower prolificacy. Since there are variations in reproductive performance in Indian goats, the objectives of the present study were, first, to obtain the status of the partial JY-1 gene (exon 3 and 3'UTR) of goats by generating a nucleotide sequence and sequence assembly in a panel of goat breeds that differ in reproductive traits, and, second, to detect possible intra- and interspecies (cattle and buffalo) polymorphisms to identify variability at these loci.
Material and Methods
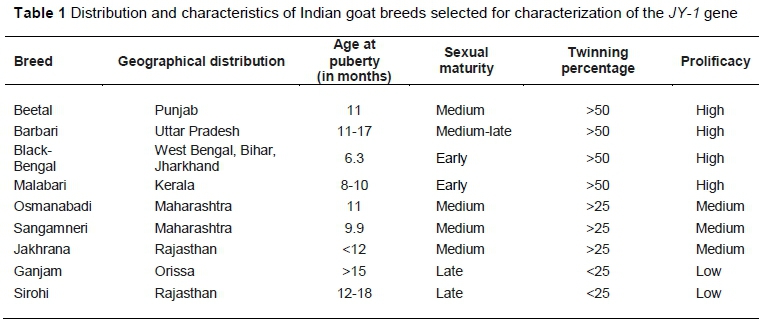
Nine well-recognized breeds of Indian goats were selected that differ in prolificacy (number per kidding) and age of sexual maturity, from different geographic and agro-climatic parts across India (Table 1). Unrelated animals of each breed were selected from their breeding tracts. Blood was collected aseptically from the jugular vein in a vacutainer tube (BD Biosciences, Germany) containing EDTA as anticoagulant. All samples were delivered back to the laboratory in iceboxes. The genomic DNA was extracted from white blood cells using a standard phenol-chloroform extraction protocol (Sambrook & Fristch, 1989).
To explore the distribution of genetic variations in the JY-1 gene, the panel included 45 animals (five animals per breed) (Table 1). A total of 272 animals from six breeds were utilized to estimate the polymorphism distribution. Black Begal, Barbari, Beetal, Osmanabadi and Sangamneri each contributed 48 animals, while 32 animals were of the Jakhrana breed.
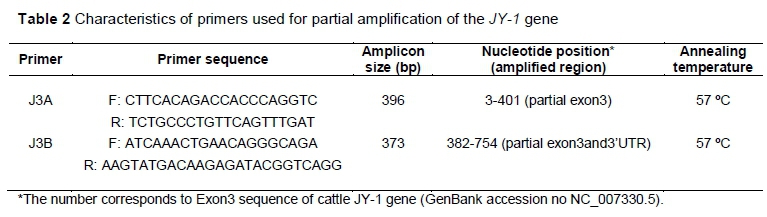
Two pairs of primers were utilized to amplify the exon 3 and 3'UTR of the JY-1 gene (Table 2), as reported in cattle (Camargo et al., 2012). The primers were synthesized by IDT (Integrated DNA Technologies, Inc.). Polymerase chain reaction (PCR) was carried out in 25 μΙ_ reaction volume with about 50 - 100 ng genomic DNA using i-cycler (BioRAD, USA). The reaction mixture consisted of 250 μΜ each of dATP, dCTP, dGTP, dTTP, 2.0 mM MgCl2, 50 pmol primer, 1U Taq polymerase (Sigma-Aldrich®) and corresponding Taq buffer. Amplification conditions consisted of initial denaturation for 3 min at 95 °C; followed by 35 cycles of denaturation at 94 °C for 30 sec; annealing at 57 °C for 30 sec; extension at 72 °C for 1 min; and finally extension at 72 °C for 10 min. The PCR products were separated by electrophoresis on 1.8% agarose gel in parallel with a 50 bp DNA ladder (Fermentas International, Inc.), enzymatically purified and sequenced using both primers (forward and reverse) by dideoxynucleotide chain termination reaction (Sanger et al., 1977). Sequencing was performed in an automated ABI -3100 sequencer (Applied Biosystems) using the ABI PRISM BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems).
Sequence data were edited manually using Chromas Ver.2.33, (http://www.technelysium.com.au/chromas.html).
Multiple sequence alignments were performed with the MegAlign program of Lasergene software version 5.07 (DNASTAR Inc., Madison, Wis.) to identify polymorphisms (mutations or single nucleotide polymorphisms (SNPs)). The coding DNA sequence was conceptually translated to amino acid sequences with ChromasPro software. The nucleotide BLAST program at NCBI (http://www.ncbi.nlm.nih.gov/BLAST) was used for sequence homology searches in public databases.
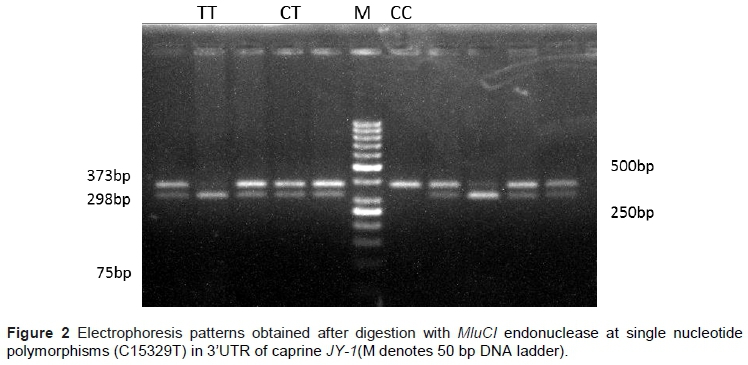
Once polymorphisms of the JY-1 gene had been identified in Indian goats, PCR-RFLP (polymerase chain reaction-restriction fragment length polymorphism) was used to genotype SNP in the 272 animals of six goat breeds, according to the manufacturer's instructions (Fermentas International, Inc.) by mixing 5 μL of the PCR product with 1X enzyme buffer and 5 units of MluCI (AATT) in a final volume of 15 μL. Digested fragments were separated by electrophoresis on 3% agarose gel stained with ethidium bromide. The fragments were photographed under a gel documentation unit and their sizes were estimated using a 50 bp DNA ladder (Fermentas International, Inc.) and the genotypes were recorded.
Results and Discussion
The two primer pairs amplified specific regions of the JY-1 gene according to expectations, with fragment sizes of 396 bp (primer pair J3A) and 373 bp (primer pair J3B). The fragments amplified from 45 goats of nine breeds were used to identify polymorphisms within and between species. For the first time, a 752 bp sequence was generated for the JY-1 gene in goats, which corresponds to the exon3 and 3'UTR of cattle JY-1 gene for which GenBank accessions were received (GenBank accession nos KJ425413.1 and KJ425414.1). BLAST analysis was carried out to find the percentage homology of this goat JY-1 sequence with cattle and buffalo. The DNA sequence of the Indian goat JY-1 region showed greater similarity (95%) with cattle (NC_007330.5) than with buffalo (93%) (JX070137.1). Since the caprine gene sequence is not yet available, the bovine JY-1 gene sequence (GenBank accession no. NC_007330.5) was used to compare the sequence information obtained by sequencing the PCR fragments (J3A and J3B) corresponding to expressive regions, as well as non-expressive ones of the JY-1 gene in Indian goat.
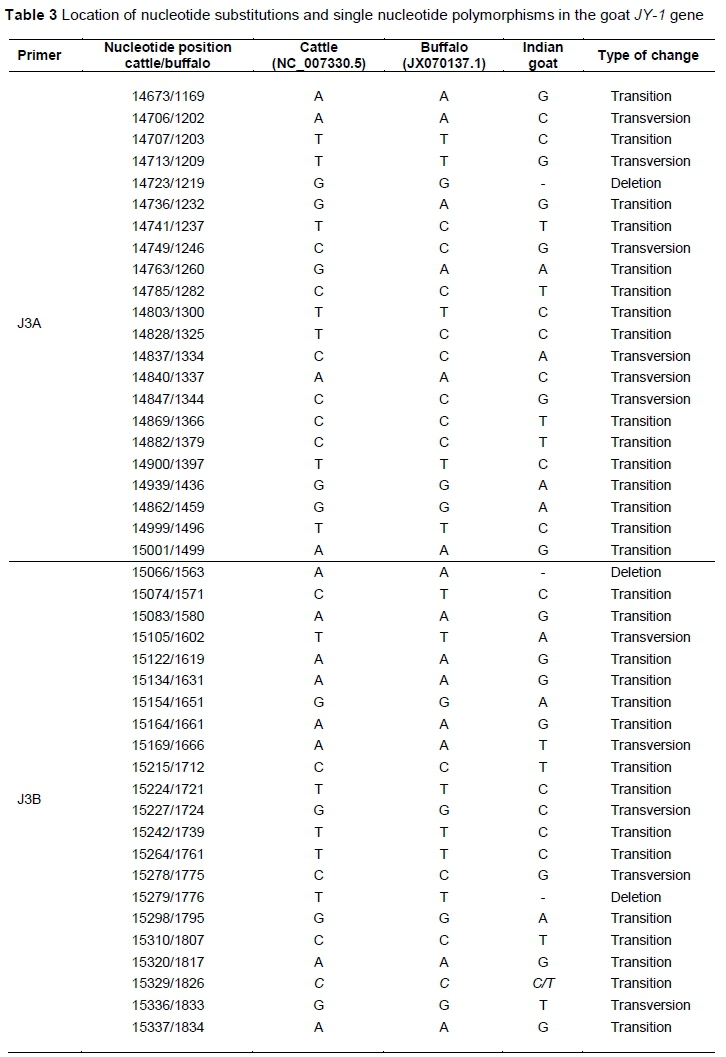
The authors observed 43 variations, including three deletions (Table 3) in Indian goat JY-1 compared with cattle, while most nucleotide substitutions were transition changes (69.8%). Sixteen nucleotide variations were observed in codifying regions, of which seven caused amino acid changes in the corresponding protein. Changes in the amino acid for the conceptualized 62 amino acid polypeptide chain were: cysteine > glycine (28th amino acid), arginine > leucine (31st amino acid), proline > leucine (32nd amino acid), valine > phenylalanine (33rd amino acid), leucine>valine (40th amino acid), leucine > phenylalanine (52nd amino acid) and phenylalanine > leucine (58th amino acid).
The amino acid residues, glycine and proline that influence chain orientation are worth noting (28th and 32nd amino acid) because the differences in the structure of a protein can reflect in altered function. Thus, these amino acid changes may reflect indifference in the biological functions of JY-1 protein between cattle and goats. Substitution of amino acids has been reported in buffalo (Bubalus bubalis), where the insertion of a thymine was detected in the codifying region of exon 3 compared with cattle (de Camargo et al., 2013). It resulted in frameshift from this point onwards, modifying the 19 terminal amino acids of the buffalo protein and creating a premature stop codon. Reproductive differences between cattle and buffaloes in terms of follicle recruitment, embryo development and incidence of twin pregnancies may be explained by the difference in the JY-1 protein. Similarly, the differences in amino acid composition among cattle, sheep and goat may explain reproductive differences in these species. JY-1 acts in early embryonic development and in the granulosa cells during luteinization (Bettegowda et al., 2007). The role of JY-1 in preovulatory events and early embryonic development (Bettegowda et al., 2007) may explain the difference in the luteinization process and pace of embryonic development owing to the different protein configurations between species. For example, embryo development is faster (12 - 24 h) in buffaloes because of the early entry of embryos into the uterus (4 - 5 days after oestrus) (Campanile et al., 2010). Another difference described by Gimenes et al. (2011) is that during folliculogenesis in buffaloes there is no decrease in follicle-stimulating hormone levels or increase in luteinizing hormone levels in blood plasma at the time of follicle recruitment. In addition, because protein JY-1 acts during the early stages of embryogenesis (when monozygotic embryos are formed) and in preovulatory events (when more than one oocyte may be recruited and ovulated at the same time, the origin of dizygotic twins), the protein difference may explain the difference in the percentage of multiple births among species. Cattle and buffaloes are uniparous (single-ovulating) livestock species. However, <5% twin births have been reported in beef cattle (Bos taurus) (Kirkpatrick, 2002). In buffaloes (Bubalus bubalis) twin births are extremely rare (Singh et al., 2009; Shukla et al., 2011). Goat and sheep are examples of livestock species that have primiparous and multiparous animals. Some Indian goat breeds, such as Ganjam and Sirohiare, are mainly primiparous, whereas others, such as Black Bengal, Barbari and Malabari, have more than 50% twinning (Acharya, 1982).
A mixed sample is ideal for SNP discovery as diverse samples can increase the yield of SNPs, thus nine well-established goat breeds of India were explored to prospect SNPs in the JY-1 gene. An SNP was identified (C/T) in the region amplified with primer pair J3B at position 15329 bp (numbering as per cattle GenBank accession no. NC_007330.5). This is the first polymorphism described in the caprine JY-1 gene.
All three possible genotypes were recognized for the SNP (C15329T) in Indian goats (Figure 1). Mutant allele (T) was predominant in goats with the allele frequency of 0.71 in the panel consisting of 45 animals belonging to nine goat breeds. Distribution of SNP C15329T in Indian goats was explored by designing PCR-RFLP and increasing the number of animals for each breed. This PCR-RFLP method is a convenient tool for establishing the genotype-phenotype association, as the method is cheap, efficient and reproducible, and thus can be used by laboratories with limited facilities. The genotypes that were identified were CC (373 bp), CT (373, 298 and 75 bp), and TT (298 and 75 bp) (Figure 2).
The allele and genotype frequencies and compliance with Hardy-Weinberg equilibrium (5%) are shown in Table 4. All breeds were in Hardy-Weinberg equilibrium at this locus.
Genotype distribution of the mutation did not show obvious differences between sexually precocious breeds and sexually late-maturing breeds, or any consistency with high or low prolificacy goat breeds. Increasing the sample size from 45 (panel utilized for prospecting the SNPs) to 272 resulted in change of allele frequencies from 0.71 (T) to 0.61 (T). Thus, further replication with a larger sample size is required to establish the allelic frequency at this locus.
The novel SNP (C15329T) identified among the Indian goats is located in the 3'UTR. The significance of the 3' untranslated region for JY-1 is not known, but based on accumulating evidence for important regulatory roles (polyadenylation, translation efficiency, localization and stability of the mRNA) of noncoding RNAs (Prasanth & Spector, 2007), a functional role for the observed mutation cannot be discounted. The regions amplified in the present study have been shown to have seven SNPs in cattle (de Camargo et al., 2012). The haplotypes of four of these SNPs were found to be correlated with sexual precocity in Nellore heifers at 8%. Eighteen polymorphism in codifying regions of cattle JY-1, including the seven reported earlier, were detected one year later (de Camargo et al., 2013) by sequencing in 20 unrelated cattle (Bos taurus indicus). Polymorphism in the codifying region has also been reported in buffaloes (Bubalus bubalis) where some of the polymorphisms in codifying regions caused amino acid changes (de Camargo et al., 2013). Seven SNPs were significant for reproductive traits in Nellore cattle and the most significant was T12999A, being associated with the occurrence of early pregnancy (de Camargo et al., 2014). The C15329T SNP reported here may serve as a marker for potential association with caprine sexual fecundity and precocity traits. Since no phenotypes on fecundity and sexual precocity are currently available for goats, it may not be possible to link novel SNP noticed in the present study with these traits. Information regarding caprine JY-1 should be useful for researchers that are willing to work with genetic markers in goats or genotyping in case-control association studies.
Conclusion
In terms of fertility traits, the markers identified so far are located in genes or are related to genes that act as transcription factors in the nervous system during body growth and in the lipid metabolism of animals. Recent results involving the JY-1 gene in cattle have shown that genes involved in the female reproductive tract contribute to the genetic variability of reproductive traits. This is the first report of partial JY-1 gene in caprines, where nucleotide variations in coding and 3'UTR, as well as a novel SNP, were identified. Considering the importance of the reproductive traits in small ruminants, investigating other regions of the JY-1 gene or expression in tissues of the goat female reproductive system should be the target of future studies.
Acknowledgements
The authors wish to thank the Indian Council of Agricultural Research, New Delhi, for supporting the study under Network Project on Animal Genetic Resources of India.
References
Acharya, R.M., 1982. Sheep and goat breeds of India. FAO Animal Production and Health Paper 30, FAO United Nations, Rome, Italy. [ Links ]
Bettegowda, A., Yao, J., Sen, A., Li, Q., Lee, K., Kobayashi, Y., Patel, O.V., Paul, M., Coussens, P.M., Ireland, J.J. & Smith, G.W., 2007. JY-1 an oocyte-specific gene regulates granulosa cell function and early embryonic development in cattle. Proc. Natl. Acad. Sci., USA.104, 17602-17607. [ Links ]
Campanile, G., Baruselli, P.S., Neglia, G., Vecchio, D., Gasparrini, B., Gimenes, L.U., Zicarelli, L. & D'Occhio, M.J., 2010. Ovarian function in the buffalo and implications for embryo development and assisted reproduction. Anim. Reprod. Sci. 121, 1-11. [ Links ]
de Camargo, G.M.F., Cardoso, D.F., Gil, F.M.M., Fonseca, P.D.S., Zetouni, L., Braz, C.U., Freitas, Fa'bio, A.C., Souza, R.P., Aspilcueta-Borquis, R.R., Baldi, F., Albuquerque, L.G., Regitano, L.C.A. & Tonhati, H., 2012. First polymorphisms in JY-1 gene in cattle (Bos taurus indicus) and their association with sexual precocity and growth traits. Mol Biol Rep. 39, 10105-10109. [ Links ]
de Camargo, G.M.F., Baldi, F., Regitano, L.C.A. & Tonhati, H., 2013. Characterization of the exonic regions of the JY-1 gene in zebu cattle and buffaloes. Reprod. Dom. Anim. 48, 918-922. [ Links ]
de Camargo, G.M.F., Costa, R.B., de Albuquerque, L.G., de Almeida Regitano, L.C., Baldi, F. & Tonhati, H., 2014. Association between JY-1 gene polymorphisms and reproductive traits in beef cattle. Gene 533, 477-480. [ Links ]
Gimenes, L.U., Carvalho, N.A.T., Sa Filho, M.F., Vannucci, F.S., Torres-Junior, J.R.S., Ayres, H., Ferreira, R.M., Trinca, L.A., Sartorelli, E.S., Barros, C.M., Beltran, M.P., Nogueira, G.P., Mapletof, R.J. & Baruselli, P.S., 2011. Ultrasonographic and endocrine aspects of follicle deviation and acquisition of ovulatory capacity in buffalo (Bubalus bubalis) heifers. Anim. Reprod. Sci. 123, 175-179. [ Links ]
Kirkpatrick, B.W., 2002. Management of twinning cow herds. J. Anim. Sci. 80, E14-E18. [ Links ]
Prasanth, K.V. & Spector, D.L., 2007. Eukaryotic regulatory RNAs, an answer to the 'genome complexity' conundrum. Genes. Dev. 21, 11-42. [ Links ]
Rajput, S.K., Kumar, P., Roy, B., Verma, A., Pandey, H.P., Singh, D., De, S. & Datta, T.K., 2013. Identification of some unknown transcripts from SSH cDNA library of buffalo follicular oocytes. Animal 4, 446-454. [ Links ]
Sambrook, J. & Fritsch, E.F., 1989. Molecular cloning- A laboratory manual. 2nd ed. Cold Spring Harbor, New York. pp. 9.16-9.23. [ Links ]
Sanger, F., Nicklen, S. & Coulson, A.R., 1977. DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74, 5463-5467. [ Links ]
Shukla, S.P., Mudasir, Q. & Nema, S.P., 2011. Dystocia due to a conjoined twin monster foetus in a female buffalo. Buffalo Bull. 30, 12-13. [ Links ]
Singh, A.K., Brar, P.S., Pandey, A.K., Gandotra, V.K. & Prabhakar, S., 2009. Amorphous globosus monster in a buffalo. Indian Vet. J. 86, 846-847. [ Links ]
Received 22 August 2014
Accepted 4 May 2015
First published online 11 July 2015
# Corresponding author: rekvik@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}