










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.45 n.2 Pretoria 2015
http://dx.doi.org/10.4314/sajas.v45i2.4
In vitro degradation of melamine by ruminal microorganisms
Department of Animal Sciences, Stellenbosch University, Private Bag X1, Matieland 7602, South Africa
ABSTRACT
An in vitro study was conducted to determine the extent of melamine degradation in rumen liquor. Rumen liquor was collected from two ruminally cannulated Holstein cows on four separate dates, one week apart. Erlenmeyer flasks (250 mL) were prepared for incubation by adding 1000 mg of a dairy feed substrate, 100 mg melamine and 100 mL incubation medium, purged with CO2 and fitted with rubber stoppers equipped with one-way gas release valves. The initial melamine concentration was thus 1000 mg/L. The substrates consisted of 600 mg of a commercial dairy concentrate, 200 mg lucerne hay and 200 mg oat hay. The incubation medium consisted of 19 mL rumen liquor, 77 mL of Van Soest buffer and 4 mL of a reducing solution. The flasks were incubated at 39 °C for 0, 6, 24 or 48 hours (two flasks per time in each of four replicates). The 0 h incubation served as a control treatment to enable the calculation of melamine recovery values. For the control treatment (0 h), fermentation was terminated at the onset of the trial by aerating the rumen liquor and submerging the flasks in 50 mm ice. On termination of the incubation, 100 mL 0.2 M perchloric acid was added to each flask in order to dissolve any undegraded melamine. Melamine concentrations were determined by liquid chromatography-tandem mass spectrometry. Melamine degradation was low after 6 hours and 24 hours of incubation (3.2% and 5.5%, respectively) and increased to 13.6% after 48 h of incubation. It was concluded that melamine has low degradability in rumen liquor.
Keywords: Non-protein nitrogen, rumen fermentation, rumen incubation
Introduction
The industrial chemical melamine (1,3,5-triazine-2,4,6-triamine) is used in manufacturing plastic ware, laminates and paints. Pure melamine has a high nitrogen content (667 g/kg; Merck, 2001), which theoretically equates to a crude protein content (N x 6.25) of 4167 g/kg. This high nitrogen content makes it an attractive adulterant for protein feedstuffs, as demonstrated during the 2008 worldwide pet food recalls and the 2009 tainted infant formula incidents in China.
Following reports by Loosli et al. (1949) that ruminants are able to utilize non-protein nitrogen (NPN) sources to produce essential amino acids, Colbey & Mesler (1958) patented the use of melamine as an NPN source for ruminants. By recognizing the high nitrogen content of melamine and its structural similarities to cyanuric acid, which, according to Clark et al. (1965), proved to be a safe NPN source for sheep, MacKenzie (1966) investigated the potential use of melamine as an NPN source for ruminants. He concluded that melamine was an inefficient NPN source owing to a reduction in apparent nitrogen digestibility, reduced feed intake and the inexplicable deaths of five sheep that were fed 10 g melamine per day. Newton & Utley (1978) found that melamine increased in vitro rumen ammonia concentration, but reported that the rate of melamine hydrolysis in the rumen was insufficient to promote maximum ruminal protein synthesis and concluded that melamine was not an acceptable NPN source for ruminants.
According to Cruywagen et al. (2009), approximately 2% of melamine ingested by dairy cows was excreted via the milk. In a study with sheep to quantify the partitioning of absorbed dietary melamine to various tissues and excretion via faeces and urine, Cruywagen et al. (2011) found that 53.2% of ingested melamine was excreted via urine and 23.3% via faeces. They reported that the melamine residue in muscle tissue accounted for approximately 3.6% of ingested melamine. Of the balance of 18.6%, which was not accounted for, it was found that a small amount was partitioned to other organs and blood, while the authors speculated that the rest could have been degraded in the rumen.
Diversity in the bacterial population is extensive and complex (Russell & Hespell, 1981) with a wide range of substrate affinities. Newton & Utley (1978) reported increased rumen ammonia (NH3) concentrations with the addition of melamine via an in vitro trial. In addition, the in vitro metabolism of melamine to melamine analogues (e.g. ammeline, ammelide, cyanuric acid) by aerobic microorganisms such as Arthrobacter spp., Klebsiella terragena and Pseudomonas spp. was reported by Strong et al. (2002), Shelton et al. (1997) and Jutzi et al. (1982), respectively. Although neither the Arthrobacter spp. nor Klebsiella spp. was isolated from the rumen, Duncan et al. (1999) reported the presence of Pseudomonas aeruginosa in the rumen of sheep. The only documented study that was found on the kinetics of ruminal melamine degradation was that of Sun et al. (2012), who reported that 44.5% of ingested melamine disappeared from the rumen within 12 hours. They explained the disappearance by postulating that it was absorbed via the rumen wall or degraded by the rumen microbes. Their study did not take feed and water intake into account or melamine losses via passage of digesta from the rumen, which could have had a significant effect on ruminal melamine concentration.
The current in vitro study was therefore planned to quantify melamine disappearance in a closed system to exclude the confounding effects of a dynamic in vivo rumen environment.
Materials and Methods
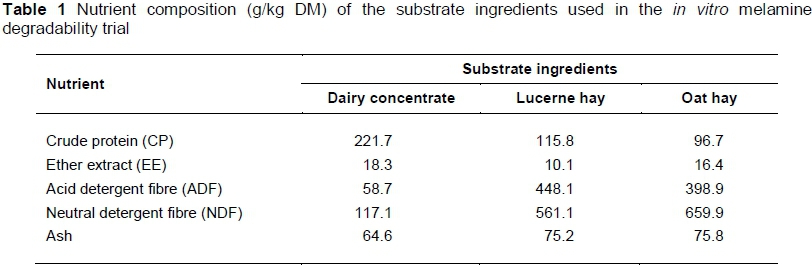
Rumen liquor was collected from two ruminally cannulated lactating Holstein cows in four consecutive runs, one week apart. Cows weighed 719 ± 9.2 (SE) kg and were kept at Welgevallen Experimental Farm of Stellenbosch University. They had free access to oat hay and received 25 kg/d of a commercial semi-complete lactating cow diet (nutrient composition in Table 1). The trial was approved by Stellenbosch University's Research Ethics Committee: Animal Care and Use (ref 10LVCRU02).
Rumen liquor was collected just before the morning feeding at 07:00 and transferred to pre-heated 1L thermos flasks. Approximately 200 g solid material from the rumen was added to each flask. Flasks were filled to the brim before the lid was replaced to prevent aeration. In the laboratory, the rumen liquor of each thermos flask was blended separately in a commercial blender for 20 seconds under a continuous stream of CO2 to maintain anaerobic conditions. For each cow, 500 mL of the blended rumen liquor was filtered through four layers of cheesecloth into a 1L preheated Erlenmeyer flask to obtain a pooled sample. The flask was purged with CO2, fitted with a rubber stopper, and placed in a water bath at 39 °C. The complete procedure was followed in four separate runs to yield four replicates. The mean initial pH of the rumen liquor was 6.35 ± 0.02 (SE).
A series of eight 250 mL Erlenmeyer flasks was prepared for each set of in vitro incubations. Each flask contained 100 mg melamine (melamine 99%, Sigma-Aldrich, St. Louis, Mo), 1000 mg substrate and 100 mL incubation medium. The initial melamine concentration was thus 1000 mg/L. This significant concentration was decided on to ensure that any degradation of melamine would be readily detected. The substrate consisted of 600 mg of a commercial dairy concentrate (provided by Meadow Feeds, Paarl, South Africa), 200 mg lucerne hay and 200 mg oat hay in order to simulate a typical dairy ration of 60 : 40 concentrate to roughage ratio. All substrate ingredients (dairy concentrate, lucerne hay and oat hay) were ground with a laboratory hammer mill (Scientec, South Africa) through a 1 mm screen. Chemical analyses of the substrate ingredients were performed in duplicate according to AOAC (2002) methods for dry matter (DM) (method 934.041), ash (method 942.05), crude protein (CP) (method 990.03) and ether extract (method 920.39). The ANKOM Fiber Analyzer (ANKOM® Technology Corp., Macedon, NY, USA) was used to determine ADF and NDF. Heat-stable alpha-amylase and sodium sulphite were used in the assay. The nutrient composition of the substrate is presented in Table 1.
The incubation medium consisted of 19 mL prepared rumen liquor, 77 mL buffer solution and 4 mL reducing solution. The buffer and reducing solutions were prepared according to Van Soest & Robertson (1985). The flasks were purged with CO2, fitted with rubber stoppers that had one-way gas release valves, and incubated at 39 °C for 0, 6, 24 or 48 hours. In each of the four replicate runs, two flasks were incubated per time to allow for duplicate analysis per replicate. The 0 h served as control treatment to calculate melamine recovery. In this treatment, fermentation was terminated before the onset of the trial by aerating and submerging the Erlenmeyer flasks in 50 mm ice water. Fermentation of the incubated samples was terminated in the same way after the predetermined incubation times. On termination of fermentation, 100 mL 0.2 M perchloric acid was added to each Erlenmeyer flask to dissolve undegraded melamine. Representative rumen liquor samples were taken and stored in airtight containers at -20 °C pending melamine analysis.
An adapted method from Shai et al. (2008) was used for melamine analyses. Thawed rumen liquor samples were centrifuged at 4 500 x g for 5 min. The conditioning of the cation-exchange solid-phase extraction cartridges (Phenomenex Strata SCX; 55 μm, 70 A, 500 mg/3 mL, supplied by Separations, Randburg, South Africa) was done with 6 mL methanol, followed by 6 mL distilled water. The supernatants of the rumen liquor samples (3 mL) were loaded onto the cartridges, together with 100 μL of a 0.5 mg/L stable isotope-labelled melamine (13C3H615N3) internal standard solution (Cambridge Isotope Laboratories Inc., Andover, Mass). Therefore, 0.05 μg of the labelled melamine was loaded onto each cartridge. The cartridges were then washed with 6 mL 0.1 N HCl, followed by 6 mL methanol, and allowed to aspirate under vacuum for 1 min. The melamine was eluted with 6 mL ammonium hydroxide : methanol : dichloromethane (1 : 5 : 5) and collected into clean glass tubes. The resulting extracts were then dried under a stream of nitrogen, re-suspended with 1 mL 50% acetonitrile and transferred to individual vials for analysis. Samples were analysed for melamine by liquid chromatography-tandem mass spectrometry (LC/MSMS) using a Waters API Quattro micro triple quadruple mass spectrometer, coupled to a Waters 2690 HPLC (Waters Corp., Milford, Mass). For this method, the limit of detection for rumen liquor samples was 0.001 mg/kg.
Melamine recovery values were determined from the control (0 h) treatment (with an initial melamine concentration of 1000 mg/L) and found to be 99.4%. The 0 h value was adjusted to 100% (1000 mg/mL) and the concentrations of the incubated samples were adjusted accordingly by multiplying the values by 1.006.
A repeated measures ANOVA with the compound symmetry assumption on the correlation structure over time was applied to analyse responses, using Statistica 64 (2013). The following model was used:
Yij = μ + αi + B(i)j + εij,
where μ is the overall mean, ai the effect of the ith time of measurement, βΜ the effect of the jth cow (or run) at time i and ε0, the random error. The Bonferroni post-hoc procedure was used to discern between significant least square means, and significance was declared at P <0.05.
Results and Discussion
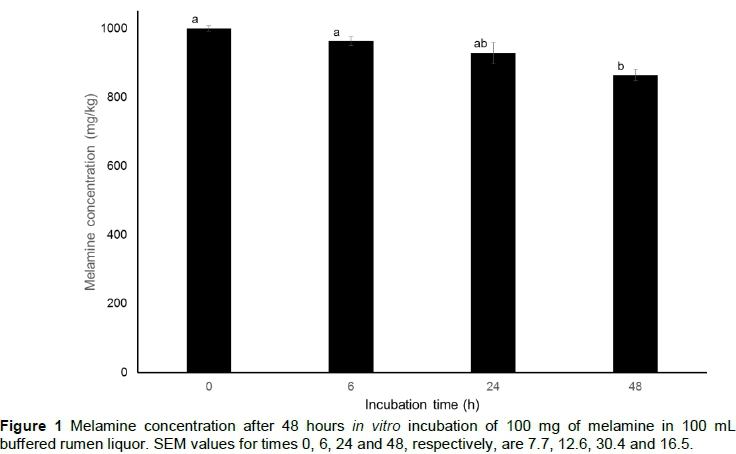
The change in ruminal melamine concentration over time is indicated in Figure 1. As incubation time increased, the melamine concentration decreased gradually. The melamine concentration at 48 h (864 mg/kg) was significantly (P = 0.004) lower compared with previous hours (1000, 963 and 928 mg/kg at 0, 6 and 24 h, respectively). Sun et al. (2012) investigated the kinetics of melamine disappearance from the rumen of dairy cows over 12 hours. They found that melamine concentration decreased exponentially, and that the value at 12 h post ingestion was approximately 50% of the 1 h value. They concluded that 44.5% of ingested melamine was absorbed via the rumen wall or degraded by rumen microbes after 12 hours. However, they did not take into account the passage of digesta from the rumen and feed and water intake, which may have had an effect on ruminal melamine concentration. To relate melamine concentration in a dynamic environment, such as the rumen, directly to degradation may result in an overestimation of degradation rates. In the current study, a closed in vitro system was used and the decrease in melamine concentration over time was probably because of microbial degradation alone.
The extent of melamine degradation (melamine disappearance expressed as % of initial melamine) in the current study is indicated in Figure 2. Initial degradation rate appeared to have been higher than later rates. By 48 h, the amount (%) of melamine that was degraded in rumen liquor (13.6%) was significantly (P = 0.004) higher than at 6 h (3.7%). However, to put these values in context under practical conditions, rumen retention time should be considered. When the accepted rumen passage rate of kp =.08 for lactating dairy cows is applied, the rumen retention time of a high concentrate feed would be 12.5 h (100/8), which would be the corresponding time point for the calculation of effective degradability. In the current study, samples were not incubated specifically for 12.5 h, but the effective degradability of melamine would be a value between 3.7% (6 h incubation) and 7.2% (24 h incubation). Results from the current study would therefore suggest that the effective degradability of melamine is very low at this dosage. The question may arise whether the significant amount of melamine added to the incubation medium (1 g/L) might have over-saturated the N supply and thus decreased degradability values. Taken into account that the melamine would have been mostly insoluble in the buffered medium (solubility is discussed below), that authors postulate that the availability of melamine N would have been low and probably not overwhelming.
Some degradation of melamine in rumen liquor was expected, as Jutzi et al. (1982) reported the hydrolytic cleavage of melamine into ammonia by Pseudomonas sp. strain A in vitro. The presence of Pseudomonas sp. strain A in rumen liquor is unclear. The production of ammonia following melamine cleavage (Jutzi et al., 1982) may also explain the increased ammonia concentration (although not significant) observed in the in vivo and in vitro trials of Newton & Utley (1978). Newton & Utley's (1978) in vitro study notes that higher ammonia concentrations were observed after 24 h of incubation when the melamine substrate was incubated in rumen liquor obtained from a steer-fed melamine compared with rumen liquor from a steer-fed cottonseed. This observation may infer that a period of adaptation may be required by rumen microbes in order to hydrolyse melamine more efficiently.
The estimated effective ruminal melamine degradability of between 3.7% and 7.2% may explain part of the 18% unaccounted for melamine observed in the trial of Cruywagen et al. (2011) who investigated excretion routes of melamine in sheep receiving a melamine-tainted concentrate supplement. McDonald (1981) reported that protein degradation is affected by retention time in the rumen and inversely related to passage rate. Furthermore, small feed particles pass through the rumen with the liquid phase via rumen contractions at a higher rate compared to larger particles (Poppi et al., 2000). This would also apply to melamine and, owing to the small particle size of melamine, it could be expected that the retention time in the rumen would be short and effective degradability very low.
Finally-, ruminal pH may affect melamine solubility and consequently its degradability. In the current study, pH levels stayed fairly constant over the incubation period. Initial pH of the rumen liquid was 6.35 ± 0.02 (SE), whereas the values in the incubation medium were 6.72 ± 0.04 (SE) at 6 h, 6.74 ± 0.05 (SE) at 24 h and 6.75 ± 0.05 (SE) at 48 h. According to Wiwanitkit & Wiwanitkit (2013) melamine is insoluble at a physiological pH (7.35 to 7.45) in the human body. Their determination of solubility was based on the canonical value (CV-CV), which is a variable that determines whether a molecule is soluble. When CV-CV > 0, the molecule would be soluble and when CV-CV < 0 it would be insoluble. These authors reported a series of CV-CV values for melamine that ranged between -0.22 and -0.26 when pH was increased from 6.4 to 8.4. In the current study, it can thus be accepted that melamine would have been mostly insoluble, which could explain the low degradability. Although information could not be found for the exact pH at which melamine becomes soluble, it is possible that when ruminal pH decreases (e.g. when feeding high concentrate diets), the chances of melamine becoming soluble increase. Even so, the relative short ruminal retention time of melamine that would be expected in dairy cows, together with the low solubility of melamine at pH > 6, would result in most of ingested melamine reaching the small intestine intact.
Conclusion
The results of the current trial showed that melamine has a low in vitro degradability in rumen liquor. Because the rumen liquor donor cows received melamine-free diets, the rumen microbes were not adapted to melamine in the substrate. It is speculated that degradability might have been somewhat higher, although still low, if the cows had been exposed to melamine-tainted feed over time. Melamine solubility in relation to pH levels and temperature of the liquid, as well as the availability of certain minerals such as potassium and carbonates, needs to be investigated. In addition, the ability of microbes that have adapted over time to deal with compounds such as melamine and the effect of melamine degradation on fermentation end products warrant further research.
References
AOAC International, 2002. Official Methods of Analysis. (17th ed). Association of Official Analytical Chemists, Arlington, Virginia, USA. [ Links ]
Clark, R., Barratt, E.L. & Kellerman, J.H., 1965. A comparison between nitrogen retention from urea, biuret, triuret and cyanuric acid by sheep on a low protein roughage diet. J. S. Afr. Vet. Med. Ass. 36, 79. [ Links ]
Colbey, R.W. & Mesler Jr., R.J., 1958. Ruminant feed compositions. U.S. Patent No.2819968. [ Links ]
Cruywagen, C.W., Stander, M.A., Adonis, M. & Calitz, T., 2009. Hot topic: Pathway confirmed for the transmission of melamine from feed to cow's milk. J. Dairy Sci. 92, 2046-2050. [ Links ]
Cruywagen, C.W., Van de Vyver, W.F.J. & Stander, M.A., 2011. Quantification of melamine absorption, distribution to tissues and excretion by sheep. J. Anim. Sci. 89, 2164-2169. [ Links ]
Duncan, S.H., Doherty, C.J., Govan, J.R.W., Neogrady, S., Galfi, P. & Stewart, C.S., 1999. Characteristics of sheep-rumen isolates of Pseudomonas aeruginosa inhibitory to the growth of Escherichia coli O157. FEMS Microbiol. Lett. 180, 305-310. [ Links ]
Jutzi, K., Cook, M. & Hutter, R., 1982. The degradative pathway of the s-triazine melamine. Biochem. J. 208: 679-684. [ Links ]
Loosli, J.K., Williams, H.H., Thomas, W.E., Ferris, F.H. & Maynard, L.A., 1949. Synthesis of amino acids in the rumen. Science 110:144 [ Links ]
MacKenzie, H.I., 1966. Melamine for sheep. J. S. Afr. Vet. Med. Ass. 37, 153-157. [ Links ]
McDonald, I., 1981. Short note: A revised model for the estimation of protein degradability in the rumen. J. Agric. Sci., Camb. 96, 251-252. [ Links ]
Merck, 2001. The Merck Index. 13th ed. Merck Research Laboratories, Whitehouse Station, NJ. pp. 5831-5834 [ Links ]
Newton, G.L. & Utley, P.R., 1978. Melamine as a dietary nitrogen source for ruminants. J. Anim. Sci. 47, 1338-1344. [ Links ]
Poppi, D.P., France, J. & McLennan, S.R., 2000. Intake, passage and digestibility. In: Feeding Systems and Feed Evaluation Models. Eds: Theodorou, M.K. & France, J., CAB International Publishing, Oxon, U.K. pp. 35-52. [ Links ]
Russell, J.B. & Hespell, R.B., 1981. Microbial rumen fermentation. J. Dairy Sci. 64, 1153-1169. [ Links ]
Shelton, D.R., Karns, J.S., McCarty, G.W. & Durham, D.R., 1997. Metabolism of melamine by Klebsiella terragena. Appl. Environ. Microbiol. 63, 2832-2835. [ Links ]
Strong, L.C., Rosendahl, C., Johnson, G., Sadowsky, M.J. & Wackett, L.P., 2002. Arthrobacter aurescens TC1 metabolizes diverse s-triazine ring compounds. Appl. Environ. Microbiol. 68, 5973-5980. [ Links ]
Shai, J., Mallet, C., Young, M., Li, J., Meng, Y. & Qi, C., 2008. Application note: Rapid specific analysis of melamine contamination in infant formula and liquid milk by UPLC/MS/MS. Waters Corporation, Milford, MA. [ Links ]
Statistica 64, 2013. StatSoft Inc. Tulsa, OK. USA. [ Links ]
Sun, P., Wang, J.Q., Shen, J.S. & Wei, H.Y., 2012. Pathway for the elimination of melamine in lactating dairy cows. J. Dairy Sci. 95, 266-271. [ Links ]
Van Soest, P.J. & Robertson, J.B., 1985. Analysis of Forages and Fibrous Foods. Cornell University, Ithaca, NY pp. 80-165. [ Links ]
Wiwanitkit, S. & Wiwanitkit, V., 2013. Solubility of Melamine Crystal in Different pH Conditions: Clinical Implication for Management of Melamine Intoxication. Letter to the Editor. Saudi J. Kidney Dis. Transpl. 24, 1019-1021. [ Links ]
Received 11 November 2014
Accepted 18 April 2015
First published online 17 May 2015
# Corresponding author: cwc@sun.ac.za
1 Present address: Tanqua Feeds, Riviersonderend


{kind=link}
{kind=link}
{kind=link}