










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.44 n.2 Pretoria Feb. 2014
Effects of in ovo administration of amino acids on hatchability and performance of meat chickens
T.M. ShafeyI, #; A.H. MahmoudII; A.A. AlsobayelI; M.A. AbouheifI
IDepartment of Animal Production, College of Food and Agriculture Sciences, King Saud University, Riyadh, Saudi Arabia
IIDepartment of Zoology, College of Science, King Saud University, Riyadh, Saudi Arabia
ABSTRACT
The effects of in ovo amino acid administration on hatchability traits and post-hatch performance were studied using eggs from a meat-type breeder flock (Ross) at 38 weeks of age. On day 15 of incubation, five treatments were applied, consisting of six replicates with 16 eggs per replicate. The treatments consisted of a non-injected negative control; a positive control in which saline was injected; and three treatments with saline, containing 23.72 mg lysine, glutamine, glycine and proline (AA1); 23.60 mg arginine, glutamine, glycine and proline (AA2); and 28.76 mg lysine, arginine, glutamine, glycine and proline (AA3). The hatchability traits were incubation period, chick weight at hatch as an absolute value (CWT) and chick weight at hatch as a percentage of egg weight (CWTP), and the post-hatch performance traits were body weight gain (BWG), feed intake (FI) and feed conversion ratio (FCR). Measurements on the chickens were taken during the starter (1 - 21 days of age), finisher (22 - 35 days of age) and entire (1 - 35 days of age) experimental periods. In ovo amino acid treatments improved CWTP without affecting hatchability traits, and increased FI without altering the FCR of chickens during the entire experimental period. Chickens of the in ovo AA1 treatment had a higher FI than those of the control treatments during the finisher period. In ovo AA2 treatment increased CWT and BWG of the chickens during the starter period and entire experimental period and FI during the finisher period, when compared with those of the control treatments. In ovo AA2 treatment resulted in a longer period of incubation than those of other treatments. It was concluded that in ovo amino acid treatment increased CWTP and FI of chickens without affecting hatchability traits and feed efficiency and in ovo of AA2 treatment increased the incubation period, CWT and BWG when compared with those of control treatments during the entire experimental period.
Keywords: Chicks, hatchability, incubated eggs, in ovo feeding
Introduction
The nutrient content of the hatching egg influences the development and growth of embryos during incubation and the post-hatch performance of chicks (Al-Murrani, 1982; Shafey et al., 2013). The administration of nutrients into the hatching eggs is called in ovo feeding (Uni et al., 2005). The overall aim of in ovo feeding is to supply adequate nutrients to the growing embryo. In ovo feeding improves the hatchability of eggs, the viability and weight of chicks at hatch, intestinal development, growth and feed efficiency of hatched chicks (Gore & Qureshi, 1997; Tako et al., 2004; Shafey et al., 2012), body weight at marketing age, and carcass quality (Hajihosaini & Mottaghitalab, 2004; Selim et al., 2012). Chicken embryos develop and grow in a predetermined nutrient environment. The availability and reserves of nutrients in the egg determine the glycogen stores and subcutaneous fat depots of hatching chicks (Speake et al., 1998; Uni et al., 2005; Shafey et al., 2010), and consequently the development and metabolic transition for digestion and metabolism of exogenous feed nutrients (Moran, 2007). Therefore, providing sufficient nutrients in the egg is a good starting point for the hatching chicks.
An egg contains substantial amounts of amino acids for embryonic growth and development in the yolk and albumin. The embryo relies on the metabolism of carbohydrates and protein in the first two weeks of life, and utilizes amino acids for tissue growth at a higher rate during the second half of incubation. All amino acids are required for tissue synthesis. If there is a shortage of a particular amino acid, others are wasted (Schaible & Patrick, 1980). A deficient supply of amino acids could reduce embryonic growth and development. Hens fed a diet that was low in crude protein or deficient in amino acids laid fewer and smaller eggs (Keshavarz & Jackson, 1992). Sunny & Bequette (2010) suggested that at a late stage of incubation some amino acids may not adequately meet embryonic requirements for development, especially in smallsized eggs. Ohta et al. (1999) suggested that amino acid concentrations in the egg, such as glycine (Gly) and proline (Pro), do not seem sufficient to support embryonic development during the final stage of incubation. A deficiency of amino acids would impair not only protein synthesis, but also the homeostasis of the body. Therefore the synthesis of these amino acids must occur from other substrates. This confirms that available amounts of limiting amino acids are extremely important and that in ovo administration of amino acids at a late stage of incubation may enhance the amino acid status of the chicken embryo (Ohta et al., 1999). In ovo supplementation of amino acids such as arginine (Arg) and glutamine (Gln) can be utilized to synthesize other amino acids and consequently provide adequate amino acids to maximize the potential growth of the embryo. In ovo administration of arginine into Japanese quail embryos increased hatchability (Al-Daraji et al., 2012). Al-Asadi (2013) reported that in ovo administration of arginine and lysine (Lys) at 18 d of age increased hatchability, chick weight at hatch and at 42 d of age, as well as feed intake (FI) of broiler chickens. In ovo feeding of glutamine to chicken embryos was investigated by Pedroso et al. (2006), who found that in ovo feeding of glutamine did not influence chick weight at hatch, as an absolute value (CWT), but reduced hatching time. However, Tavassoli et al. (2011) reported that in ovo administration of glutamine increased CWT.
An adequate amount of amino acids must be available to meet the increased physiological demands of the embryo, especially in the last stage of incubation. Al-Murrani (1982) demonstrated that injecting an amino acid mixture identical to the amino acid profile of egg protein into growing embryos from broiler breeder eggs on the seventh day of incubation resulted in a higher chick weight at hatch and at 56 d of age. In addition, Ohta et al. (1999) injected 53 mg amino acids into broiler breeder eggs at 7 d of age and observed that the ratio of body weight of hatched chicks to egg weight improved. Furthermore, Bhanja & Mandal (2005) reported that in ovo injection of certain amino acids (isoleucine (Ile) + leucine (Leu) + valine (Val) or (Gly +Pro)) resulted in higher body weight at the third week of age.
This study was designed to investigate the effects of administration of a mixture of amino acids ((Lys + Gln + Gly + Pro); (Arg + Gln + Gly + Pro); (Arg + Lys + Gln + Gly + Pro)) at a late stage of incubation on hatchability traits and performance of hatching chicks.
Materials and Methods
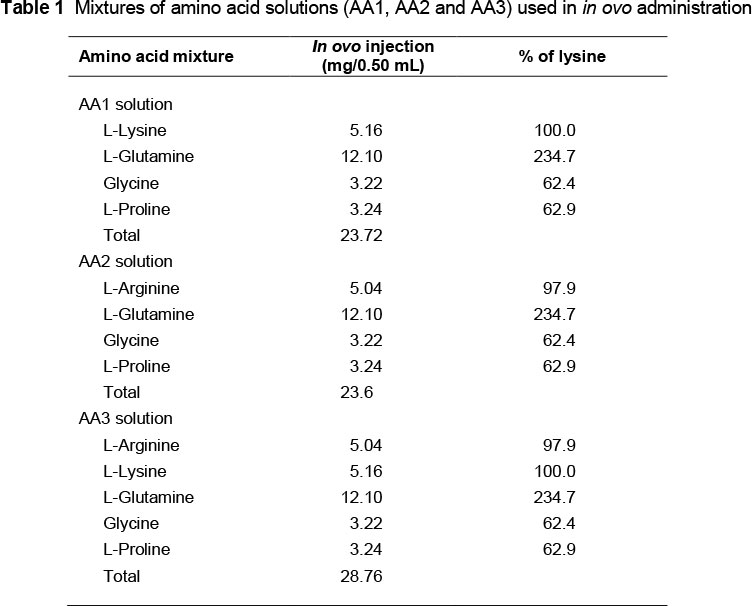
A total of 600 fresh laid eggs produced by a meat-type breeder Ross flock at 38 weeks of age (Al-Wady Company, Al-Riyadh, Saudi Arabia) were used in this trial. Eggs were numbered and weighed individually. Eggs were set in a Maino force-draft incubator (Model II, Maino Enrico Co., Rome, Italy) and incubated at 37.5 °C (99.5 °F) and 55% relative humidity. The eggs were turned every 2 h. Eggs were examined by candling at d 15 of incubation, and infertile eggs and those that contained dead embryos were removed. After examination, 480 eggs were evenly assigned to 30 replicates of 16 eggs of equal weight per replicate. Six replicates were randomly assigned to each of the five experimental treatments. There were two control treatments: non injected (negative control), and injected with 0.9% sterilised saline (positive control); and three experimental treatments with sterilised saline containing a mixture of amino acids: treatment AA1 (Lys + Gln + Pro), treatment AA2 (Arg + Gln + Gly + Pro) and treatment AA3 (Arg + Lys + Gln + Gly + Pro), as shown in Table 1. The amino acids were provided by Merck, Darmstadt, Germany. The concentrations of amino acids in the in ovo injection solutions were formulated according to the amino acid composition of albumin (Belitz et al., 2009).
The wide end of the eggshell was disinfected with 70% isopropyl alcohol and pierced for an injection hole (Shafey et al., 2010). A 0.50 mL solution was injected into the yolk sac using a 25-mm 21-G needle. The 0.50 mL contained 23.72 mg, 23.60 mg and 28.76 mg amino acids for the AA1, AA2 and AA3 treatments, respectively. After injection, the hole was sealed with melted paraffin wax, and the eggs were returned to the incubator. The eggs were transferred to separate compartments in the hatching tray on the morning of d 19 of incubation, for chick identification at hatch. The hatching tray was divided into compartments with thin sheets of mesh wire. The hatching compartment was set at 37 °C (98.6 °F) and 65% relative humidity until the end of d 21 of incubation, at which time chicks, pips (unhatched eggs with live or dead chicks) and dead embryos (unhatched eggs with unbroken shell) were counted. Hatchability percentage was calculated based on the number of hatched chicks as a percentage of the in ovo treated eggs per replicate. Hatching time was recorded every 12 h. Chicks were observed at 12-h intervals from d 18.5 to d 21 (444 to 504 h) of incubation, and hatching weight was recorded to the nearest 0.1 g.
At the end of the incubation period of 21 days, as is the practice in the industry, a total of 35 chicks from each of the five experimental treatments, representing the mean body weight, were assigned to seven replicates of five birds each. Birds were housed in electrically heated battery cages, and offered a commercial starter diet containing 210 g protein/kg and 13.0 MJ metabolizable energy (ME)/kg to 21 d (starter period), followed by a commercial finisher diet containing 190 g protein/kg and 13.4 MJ ME/kg (Arasco, Riyadh, Saudi Arabia) until the termination of the experiment at 35 d of age (finisher period). Feed and fresh water were available ad libitum at all times.
Measurements were made of hatchability percentage and hatchability failures (pips with live embryos, pips with dead embryos, dead embryos), CWT expressed on the basis of absolute value (g) or as a percentage of egg weight (CWTP), hatching period. Post-hatch performance of chickens were body weight gain (BWG), feed consumption (FI) and feed conversion ratio (FCR). All statistical analysis was performed with the Statistical Analysis System (SAS, 2006). All data were checked for normality before analysis. All percentage data were transformed using arc sine square root percentage transformation before analysis. Data were subjected to a one-way ANOVA analysis using the GLM procedure. The statistical model was as follow:
Yij = µ + Fi + Eij
where Yij = dependent variable
µ = The overall mean
Fi = The fixed effect of in ovo treatment
Eij= The experimental error.
When significant variance ratios were detected, differences between treatment means were tested using the least significant difference (LSD) procedure.
Results
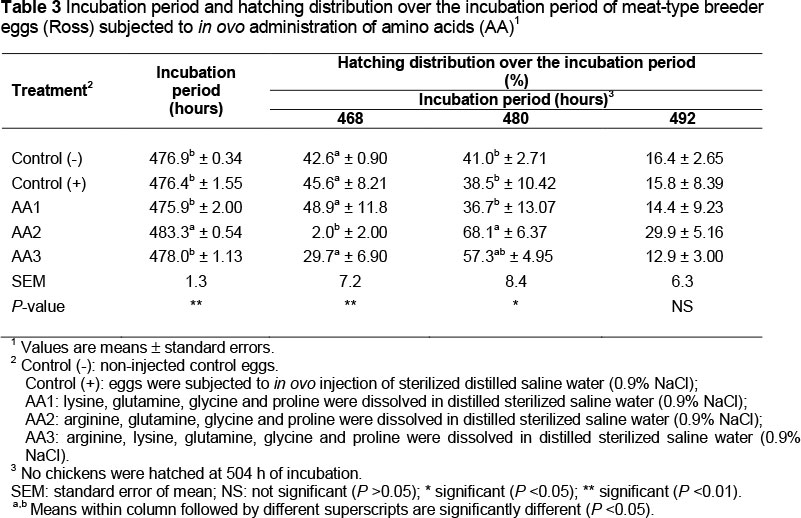
The effects on hatchability traits, CWT, incubation period and hatching distribution over the incubation period of the in ovo administration of amino acids into meat-type breeder eggs are shown in Tables 2 and 3, respectively.
Results from this study indicated that in ovo administration of a mixture of amino acids (23.72 mg of AA1 (Lys, Gln, Gly and Pro), 23.60 mg of AA2 (Arg, Gln, Gly and Pro) or 28.76 mg of AA3 (Arg, Lys, Gln, Gly and Pro)) at 15 days of incubation significantly (P <0.01) improved CWTP without affecting hatchability traits when compared with those of the non injected (negative) and saline injected (positive) control treatments, respectively (Table 2). In ovo administration of the AA2 treatment increased (P <0.01) the incubation period and reduced the percentage of hatched chicks at 468 h of incubation when compared with those of other treatments. The AA2 treatment increased (P <0.05) the proportion of hatched chicks at 480 h of incubation and CWT as an absolute value when compared with those of the control treatments (Table 3).
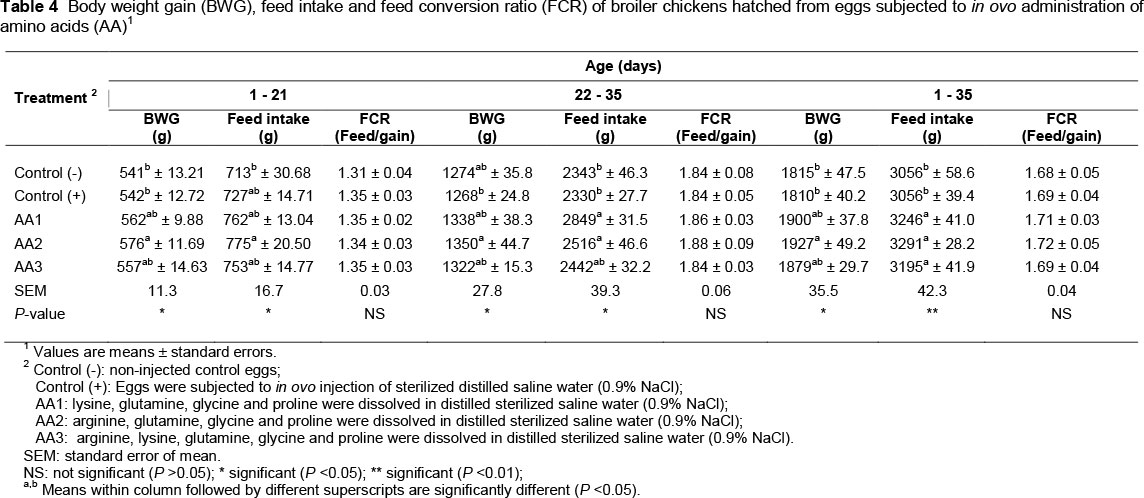
The effects of in ovo amino acid administration of eggs on post-hatch performance of chickens are shown in Table 4. In ovo administration of amino acids (AA1, AA2 and AA3) resulted in a significantly (P <0.01) higher FI of chickens during the entire experimental period (1 - 35 d of age) when compared with those of the control treatments (Table 4). However, FI of chickens differed among in ovo amino acid treatments during the starter and finisher periods of the experiment. Chickens of AA1 treatment had a higher (P <0.05) FI than those of the control treatments during the finisher period of the experiment. Chickens of AA2 treatment had higher (P <0.05) BWG during the starter and entire experimental period, and FI during the finisher period when compared with those of the control treatments. The BWG of chickens at 35 days of age of AA2 treatment was higher (P <0.01) than the average of those of the control treatments by approximately 6.3%. However, the BWG of chickens at 35 days of age of AA1 and AA3 was not significantly different from that of the control treatments. The BWG of chickens at 35 days of age of AA1 and AA3 was higher than average of those of the control treatments by approximately 4.8% and 3.7%, respectively, though non-significantly. Chickens of the AA2 treatment had a higher (P <0.05) BWG during the finisher period than the positive control and higher FI during the starter period than the negative control.
There was no significant difference among treatments in hatchability traits (percentages of hatchability, dead embryos and pipped embryos), percentage of hatched chicks at 492 h of incubation and feed efficiency of chickens during starter, finisher and the entire experimental periods.
Discussion
The finding that in ovo administration of a mixture of amino acids (AA1, AA2 and AA3) at 15 days of incubation did not influence the hatchability of eggs, but increased relative chick weight at hatch to egg weight, is in agreement with Ohta et al. (1999), who found that in ovo administration of 53 mg of amino acids improved relative chick weight at hatch to egg weight without affecting hatchability. These findings suggest that the amino acid content of the egg was sufficient to support the hatching process, but not to maximize embryonic growth. Furthermore, in ovo administration of amino acid mixture increased the amino acid content of the egg for embryonic growth and consequently chick weight at hatch and at 35 days of age. This suggestion is supported by Ohta et al. (2001), who reported that in ovo administration of amino acids increased amino acid utilization and concentration in chicken embryos and other egg contents. Embryos may utilize in ovo administered nutrients to improve energy status and save muscle protein from mobilization, to improve enteric development, and to provide energy for hatching and subsequent growth (Tinker et al., 1986; Tako et al., 2004; Tong & Barbul, 2004; Uni et al., 2005; Foye et al., 2006; Shafey et al., 2010). A significant amount of both essential and non-essential amino acids remains unused in the egg by day 19 of incubation (Ohta et al., 1999). However, during the final stage of incubation, the concentrations of some amino acids, such as glycine and proline, do not seem to meet the requirements of the embryo. Glycine and proline are involved in collagen production (Murray et al., 2006). Since collagen is essential for tissue development, these amino acids need to be available or synthesized from other precursor amino acids that are present in higher concentrations in the egg.
The finding that AA2 treatment improved the weight of chickens at hatch (as an absolute value and as a relative weight to egg weight) and at 35 days of age when compared with those of the control treatments suggests that the amino acid mixture of the AA2 treatment might be required at a higher concentration for body weight at hatch and at marketing age. This finding suggests that the positive effect of in ovo administration of AA2 was not limited to the CWT, but continues to improve chick weight up to 35 days of age. However, the positive effects of the AA1 and AA3 treatments were limited to the relative chick weight at hatch to egg weight. The BWG at 35 days of age of the AA1 and AA3 treatments was not significantly different from that of AA2, though lower, albeit non significantly. These findings are probably related to the contribution of amino acid in ovo treatments to the available amino acids of the embryo and hatching chick. The composition of amino acids in the AA1, AA2 and AA3 in ovo treatments indicated that amino acids in the in ovo treatments differed in the arginine : lysine ratio. According to Al-Asadi (2013) in ovo administration of arginine and lysine into chicken embryos increased hatchability, chick weight at hatch and at 42 d of age, as well as FI of broiler chickens. The arginine : lysine ratio (mg) in AA1, AA2 and AA3 was 0 : 5.16, 5.04 : 0 and 5.04 : 5.16, respectively. AA2 had a higher arginine : lysine ratio than AA1 and AA3. The additional arginine probably supported chick weight at 35 days of age as a result of the increasing arginine : lysine ratio remaining in the peritoneal cavity post hatching (Romanoff, 1960), which might have enhanced the protein synthesis after hatching. This finding is in agreement with Al-Daraji et al. (2012), who found that in ovo administration of arginine into Japanese quail embryos increased BWG at 7 and 42 days of age. The finding that amino acids in the in ovo treatment increased FI at 35 d of age without altering the FCR of chickens is in agreement with Bhanja & Mandal (2005), who found that higher body weight and FI were observed in the amino acid-injected groups (Lys + Arg; Lys + methionine + cysteine ; threonine + Gly + serine ; Ile + leu + Val; Gly + Pro) and (Ile + Leu + Val; Gly + Pro) when compared with the control group. In addition, Bakyaraj et al. (2012) reported that in ovo injection of a mixture of amino acids (Lys + Meth + Arg + Leu + Ile) resulted in significantly higher body weight and FI with no variation in FCR of chickens when compared with those of the untreated control. However, the non-significant effect of in ovo treatment on FCR in this study was not in agreement with Al-Daraji et al. (2012), who found that in ovo administration of arginine into Japanese quail embryos increased their FCR.
It was concluded that in ovo administration of amino acid mixtures of 23.7 mg of Lys + Gln + Gly + Pro; 23.6 mg of Arg + Gln + Gly + Pro; and 28.76 mg of Arg + Lys + Gln + Gly + Pro improved relative chick weight at hatch to egg weight without altering hatchability traits and increased FI between 1 d and 35 d of age without altering feed conversion ratio. In ovo administration of 23.6 mg of arginine, glutamine, glycine and proline increased incubation period, CWT when expressed as an absolute value and BWG at 35 d of age. There was no significant difference among the three amino acid mixtures in hatchability traits, CWT and chicken performance during the experimental period.
Acknowledgements
The authors extend their appreciation to the Deanship of Scientific Research at King Saud University for funding the work through the research group project No RGP-VPP- 282.
References
Al-Asadi, A.N.O., 2013. Effect of early feeding (in ovo injection) amino acids on hatchability, some productive and physiology traits of broiler. Int. J. Sci. Tech. 8, 6-13. [ Links ]
Al-Daraji, H.J., Al-Mashadani, A.A., Al-Hayani, W.K., Al-Hassani, A.S. & Mirza, H.A., 2012. Effect of in ovo injection with L-arginine on productive and physiological traits of Japanese quail. S. Afr. J. Anim. Sci. 42, 139-145. [ Links ]
Al-Murrani, W.K., 1982. Effect of injecting amino acids into the egg on embryonic and subsequent growth in the domestic fowl. Br. Poult. Sci. 23, 171-174. [ Links ]
Bakyaraj, S., Bhanja, S.K., Majumdar, S. & Dash, B., 2012. Modulation of post-hatch growth and immunity through in ovo supplemented nutrients in broiler chickens. J. Sci. Food Agric. 92, 313-320. [ Links ]
Belitz, H.D., Grosch, W. & Schieberle, P., 2009. Egg. In: Food Chemistry. Eds: Belitz, H.D., Grosch, W. & Schieberle, P., Heidelberg: Springer. pp. 546-561. [ Links ]
Bhanja, S.K. & Mandal, A.B., 2005. Effect of in ovo injection of critical amino acids on pre and post-hatch growth, immunocompetence and development of digestive organs in broiler chickens. Asian-Austral. J. Anim. Sci. 18, 524-531. [ Links ]
Foye, O.T., Uni, Z., McMurtry, J.P. & Ferket, R.P., 2006. The effects of amniotic nutrients administration, in ovo feeding of arginine and/or p-hydroxy p-methylbutyrate (HMB) on insulin-like growth factors, energy metabolism and growth in turkey poults. Int. J. Poult. Sci. 5, 309-317. [ Links ]
Gore, A.B. & Qureshi, M.A., 1997. Enhancement of humoral and cellular immunity by vitamin E after embryonic exposure. Poult. Sci. 76, 984-991. [ Links ]
Hajihosaini, M. & Mottaghitalab, M., 2004. Effect of amino acid injection in broiler breeder eggs on hatchability and growth of hatched chicken. J. Agric. Sci. 1, 23-32. [ Links ]
Keshavarz, K. & Jackson, M.E., 1992. Performance of growing pullets and laying hens fed low-protein, amino acid supplemented diets. Poult. Sci. 71, 905-918. [ Links ]
Moran Jr, E.T., 2007. Nutrition of the developing embryo and hatchling. Poult. Sci. 86, 1043-1049. [ Links ]
Murray, K.R., Granner, D.K. & Rodwell, V.W., 2006. Metabolism of proteins and amino acids. In: Harper's Illustrated Biochemistry. Eds:. Appleton and Lange McGraw Hill, New York, USA. pp. 241-245. [ Links ]
Ohta, Y., Tsushima, N., Koide, K., Kidd, M.T. & Ishibashi, T., 1999. Effect of amino acid injection in broiler breeder egg on embryonic growth and hatchability of chicks. Poult. Sci. 78, 1493-1498. [ Links ]
Ohta, Y., Kidd, M.T. & Ishibashi, T., 2001. Embryo growth and amino acid concentration profiles of broiler breeder eggs, embryo and chicks after in ovo administration of amino acids. Poult. Sci. 80, 1430-1436. [ Links ]
Pedroso, A.A., Chaves, L.S., Lopes, K.L.A.M., Leandro, N.S.M., Cafe, M.B. & Stringhini, J.H., 2006. Nutrient inoculation in eggs from heavy breeders. Rev. Bras. Zootec. 5, 2018-2026. [ Links ]
Romanoff, A.L., 1960. The Avian Embryo. The Macmillan Publishing Company, New York. pp. 1-1305. [ Links ]
SAS, 2006. Statistical analysis System, 2006. SAS Institute Inc, Cary, N.C., USA. [ Links ]
Schaible, P.J. & Patrick, H., 1980. Poultry: Feeds and Nutrition. 2nd ed. AVI Publishing Company, Inc., Westport, Connecticut, USA. [ Links ]
Selim, Sh.A., Gaafar, K.M. & El-ballal, S.S., 2012. Influence of in-ovo administration with vitamin E and ascorbic acid on the performance of Muscovy ducks. Emir. J. Food. Agric. 24, 264-271. [ Links ]
Shafey, T.M., Al-Batshan, H.A., Al-Owaimer, A.N. & AL-Samawei, K.A., 2010. Effects of in ovo administration of L-carnitine on hatchability performance, glycogen status and insulin-like growth factor-1 of broiler chickens. Br. Poult. Sci. 51, 122-131. [ Links ]
Shafey, T.M., Alodan, M.A., Al-Ruqaie, I.M. & Abouheif, M.A., 2012. In ovo feeding of carbohydrates and incubated at a high incubation temperature on hatchability and glycogen status of chicks. S. Afr. J. Anim. Sci. 42, 210-220. [ Links ]
Shafey, T.M., Sami, A.S. & Abouheif, M.A., 2013. Effects of in ovo feeding of L-glutamine on hatchability performance and hatching time of meat-type breeder eggs. JAVA 12, 135-139. [ Links ]
Speake, B.K., Murray, A.M.B. & Noble, C.R., 1998. Transport and transformations of yolk lipids during development of the avian embryo. Prog. Lipid Res. 37, 1-32. [ Links ]
Sunny, N.E. & Bequette, B.J., 2010. Gluconeogenesis differs in developing chick embryos derived from small compared with typical size broiler breeder eggs. J. Anim. Sci. 88, 912-921. [ Links ]
Tako, E., Ferket, P.R. & Uni, Z., 2004. Effects of in ovo feeding of carbohydrates and beta-hydroxy-beta-methyl butyrate on the development of chicken intestine. Poult. Sci. 83, 2023-2028. [ Links ]
Tavassoli, M., Mousavi, S.N. & Abedini, M.R., 2011. Effects of in ovo feeding of glutamine on performance, small intestine morphology and immune response of broiler chicks. J. Anim. Environ. 3, 1-6. [ Links ]
Tinker, D.A., Brosnan, J.T. & Herzberg, G.R., 1986. Interorgan metabolism of amino acids, glucose, lactate, glycerol and uric acid in the domestic fowl (Gallus domesticus). Biochem. J. 240, 829-836. [ Links ]
Tong, B.C. & Barbul, A., 2004. Cellular and physiological effects of arginine. Mini Rev. Med. Chem. 4, 823-832. [ Links ]
Uni, Z., Ferket, P.R., Tako, E. & Kedar, O., 2005. In ovo feeding improves energy status of late-term chicken embryos. Poult. Sci. 84, 764-770. [ Links ]
Received 14 June 2013
Accepted 8 April 2014
First published online 21 May 2014
# Corresponding author: tmshafey@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}