










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.44 no.4 Pretoria 2014
http://dx.doi.org/10.4314/sajas.v44i4.3
Crossbreeding to increase beef production: additive and non-additive effects on fitness traits
A. TheunissenI, II, #; M.M. ScholtzI, III; F.W.C. NeserI; M.D. MacNeilI, III, IV
IDepartment of Animal, Wildlife and Grassland Sciences, UFS, P.O. Box 339, Bloemfontein, 9300, South Africa
IINorthern Cape Department of Agricultural, Land Reform and Rural Development, Private Bag X9, Jan Kempdorp 8550, South Africa
IIIARC-Animal Production Institute, Private Bag X2, Irene, 0062, South Africa
IVDelta G, 145 Ice Cave Rd., Miles City, Montana 59301, USA
ABSTRACT
Fitness is of paramount importance to efficient and profitable beef production. Thus, the objective of this study was to estimate genetic components of fitness traits measured in Afrikaner (A), Brahman (B), Charolais (C), Hereford (H) and Simmentaler (S). For this study, the fitness traits recorded were percentage of cows exposed that were subsequently certified pregnant (PR), percentage of certified pregnant cows that subsequently calved (CR), percentage of calves born that survived to weaning (SV) and the percentage of cows exposed that ultimately weaned a calf (WR). Data were mean performance of straightbred, F1 cross, backcross and three-breed cross females. All crossbred females were of at least 25% A heritage. Breed group means were equated with their genetic expectations assuming recombination effects were nil and the heterosis effects were proportional to the expected heterozygosity in the crosses relative to the purebreds. With the exception of B-sired females from CA cross dams, the genetic model fit the breed group means with a high degree of fidelity. Breed-specific genetic effects tended not to individually exceed the magnitude of their standard errors. However, when the breed-specific genetic effects were combined to predict breed group means, the fitness of crossbred females, on average, exceeded that of their straightbred contemporaries. No particular advantage was noted for adding Brahman to the breed composition of crossbred females with at least 25% Afrikaner heritage. In summary, these data are viewed as being supportive of the use of breed resources in organized crossbreeding systems, such as two- and three-breed rotations that maintain at least 25% Afrikaner germplasm in the breeding females.
Keywords: Afrikaner cattle genotypes, heterosis effects, predicted phenotypic values, weaning rate
Introduction
To develop effective suckler cow replacement strategies, beef producers must have information about breed-specific genetic effects for economically important traits. In dam lines, traits affecting maternal ability and calf production are of primary importance (Roughsedge et al., 2001). A sire line that is appropriate for the ultimate use of progeny should optimize production from the dam line (Weaber, 2010).
Fitness has the highest impact on the profitability of a beef cattle enterprise in that a unit increase in genetic gain obtained in number of calves weaned influences profit to a greater extent than increases or similar magnitude in growth traits, especially in extensive tropical production systems where calving rates are relatively low. For example, MacNeil & Matjuda (2007) developed an aggregated simulation model to facilitate breeding strategies in mating exotic sires with adapted dam lines in a specific-cross production system to produce value-added weaned calves to the feedlot industry. As well as phenotypic traits, they found the relative emphasis on calf survival trait to be equal in importance to direct additive effects on weaning weight in the selection index.
Immediate rapid improvement of low heritable fitness traits may be feasible by exploiting heterosis through crossbreeding (Long, 1980). Although low in heritability, further cumulative and permanent changes in fitness traits may arise through selection based on estimated breeding values EBV (Van der Westhuizen et al., 2001). Optimal crossbreeding systems are predicated on the choice of breeds that contribute to them (Kress & MacNeil, 1999). Thus, the aim of this paper was to estimate breed additive and heterosis effects from a beef-crossing experiment conducted at Vaalharts Research Station in South Africa. A preliminary report of these data was presented in the thesis of the late D. Els (1988).
Materials and Methods
The crossbreeding experiments in which the original data were generated were carried out at Vaalharts Research Station, near Jan Kempdorp, from 1979 to 1981. The research station is located in the centre of South Africa at 27°51' south and 24°50' east, at an altitude of 1175 metres, and is in an area with sandy red soil underlain by limestone. These soils form part of the Hutton formation and represent mainly the Manganese series (Laker, 2003). The veld type is mixed Tarchonanthus veld, Veld type No 16b, 4 (Acocks, 1988). The research station has a recommended carrying capacity of 10 ha/LSU (livestock unit). Climate at Vaalharts is classified as semi-arid. It is characterized by hot summers and cold winters with frost a common occurrence. The highest monthly average temperature is approximately 32 °C and occurs in December and January. The lowest monthly average temperature is approximately -0.5 °C and occurs in July. The average precipitation is approximately 450 mm per annum, of which 88% is experienced during the summer months from October to April in the form of thunderstorms (Els, 1988).
Five purebred sire lines, namely Afrikaner (A), Brahman (B), Charolais (C), Hereford (H) and Simmentaler (S), were evaluated as purebreds, as top-crosses on A as a dam line producing F1 female progeny, and on BA, CA, HA and SA F1 females producing backcross and three-breed cross female progenies. Cows (n = 1820) of all these genotypes were evaluated for fitness traits when joined with Bonsmara sires. The beef cattle herd was raised under extensive conditions. Management and selection procedures of the herd were described by Els (1988). Least squares means for conception rate (PR) (percentage of exposed cows certified pregnant); calving rate (CR) (percentage of cows certified pregnant that calved); calf survival (SV) (percentage of calves born that survived to weaning); and weaning rate (WR) (number of calves weaned as a percentage of the number of cows exposed for breeding) in various breed group combinations were extracted from tables in Els's (1988) thesis. These means were adjusted for contemporary group (year of birth, calving season, age of dam) and sex of calf effects, which were significant (P <0.05) sources of variation for all the traits.
Dickerson (1973) proposed partitioning breed group means into breed-specific additive and heterosis effects as follows:
Pure breeds
C = GlC+ GMC
Two-breed crosses
A x B = 1/2GlA + 1/2GlB + HlAB + GMB
Backcrosses
A x BA = 3/4GlA + 1/4GlB + 1/2HlBA + HMBA + 1/2GMB + 1/2GMA
Three-breed crosses
A x BC = 1/2GlA + 1/4GlB + 1/4GlC + 1/2HlAB + 1/2HlAC + HMBC + 1/2GMB + 1/2GMC
In the above formulas A, B and C designate different breeds; Gland GMrepresent individual additive and maternal effects; and hland Hmrepresent individual and maternal heterosis effects, respectively. Here, recombination effects were assumed to be nil and heterosis effects were assumed to be proportional to expected heterozygosity in the crosses relative to the purebreds and recombination effects were assumed to be nil. Thus, the general model can be stated as:
Y = G0+ βiGl + βjGM + βkΗl+ βlHM + ε
where Y is the phenotypic value of the cross for the trait of interest; G0is the intercept; βiand βjare partial regression coefficients representing individual and maternal additive effects; βkand βlare partial regression coefficients representing individual and maternal heterosis effects and ε is the residual lack of fit (not estimated).
For each trait, the least squares means were equated with their expectations and the resulting system of equations was solved by weighted least squares, in which the weight given to each mean was the reciprocal of its standard error, using the GLM procedure of SAS (2010). In order to uniquely solve the system of equations represented by the model, two constraints were necessary. These were that gla= GMA= 0. Thus, breed-specific individual and maternal additive effects were expressed as deviations from the respective Afrikaner breed effects and the intercept estimated the mean for A. Similar methodology was used by MacNeil et al. (1988) to summarize multiple phases of a crossbreeding experiment and by Williams et al. (2010) to analyse a number of cattle breeds from an extensive literature review of crossbreeding studies. For instance, the equilibrium value (2RAH) for the two breed rotation of A and H is given by:
2RAH= 1G0+ 1/2GlH + 1/2GMH + 2/3HlAH + 2/3HMAH
Here, predicted values for the breed group means presented by Els (1988) were calculated from the regression model given above as a check on the goodness of fit of the model. For each trait, the correlation between the observed breed group means and their predicted values was calculated as a summary statistic.
Results and Discussion
Although the data were collected years ago, it is believed to be reliable and accurate. However, at the time it was not analysed to estimate breed additive and heterosis effects. The mean level of performance for some of the breeds may have changed as a result of response to selection and/or inbreeding that has accrued in the intervening generations and this should be considered in contemporary application of these results by the South African beef industry. However, crossbreeding results have not been collected recently under South African conditions.
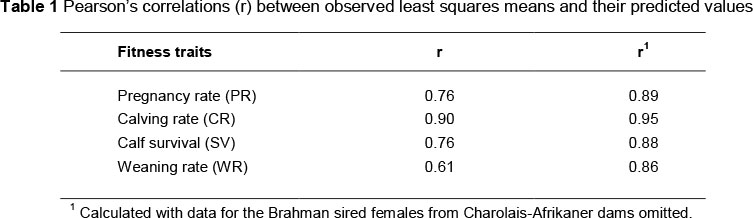
When partitioned into genetic components, direct additive effects explained 22% to 41% and direct heterosis effects explained 30% to 56% of the variation in breed group means across all traits considered in the study. In contrast, maternal additive effects explained less than 11% and maternal heterosis effects explained less than 3% of the variation in breed group means for these fitness traits. The apparent lack of important maternal effects is seemingly consistent with their definition in which the genes of the dam create environmental variation contributing to phenotypic expression in the progeny (Dickerson, 1973) and the expression of these fitness traits as adult females is long removed from the proximity of their dams. Therefore, presentation of these results will focus solely on the direct effects. In addition, there is evidence that intra-breed specific heterosis effects may be important. However, the numbers of observations contributing to the individual breed group means are relatively small with 50% of these classes comprising fewer than 47 observations. Therefore, the authors chose to present the heterosis estimates at the level of sub-species crosses (i.e. indicus vs. sanga vs. taurus) as well. Pearson's correlation coefficients between the least squares means reported by Els (1988) and their predicted values based on the linear model used to partition additive and heterosis effects were calculated to quantify the adequacy of this partitioning (Table 1). These correlation coefficients ranged from 0.61 to 0.90, suggesting some potential for a lack of fidelity between the original data and the predicted values derived from the linear model that was used here. On further investigation, it was discovered that the phenotypic performance of Brahman-sired females from Charolais-Afrikaner dams was fortuitously exceptional and of great influence on the correlations between observed and predicted values. Omitting this single influential breed group from the calculation of the correlation of observed and predicted values markedly increased the fidelity of their relationship (Table 1). Other possible explanations for the departure of these correlations from unity include unknown environmental effects on these traits; genetic effects that were not accounted for in the model (e.g. recombination or epistasis and grand maternal additive effects); and sampling of genetic effects in the breed group means.
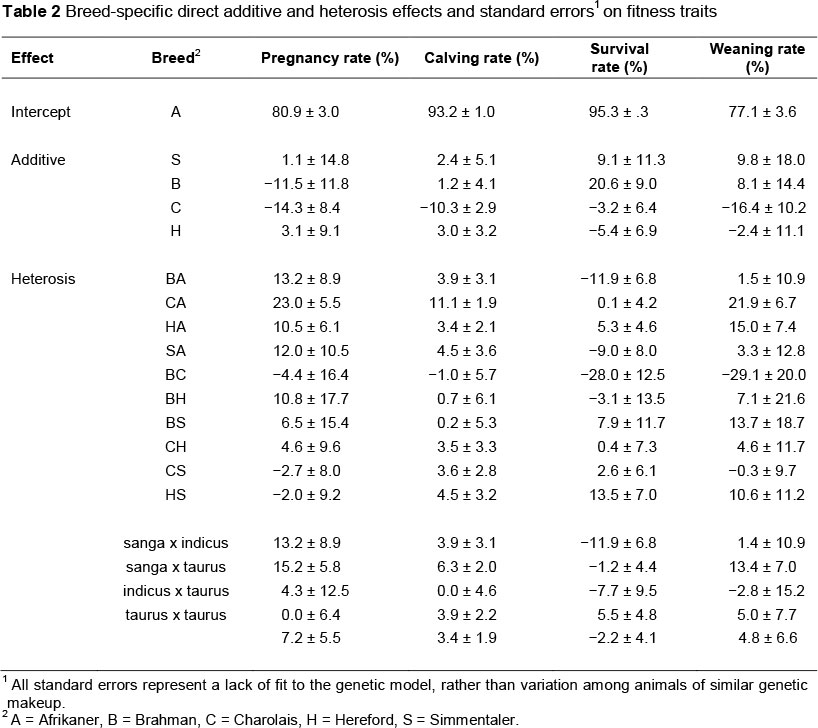
Estimated breed-specific genetic effects for fitness traits are shown in Table 2. These values were estimated from a statistical model that accounts for 20 of the 28 degrees of freedom available in the dataset. Relatively few of the individual genetic effects exceed the magnitude of their standard errors. In these data, power of the test to detect breed-specific genetic effects is low owing to the relatively small sample sizes and substantial variance of fitness traits. Thus, any conclusion that a particular effect is zero has a fairly high probability of being incorrect.
Natural selection has advanced reproduction and survival for many generations. Therefore, the heritability of fitness traits tends to be low, and variation in them among individuals is largely owing to environmental factors (MacNeil et al., 1984; Van der Westhuizen et al., 2001; Weaber, 2009). Still, heterosis effects are expected to improve the productivity of cows (Weaber, 2009), perhaps by reducing the frequency of loci that are homozygous for unfavourable recessive alleles (Charlesworth & Willis, 2009). It is also notable that the conscientious managers attempt to minimize variation in pregnancy rate and calf survival, keeping both at high levels.
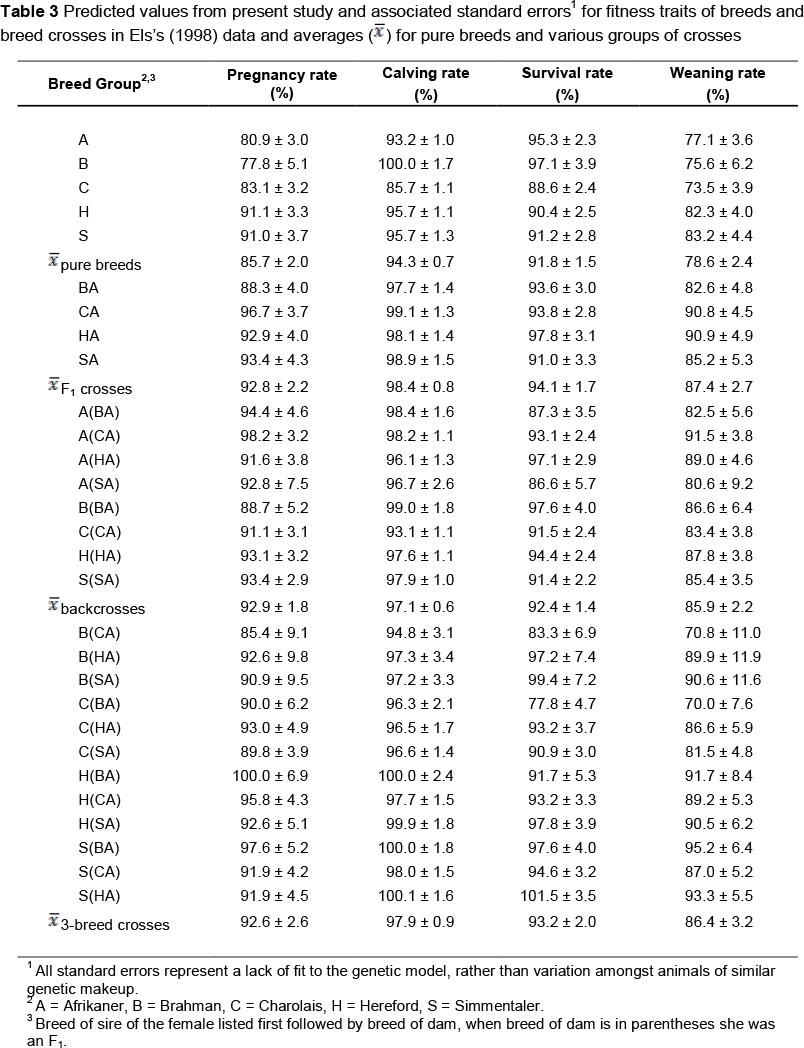
Production utilizes combinations of these effects and not the individual effects. Predicted performance of the breed crosses for the various fitness traits are presented in Table 3. These predicted values have greater precision than the original least squares means, because estimates of the genetic effects of which they are composed are informed by substantially more data from the other breed combinations. Inference from the estimates of genetic effects (Table 2) is not limited to these breed combinations. Rather, an important advantage of partitioning breed effects as described above is that the merit of untested breed combinations can be predicted in a straightforward manner from the results (e.g. MacNeil et al., 1988). One minor disadvantage of the prediction methodology is that the model assumes the dependent variable is unbounded, and thus may give rise to predicted values that exceed biological maxima, as was occasionally observed here (i.e. rates >100%).
The estimated direct effects suggest that as straightbred females, Hereford and Simmentaler would have the greatest fitness. However, crossbred females, on average, had a 7% greater pregnancy rate than their purebred contemporaries. Nineteen of the 24 crossbred breed groups had numerically equal to or greater pregnancy rates than the highest performing purebred females. Departure of the calving rate from 100% is indicative of in utero calf mortality occurring after the pregnancy testing. The greatest in utero calf mortality occurred in purebred Afrikaner and Charolais dams and in 3/4 Charolais 1/4 Afrikaner backcross dams. On average, crossbred dams lost fewer calves between pregnancy testing and parturition than their purebred contemporaries. In the present study, there was no detectable difference in post-natal calf survival to weaning that was attributable to breed type of dam. The net effect on weaning percentage was that crossbred dams outperformed their straightbred contemporaries by 8%. Prayaga (2004) observed significant heterosis for fitness traits in crosses among tropically adapted Bos taurus and Bos indicus cattle, and thus supported the use of crossbreeding in a challenging environment.
For fitness traits, Mpofu (2002) claimed superiority of adapted indigenous cattle and inferiority of exotic cattle not adapted to the environment. However, the present results illustrate the potential for using exotic germplasm as contributors to crossbred beef females. Differences between these studies may result from the semi-arid environment of Vaalharts Research Station being less environmentally stressful than the environment of the Matopos Research Station in Zimbabwe and thus a quarter adapted germplasm was sufficient to confer adaptability. Thus, crossbreeding systems, such as two- and three-breed rotations, which maintain at least 25% Afrikaner germplasm in the breeding females are expected to be viable and efficient in environments similar to Vaalharts Research Station. Results from Skrypzeck et al. (2000) support fractional use of Afrikaner germplasm in the breeding herd.
Brahman cattle are thought to be adapted to environments that are challenging because of heat, humidity and parasites (Greiner, 2009). In the present study, there was no particular advantage to adding the Brahman to the breed composition of crossbred females with at least 25% Afrikaner heritage. Williams et al. (1990) examined rotational crossbreeding systems that incorporated the Brahman breed in the sub-tropical environment of the Gulf Coast of the USA. Straightbred Brahman displayed generally less fitness than straightbred Bos taurus breeds. However, performance of multi-breed rotational crossbreeding systems that incorporated Brahman was consistently greater than the average of the parental breeds. When the data were partitioned into genetic effects (Williams et al., 1991), additive effects of Brahman on the fitness traits were consistently less than those of the Bos taurus breeds. However, indicus x taurus heterosis effects were consistently greater than the taurus x taurus heterosis effects.
Conclusions
Crossbred females had greater fitness than straightbred contemporaries in the environment of Vaalharts Research Station. Effects of pregnancy rate, prenatal survival and postnatal survival interact in determining weaning rate and it is important to choose breeds and breeding systems based on consideration of all components of weaning rate. No particular advantage was noted for adding Brahman to the breed composition of crossbred females with at least 25% Afrikaner heritage. These data are viewed as supportive of the use of adapted and exotic breed resources in organized crossbreeding systems, such as two- and three-breed rotations, that maintain at least 25% Afrikaner germplasm in the breeding females. Some caution is advisable in interpreting specific heterosis effects as the number of observations contributing to each estimate is small. The data did not take cognizance of the genetic trends in the traits and the effect on heterosis parameters in any of the breeds since conducting the crossbreeding experiments.
Acknowledgement
This article forms part of an MSc. study by the first author. This research is supported financially by RMRD SA (Red Meat Research and Development South Africa) and THRIP (Technology and Human Resources for Industry Programme). The dedication of the late D. Els (1988), who conducted the preliminary statistical analyses and summarized this extensive dataset, and the excellent work by researchers at Vaalharts Research Station to collect the data are acknowledged.
References
Acocks, J.P.H., 1988. Veldtypes of South Africa. Memoirs of the Botanical Survey of South Africa. Botanical Research Institute. Pretoria. [ Links ]
Charlesworth, D. & Willis, J.H., 2009. The genetics of inbreeding depression. Nature Rev. Genet. 10, 783-796. [ Links ]
Dickerson, G.E., 1973. Inbreeding and heterosis in animals. In: Proc. Anim. Breed & Genet. Symp. in honor of Dr J.L. Lush, Champaign, Illinois. Amer. Soc. Anim. Sci. 54-77. [ Links ]
Els, D.L., 1988. Kruisteling vir vleisproduksie. PhD thesis, University of the Free State, South Africa. [ Links ]
Greiner, S.P., 2009. Beef cattle breeds and biological types. http://pubs.ext.vt.edu/400/400-803/400-803.html [ Links ]
Kress, D.D. & MacNeil, M.D., 1999. Crossbreeding beef cattle for western range environments, 2nd edition. Western Coordinating Committee 1 Publ. No. TB-99-1. The Samuel Roberts Noble Foundation, Ardmore, OK 73402 USA. [ Links ]
Laker, M.C., 2003. Soil Classification - A desk reference. Ed: Rice, T., Eswaran, H., Stewart, B.A. & Ahrens, R., CRC Press. Chapter 16. Advances in South African soil classification system. Print ISBN: 978-0-8493-1339-4; eBook ISBN: 978-1-4200-4036-4. [ Links ]
Long, C.R., 1980. Crossbreeding for beef production: Experimental results. J. Anim. Sci. 51, 1197-1223. [ Links ]
MacNeil, M.D., Cundiff, L.V., Dinkel, C.A. & Koch, R.M., 1984. Genetics correlations among sex-limited traits in beef cattle. J. Anim. Sci. 58, 1171-1180. [ Links ]
MacNeil, M.D. & Matjuda, L.E., 2007. Breeding objectives for Angus and Charolais specialized sire lines for use in the emerging sector of South African beef production. S. Afr. J. Anim. Sci. 37, 1-7. [ Links ]
MacNeil, M.D., Cundiff, L.V. & Gregory, K.E., 1988. Crossbreeding systems for beef production. Appl. Agric. Res. 3, 44-54. [ Links ]
Mpofu, N., 2002. Comparison of indigenous and foreign cattle for beef production at Matopos Research Station in Zimbabwe. http://agtr.ilri.cgiar.org/casestudy [ Links ]
Prayaga, K.C., 2004. Evaluation of beef genotypes and estimation of direct and maternal genetic effects in a tropical environment. 3. Fertility and calf survival traits. Aust. J. Agric. Res. 55, 811-824. [ Links ]
Roughsedge, T., Thompson, R., Villanueva, B. & Simm, G., 2001. Synthesis of direct and maternal genetic components of economically important traits from beef breed-cross evaluations. J. Anim. Sci. 79, 2307-2319. [ Links ]
SAS, 2010. Statistical analysis system User's Guide: Statistics. SAS Inst., Inc., Cary. N.C., USA. [ Links ]
Skrypzeck, H., Schoeman, S.J., Jordaan, G.F. & Neser, F.W.C., 2000. Estimates of crossbreeding parameters in a mutibreed beef cattle crossbreeding project. S. Afr. J. Anim. Sci. 30, 193-204. [ Links ]
Van der Westhuizen, R.R., Schoeman, S.J., Jordaan, G.F. & Van Wyk, J.B., 2001. Heritability estimates derived from threshold analysis for reproduction and stayability traits in a beef cattle herd. J. Anim. Sci. 31, 25-32. [ Links ]
Weaber, R.L., 2009. Cattle breeds: Considering crossbreeding? http://www.cattlenetwork.com/ [ Links ]
Weaber, B., 2010. Crossbreeding for commercial beef production. In: Beef Sire Selection Manual. http://www.ca.uky.edu/agcomm/PUBS/ASC/asc169 2nd Edition. [ Links ]
Williams, A.R., Franke, D.E., Saxton, A.M. & Turner, J.W., 1990. Two-, three- and four-breed rotational crossbreeding of beef cattle: Reproductive traits. J. Anim. Sci. 68, 1536-546. [ Links ]
Williams, A.R., Franke, D.E. & Saxton, A.M., 1991. Genetic effects for reproductive traits in beef cattle and predicted performance. J. Anim. Sci. 69, 531-542. [ Links ]
Williams, J.L., Aguilar, I., Rekaya, R. & Bertrand, J.K., 2010. Estimation of breed and heterosis effects for growth and carcass traits in cattle using published crossbreeding studies. J. Anim. Sci. 88, 460-466. [ Links ]
Received 6 May 2013
Accepted 24 October 2014
First published online 9 November 2014
# Corresponding author: atheunissen@ncpg.gov.za


{kind=link}
{kind=link}
{kind=link}