










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.44 no.2 Pretoria Fev. 2014
Index-in-retrospect and breeding objectives characterizing genetic improvement programmes for South African Nguni cattle
L.E. MatjudaI, II; M.D. MacNeilI, III, IV; A. MaiwasheI, IV, #; V.R. LeesburgV; M. MalatjeII
IARC-Animal Production Institute, Private Bag X2, Irene, 0062, South Africa
IIUniversity of Limpopo, Private Bag X1106, Sovenga, 0727, South Africa
IIIDelta G, 145 Ice Cave Road, Miles City, Montana, 59301, USA
IVUniversity of the Free State, PO Box 339, Bloemfontein, 9300, South Africa
VUSDA-Agricultural Research Service, 243 Fort Keogh Road, Miles City, Montana, 59301, USA
ABSTRACT
The goals of this research were to document the historical selection applied by Nguni breeders that contributed to the national genetic evaluation scheme, and to offer prospective breeding objectives to guide future selection practices. Genetic selection differentials, weighted by number of offspring, and unweighted, were calculated for the recorded traits, and were used to quantify differences between natural and imposed selection. Index-in-retrospect methods were used to quantify differences between the traits in selection that were applied. Economic values for the prospective breeding objectives were derived through simulation. Results indicate that relatively little directional selection has been applied to Nguni as a breed, and that historically bulls and heifers were selected according to divergent criteria. Breeding objectives indicated that production, profitability and sustainability goals were largely complementary, but reflected a change from past selection practices. Breeders are advised to be intentional and consistent in the application of selection criteria.
Keywords: Breeding goals, production systems, selection, sexual dimorphism simulation
Introduction
Nguni cattle have been characterized as having high fertility, low maintenance requirements, ease of calving, adaptability, resistance to parasites, resistance to tick-borne diseases, good temperament, longevity, browsing and good walking ability (Scholtz, 1988; South African Livestock Breeding, 2004). Consequently, the breed is viewed as having cost-effective capacity for beef production and the ability to thrive under harsh conditions. These characteristics seemingly position Nguni for use as a specialized dam line and it has long been promoted as a maternal breed. The local environment, cultures and values of communities of emerging farmers are reflected in the key traits and functions of Nguni cattle (Rege, 2001). Nguni cattle are adapted to local environmental stresses, are able to provide traction, and produce meat and milk. Behind these traits are the breeding practices and other indigenous knowledge systems that have been applied to maintain these populations, and the genetic diversity that they represent. However, historical selection practices with respect to economically relevant traits have not been well documented.
Selection is the primary force for changing the additive genetic merit of herds and breeds. Selection, which manifests as differential reproduction of individuals, can occur at any stage of the lifecycle, and be the result of natural causes or directed by human beings (Koch et al., 1974). For single trait selection, genetic change is predicted by observed selection differentials multiplied by heritability. However, if multiple traits are involved, this simple prediction is inappropriate (Magee, 1965). The observed cumulative selection differentials of parents are a function of multiple trait relationships in the population over time (Koch et al., 1994). Therefore, change from selection depends on selection differentials in primary and correlated traits; the fraction of selection differentials because of average genetic differences of parents; and the interval between selected generations. Index-in-retrospect methods (Dickerson et al., 1954) can provide insight into the traits that were historically used as selection criteria and the emphasis placed on them in deciding which animals ultimately became parents.
The critical question of what exactly constitutes improvement requires attention (MacNeil et al., 1997). In pursuit of genetic improvement, an obvious, though far from trivial problem in defining the objectives is to identify the economic values of traits that affect the breeding goal. However, while James (1982) has suggested that the breeder's most important decision is the choice of breeding objective, he cautions that highly efficient selection for the wrong objective may be worse than no selection at all. The existence of industry-wide specifications for beef products, production systems, and therefore different enterprises does not necessarily suggest that there should be an industry-wide selection index. Resources for production and level of production vary among production systems and units, resulting in different economic structures. As a result, relative economic values may differ among production systems and units, and each may have a numerically different selection index. However, even across production and marketing systems as widely divergent as those in South Africa and in the US, the correlation between breeding objectives for Angus as terminal sires was found to be positive and substantial (MacNeil, 2005a). Additionally, correlations among breeding objectives for terminal sires across breeds and marketing systems in the US ranged from 0.74 to 0.98 (MacNeil, 2005b). Thus, where there is general agreement on the purpose of a breed, such as Nguni, the direction could be set. This direction is captured in a breeding objective.
Therefore, the first objective of the current study was to evaluate past selection for performance traits imposed on South African Nguni cattle using index-in-retrospect. The second was to develop prospective breeding objectives for a straight-bred Nguni production system typical of the communal and emerging sectors of the South African cattle industry. Finally, the aim was to ascertain the consistency of breeding objectives reflective of the phases of agricultural development (i.e. production, profitability, and sustainability), as identified by Bawden (1989).
Materials and Methods
Data used in the current study were estimated breeding values (EBVs) for animals born from 1973 to 2006. These EBVs were obtained from an official routine national genetic evaluation of the South African Nguni breed conducted by the Agricultural Research Council (ARC). The data included EBVs for birth weight direct (BWTd), birth weight maternal (BWTm), weaning weight direct (WWTd), weaning weight maternal (WWTm), yearling weight (YWT), 18-month weight (18WT), average daily gain (ADG), shoulder height (SDH), scrotal circumference (SCR), body length (LEN), mature weight (MWT) and calf tempo (CT). These data are analysed by the ARC with a series of multiple trait models (Table 1). With the exception of CT, all EBVs are expressed on a per animal basis. Calf tempo is expressed per 100 daughters, and thus the results obtained from the national evaluation were divided by 100 to express all EBVs and results derived from them on a per-animal basis.
Index-in-retrospect
The EBVs were used to calculate the genetic selection differentials (Δgi) required to determine the weights of an index-in-retrospect. Those animals that subsequently became parents (that is, were selected) were identified and their individual selection differentials were calculated as the difference between their EBVs and the average EBV of the population (Van Vleck, 1993). The genetic selection differentials were calculated separately, and accumulated over the four selection paths for registered sires and dams. The index-in-retrospect may have been used for selection to produce gains that have occurred, even though the weights for the index may have been unknown at the time of selection (Dickerson et al., 1954). The index-in-retrospect is given by:

where I is the underlying unknown index. Letting D be the selection intensity factor, the regression of EBVi on I will give the expected genetic selection differential for trait i:
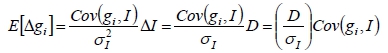
If D/σI (is a constant for all the traits in the index) is set to 1, the expected values of the genetic selection differentials are:

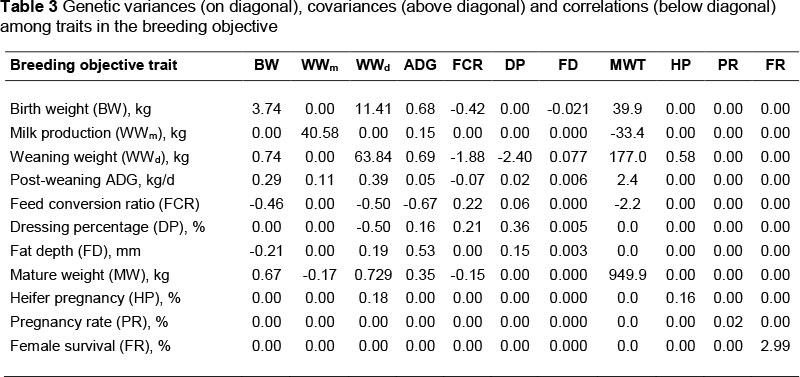
The weights for the underlying index (wi) can be estimated by equating the estimated genetic selection differentials to the right hand sides of the above equations as: w = G-1Δg (Van Vleck, 1993). The genetic variance-covariance matrix (G) (Table 2) was previously generated by the genetic evaluations done by the ARC, using REML.
Simulation model
A lifecycle simulation starting with 1000 Nguni heifer calves at weaning was used to calculate breeding objectives applicable to an integrated production system using Nguni cattle. The simulation was constructed at whole-animal level and included continuous time-dependent processes, events, and stochastic and deterministic elements. Parameters of the simulation were based on phenotypic data gathered in the South African National Beef Cattle Recording and Improvement Scheme (NBCRIS).
The mental model of the simulated system can be described as follows. At time (yr) 0.0, the simulation was initiated with 1000 weaned heifer calves. These heifer calves may be thought of as a group of "replacement" females. Expense and income were accumulated over the lifecycle of this set of females. Sources of income are sale of females culled for infertility, sale of progenies marketed as beef carcasses after feedlot feeding, and sales of any surviving cows at time = 12.0 on termination of the simulation. Sources of expense are harvested and grazed feeds consumed by the cows and their progeny, and fixed per-head costs associated with each female and each feedlot fed progeny. Lifecycle profitability was calculated at the conclusion of the simulation as the difference between total income and total expense.
The following general model was used to simulate mean performance for maternally influenced traits (Dickerson, 1969):

where P = level of phenotypic performance, µ = constant base level of performance, ad = direct genetic effect of Nguni on performance and am = maternal genetic effect of Nguni on performance. For traits that were assumed not to be influenced by maternal effects, the am term was set to zero.
Weight for Nguni females was simulated from their average growth curve as fitted to data from the NBCRIS. Specifically, the growth curve was derived from individual weight measurements taken at birth, weaning, yearling, 18 months and maturity.
Simulated milk production assumed proportionality of Hereford and Nguni maternal effects on weaning weight from the NBCRIS and level of milk produced by Hereford cows (1062 kg/lactation) as measured by MacNeil & Mott (2000). The simulated milk production of cows <5 yr old was adjusted relative that of mature cows following the usual age of dam correction factors for weaning weight.
Feed intake often represents the single greatest cost of beef production (Ferrell & Jenkins, 1984). In this simulation, the feed intake of the cows was simulated as a continuous deterministic process occurring over the lifetime of each female, following Anderson et al. (1983), who modelled feed intake as a function of weight, weight change and milk production. Feed intake of the feedlot-fed progenies was derived from data collected on animals that participated in the centralised growth test (Phase C) of the NBCRIS.
The original females were first exposed for breeding at time ≈ 0.6 (that is, 0.6 years after they were weaned) and thus, provided they had conceived, expected to calve at two years. Success or failure to conceive was simulated as a stochastic event from the binomial distribution. In the simulation, it was assumed that females that conceive would also calve. An overall probability (p) of conception (and calving) can be approximated from the inter-calving period (ICP) reported in the NBCRIS from the equation:

or

This formula assumes that any simulated female will calve at least in alternate years, and thus may be a presumably slight overestimate of the true probability of conception if less fertile females are considered. Equivalently, the inter-calving period of the simulated females is constrained to the interval of 365 to 730 days.
Calf survival was simulated as a stochastic process, with mean level determined from the difference between the probability of conception and calving (as shown above) and the weaning rate. Weaning weight was simulated as having mean level equal to that observed in the NBCRIS.
At weaning, the calves were divided into two groups: replacement females and those entering the feedlot. The replacement female group represents an opportunity cost of production and was not considered further. Those entering the feedlot were fed to a breed-specific pseudo-optimal endpoint, established to avoid selling their carcasses at a discounted price, based on fat depth and/or weight.
The feedlot component of the production system was simulated as having two phases. The first phase was a 28-d adaptation period during which a ration of moderate energy was fed. This phase was followed by a longer second phase (127 d), during which a ration of greater energy density was fed, and feed conversion decreased. Data presented by Strydom et al. (2001) were used to benchmark the phenotypes simulated during the feedlot phase and at harvest.
Beef carcasses were the end-product of the simulated production system. Carcass weight and fat depth were simulated stochastically from a bi-variate normal distribution, given their respective deterministically simulated means and coefficients of variation and correlation taken from the literature. Carcasses were valued based on a series of discounts to the base price for carcass weight. Discounts were applied to carcasses that weigh less than 185 kg, carcasses that weigh more than 245 kg, carcasses with fat depth less than 1 mm, carcasses with fat depth between 5 and 10 mm, and carcasses with fat depth greater than 10 mm.
Breeding objectives
Three breeding objectives were derived to determine whether the objectives were consistent during the production, profitability and sustainability phases of agricultural development. These breeding objectives for the three phases were defined as income, profit and ratio of income to expense.
The genetic correlation (rA) between breeding objectives was calculated as follows (James, 1982):
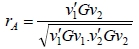
where vi is a vector of relative economic values for the ith breeding objective, and G is a variance-covariance matrix among traits in the breeding objective (Table 3).
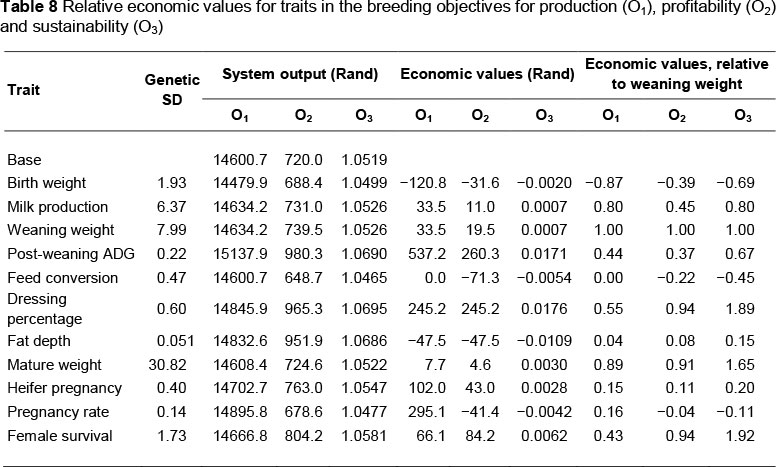
Income and expense (as described above) were discounted to time = 0, and accumulated for the entire simulation. Profit was then calculated as the difference between the accumulated discounted income and expense. Relative economic values that make up the breeding objective (that is, production, profitability and sustainability) were calculated by comparison of output (that is, income, profit or ratio of income to expenses) arising from a base simulation and a similar simulation in which the phenotype for one trait was perturbed by a single unit. Economic values were multiplied by their respective genetic standard deviation in order to provide an indication of their magnitude relative to the expected genetic variation. These values were then divided by the corresponding value for weaning weight to calculate the relative economic values.
Genetic trends were estimated from the EBVs generated by the ARC. The genetic trend was quantified by the regression of yearly mean EBV on generation number. The formula of Brinks et al. (1961) was used to calculate the generation number (gni) of an individual:
gni = 1 + (gns + gnd)/2
where gns and gnd are the generation numbers of the sire and dam, respectively. The base population was defined by recorded animals with unknown parents whose generation number was thus set to zero.
Results
Index-in-retrospect
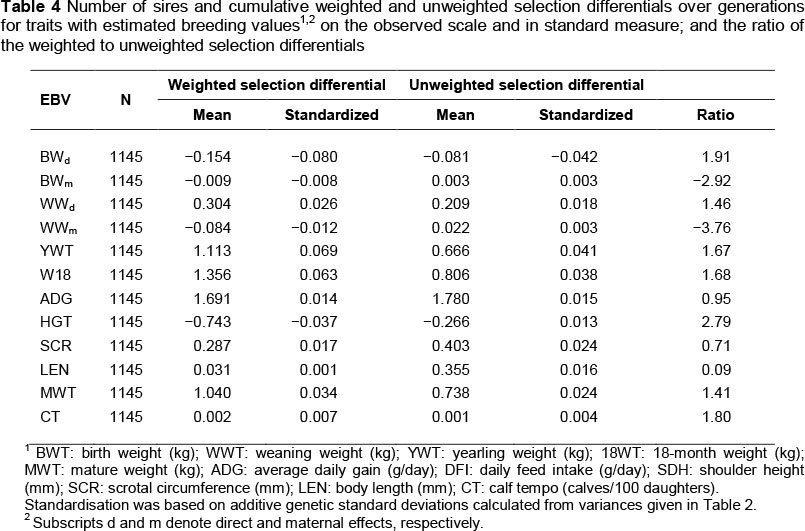
Numbers of sires and dams, and their weighted cumulated selection differentials, unweighted cumulated selection differentials, ratios of weighted to unweighted selection differentials, and standardized weighted and unweighted selection differentials are shown in Tables 4 and 5, respectively. It appears that relatively little directional selection has been applied for the performance traits with EBVs, because the standardized selection differentials are consistently much less than 1 standard deviation (except for calf tempo). Ignoring calf tempo because the EBV for it is not calculated for females, approximately 6.4 times more selection has been applied to males than to females for the traits with EBVs. However, the greater magnitude of the ratios of weighted to unweighted selection differential indicates a higher degree of natural selection affecting females than males.
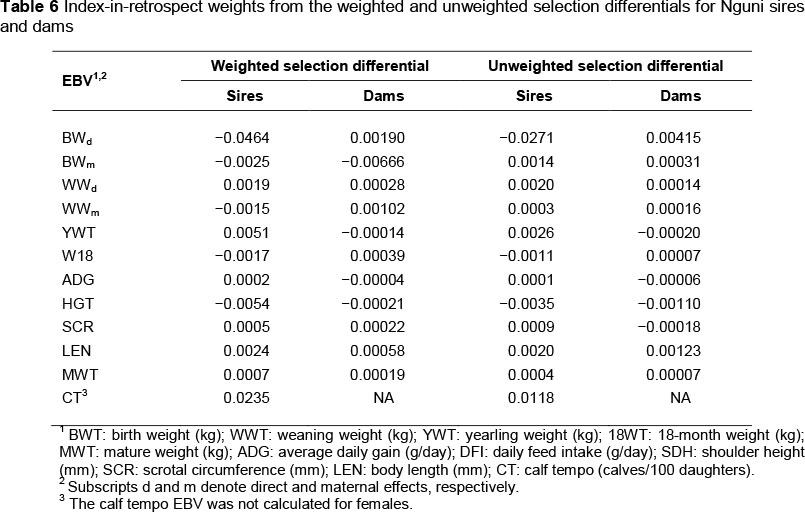
Results from the index-in-retrospect calculations using the weighted and unweighted selection differentials for the Nguni sires and dams are presented in Table 6. For sires, the indexes-in-retrospect weights derived from the weighted and unweighted selection differentials are quite similar (r = 0.96). However, for dams the indexes-in-retrospect derived from the weighted and unweighted selection differentials are somewhat different (r = 0.51). Correlations between sire and dam indexes-in-retrospect derived from weighted and unweighted selection differentials were 0.21 and 0.16, respectively.
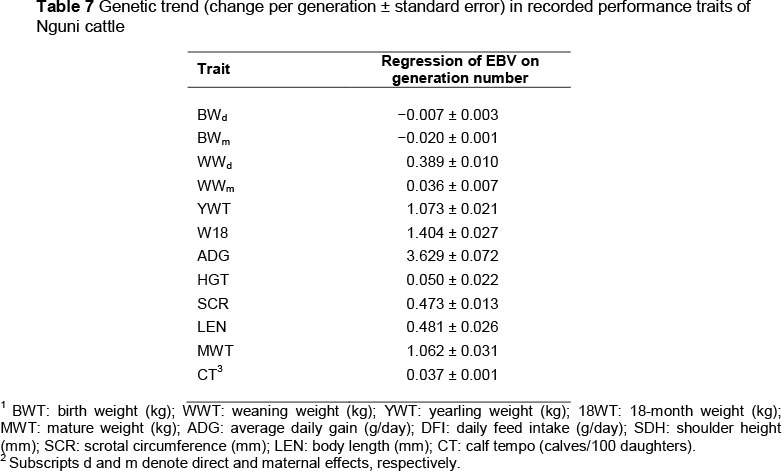
Genetic trends for recorded performance traits, calculated as the regression of yearly mean EBV on generation number, are presented in Table 7. From a biological perspective, the trends are consistently small across all traits.
Breeding objectives
Results from the base simulation and from simulations in which each trait was perturbed by a single unit are shown in Table 8. Results for the production phase are simulated incomes. Results for the profitability phase are simulated differences between income and expense. Results for the sustainability phase, reflecting production efficiency, are ratios of income over expense. All measures of income and expense were discounted to time 0; the time when the simulated weaned heifer calves entered the system.
Relative economic values were the product of the economic values and the genetic standard deviations for the traits. These products were rescaled such that the relative economic value of weaning weight (direct) was fixed at 1.0 across the three breeding objectives. For the ratio traits, feed conversion ratio reflects a change in the ratio with ADG held constant, and hence a perturbation of feed intake; and dressing percentage reflects a change in the ratio with live animal growth held constant, and hence a change in carcass weight. Correlations among breeding objectives were: 0.92 for the production and profitability objectives, 0.84 for the production and sustainability objectives, and 0.84 for the profitability and sustainability objectives.
Discussion
Selection
Overall, the cumulative selection differentials were small and had no clear direction. Mrode (1988) attributed the inability to achieve large differentials to unsoundness, selection on colour, death before production of any offspring, and failure to conceive as heifers. In an evaluation of a national recording scheme with many contributing breeders, the lack of a unified breeding objective would result in reduced selection differentials for the population. To the extent that there is any notable trend in the unweighted selection differentials, it would seem breeders favour larger males and smaller early maturing females.
Comparison of the weighted and unweighted selection differentials provides insight into joint effects of natural and deliberate or intended selection (Koch et al., 1994). Their ratio gives an indication of the effect of natural or deliberate selection on fertility of sires and dams in a population. Ratios greater than 1.0 indicate that animals with greater EBV have more progeny, with the magnitude of the ratio indicating the relative strength of natural versus imposed selection (Koch et al., 1994). Negative ratios indicate selection practices that are opposed to natural selection. For sires, ratios of weighted to unweighted selection differentials for most traits indicated some harmony between the selection imposed by the breeders and natural selection. Fluctuation of the ratios across traits is interpreted to reflect differing relative emphasis resulting from imposed and natural selection, but not conflicting goals with respect to the recorded EBV. For dams, breeder preference for small early maturing females seems opposed by natural selection. The weighted selection differentials, which take into account the number of progeny, were consistently small and positive, whereas the unweighted selection differentials were positive for early growth and negative for post-weaning traits, reflecting breeder preference. The positive unweighted selection differential for mature weight may reflect a lack of consistency in breeders' implementation of their selection objective. Similar to the weighted selection differentials for sires, the weighted selection differentials for dams were almost always positive.
Birth weight and weaning weight are both affected by direct effects (because of genes of the progeny) and maternal effects (because of genes of the dam). The selection differentials for maternal effects are consistently smaller than those for direct effects, despite the equality of units of expression. Because the weighted selection differentials for birth weight are consistently less than the unweighted values, natural selection appears to favour lighter birth weights. At a phenotypic level, increases in birth weight generally have been associated with increases in calving difficulty and reductions in calf survival (Bellows et al., 1971; Notter, 1978; Nelson & Beavers, 1982). Birth weight has been shown to be more strongly influenced by variation in direct than by maternal genetic effects (Koch et al., 1994). Greater direct effects on weaning weight were consistently favoured by selection in this study. However, the observation that sires whose daughters produced less milk seem favoured by natural selection, but dams that give more milk leave more progeny poses an interesting conflict. This conflict may, in part, be because of an antagonism of direct and maternal effects for pre-weaning growth (Koch et al., 1974; MacNeil et al., 1984).
Correlations among indexes
The observed sire and dam index-in-retrospect weights were close to zero, and generally smaller than those resulting from the selection experiment summarized by Koch et al. (1974). The present result implies that little consistent directional selection has been applied by Nguni breeders for the recorded traits. The correlations between the indexes derived from the weighted and unweighted selection differentials were positive for both the sires and dams. Thus, selection applied was similar, whether or not the selection differentials were weighted by the number of progeny produced. The moderate correlation among the indexes-in-retrospect for dams seemingly indicates somewhat different emphases in the goals of breeders and nature.
However, there was a notable lack of consistency in selection between the sire and dam for the same trait. As observed above, if index-in-retrospect weights derived from the unweighted selection differentials were positive for sires, then the corresponding index weights were negative for 73% of the EBV in the index-in-retrospect for dams. This causes the correlation between these indexes to be low. This result is interpreted to suggest breeders have conflicting goals for selecting bulls and heifers. These inconsistent goals compromise genetic improvement of the Nguni breed. Thus, there does not seem to have been any clear breeding objective employed by breeders that may facilitate the Nguni breed reaching its potential as a hardy dam line, as envisioned by Scholtz (1988) and MacNeil & Matjuda (2007), among others.
Genetic trends
Genetic trends of estimated breeding values for direct and maternal effects were very small, but in some instances were comparable with trends observed for the Afrikaner, another South African indigenous breed in the region (Beffa et al., 2009c). Data used here were from the field and any selection response across the data set would have resulted from a unified breeding objective being implemented by the breeders. The lack of substantial consistent genetic trends again highlights the lack of a consensus breeding objective for Nguni in South Africa.
Breeding objectives
The perspective taken here was that of a general purpose, self-replacing herd of Nguni cattle that are adapted to the harsh environment of southern Africa. The breed evolved through a subsistence phase during which directional selection pressure was not applied to performance traits. This seems to be supported by the index-in-retrospect calculations discussed above. However, in meeting national goals for alleviating poverty and reducing hunger (Department of Agriculture, Republic of South Africa, 2008) the formal definition of breeding objectives may be helpful in guiding future selection practices. The uniqueness of the South African beef production scenario means that there are commercial farms in the profitability phase, and there is on-going discussion of sustainable production practices, while emerging and communal farm that were established on redistributed land are largely in the production phase. In addition, a portion of breeders continue to advocate for breeding Nguni cattle under natural selection alone.
To achieve the breeding goals of the production, profitability and sustainability phases of development, continued selection pressure will be required to improve the performance attributes of Nguni cattle. This is achieved through balancing breeding values for growth, carcass and fertility, as indicated by the economic values for each trait (Table 8). In general, the breeding objective for the production phase places greatest emphasis on growth traits, namely direct and maternal (milk production) weaning weight, mature weight and carcass weight, as manifest through dressing percentage. Fertility and carcass traits received less emphasis. In the breeding objective for the profitability phase, the weaning, carcass and mature weights remained important, with female survival or longevity having increased emphasis. In the breeding objectives for both the production and profitability phases, direct effects on weaning weight were the single most important trait. However, the relative importance of direct effects on weaning weight was reduced relative to carcass weight and female survival in the breeding objective for the sustainability phase.
While successful reproduction constituted the single most important phenotypic driver of profitable and efficient beef production (Ponzoni & Newman, 1989; Melton, 1995), the pregnancy rates of heifers and cows received relatively little emphasis in the breeding objectives. In heifer pregnancy, this resulted from relatively few expressions (once per lifecycle per female) and low genetic variance. The number of expressions of pregnancy rate is greater for cows; however, its genetic variance is even less.
Robertson (1959) suggested that a genetic correlation above 0.8 for a trait measured in two environments indicated the lack of genotype by environment interaction. Here, this rule is adopted to evaluate the similarity of breeding objectives (traits) across production phases (environments). The correlations among the breeding objectives of the three phases that were estimated here indicate that they are geared for breeding in the same general direction. It should be borne in mind that modelling of the production system was done at farm level in the current study. It might be worthwhile in future to investigate the magnitude of the correlation between breeding objectives when modelling is done at levels higher than the farm. However, these breeding goals all imply the application of directional selection pressure, thus they are uncorrelated with the breeding goal under the subsistence phase, when by definition no directional selection pressure was applied for any of the recorded performance traits.
Conclusions
In the past, inconsistencies in selection by Nguni breeders had limited genetic improvement towards the often-stated goal of producing a hardy and fertile breed. Resulting genetic trends for recorded traits have been near zero and substantially less than those that are theoretically attainable. Breeding objectives focused on production, profitability and sustainability were highly correlated and, if implemented, are expected to create genetic trend in the same general direction. Granted, the emphasis placed on growth traits tended to diminish, and emphasis placed on fitness increased with progression through the phases. As Nguni breeders may be divided with regards to the appropriateness of a breeding objective focused on production, profitability or sustainability, substantial positive correlations between the objectives indicate opportunities for them to work together in the breeding of Nguni cattle.
References
Anderson, V.L., Jost, L., Dinkel, C.A. & Brown, M.A., 1983. Prediction of daily total digestible nutrient requirements of beef cows in northern climates. J. Anim. Sci. 57, 373-386. [ Links ]
Bawden, R.J., 1989. Towards action researching systems. In: Action Research for Change and Development. Ed: Zuber-Skeritt, O., Centre for the Advancement of Learning and Teaching, Griffith University, Brisbane, Australia. pp. 20-51. [ Links ]
Beffa, L.M., Van Wyk, J.B. & Erasmus, G.J., 2009. Long-term selection experiment with Afrikaner cattle. 3. Selection applied and selection response in calf growth traits. S. Afr. J. Anim. Sci. 39, 106-113. [ Links ]
Bellows, R.A., Short, R.E., Anderson, D.C., Knapp, B.W. & Pahnish, O.F., 1971. Cause and effect relationships associated with calving difficulty and calf birth weight. J. Anim. Sci. 33, 407-415. [ Links ]
Brinks, J.S., Clark, R.T. & Rice, F.J., 1961. Estimation of genetic trend in beef cattle. J. Anim. Sci. 20, 903. (Abstract). [ Links ]
Department of Agriculture - Republic of South Africa. 2008. National agricultural research and development strategy. [ Links ]
Dickerson, G.E., 1969. Experimental approaches in utilizing breed resources. Anim. Breed. Abstr. 37, 191-202. [ Links ]
Dickerson, G.E., Blunn, C.T., Chapman, A.B., Kottman, R.M., Krider, J.L., Warwick, E.J., & Whatley Jr, J.A., 1954. Evaluation of selection in developing inbred lines of swine. North Central Regional Pub. 38, Missouri Research Bulletin No. 551. [ Links ]
James, J.W., 1982. Construction, uses and problems of multitrait selection indices. Proceeding of the 2nd World Congress on Genetics Applied to Livestock Production. Madrid. [ Links ]
Koch, R.M., Gregory, K.E. & Cundiff, L.V., 1974. Selection in beef cattle. I. Selection applied and generation interval. J. Anim. Sci. 39, 449-458. [ Links ]
Koch, R.M., Cundiff, L.V. & Gregory, K.E., 1994. Cumulative Selection and genetic change for weaning or yearling weight or for yearling weight plus muscle score in Hereford cattle. J. Anim. Sci. 72, 864-885. [ Links ]
MacNeil, M.D., 2005a. Breedingth objectives for Angus cattle in South Africa and the United States, pp. 17-23. Paper presented at the 9th World Angus Forum Technology Meeting, Cape Town, South Africa. [ Links ]
MacNeil, M.D., 2005b. Breeding objectives for terminal sires for use in U.S. beef production systems. Proceedings of the Beef Improvement Federation 37th Annual Research Symposium and Annual Meeting. Billings, Montana USA. pp. 82-87. [ Links ]
MacNeil, M.D. & Matjuda, L.E., 2007. Breeding objectives for Angus and Charolais specialized sire lines for use in the emerging sector of South African beef production. S. Afr. J. Anim. Sci. 37, 1-10. [ Links ]
MacNeil, M.D. & Mott, T.B., 2000. Using genetic evaluations for growth and maternal gain from birth to weaning to predict energy requirements of Line 1 Hereford beef cows. J. Anim. Sci. 78, 2299-2304. [ Links ]
MacNeil, M.D., Cundiff, L.V., Dinkel, C.A. & Koch, R.M., 1984. Genetic correlations among sex-limited traits in beef cattle. J. Anim. Sci. 58, 1171-1179. [ Links ]
MacNeil, M.D., Nugent, R.A. & Snelling, W.M., 1997. Breeding for profit: An introduction to selection index concepts. Proceeding of the Range Beef Cow Symposium XV. Rapid City, South Dakota. [ Links ]
Magee, W.T., 1965. Estimating response to selection. J. Anim. Sci. 24, 242-247. [ Links ]
Melton, B.E., 1995. Conception to consumption: The economics of genetic improvement. Proceedings of the Beef Improvement Federation. Sheridan, WY. [ Links ]
Mrode, R.A., 1988. Selection experiments in beef cattle. Part 2: A review of responses and correlated responses. Anim. Breed. Abstr. 56, 155-167. [ Links ]
Nelson, L.A. & Beavers, G.D., 1982. Beef x beef and dairy x beef females mated to Angus and Charolais sires. I. Pregnancy rate, dystocia and birth weight. J. Anim. Sci. 54, 1138-1149. [ Links ]
Notter, D.R., Cundiff, L.V., Smith, G.M., Laster, D.B. & Gregory, K.E., 1978. Characterization of biological types of cattle. VI. Transmitted and maternal effects on birth and survival traits in progeny of young cows. J. Anim. Sci. 46, 892-907. [ Links ]
Ponzoni, R.W. & Newman, S., 1989. Developing breeding objectives for Australian beef cattle breeding. Anim. Prod. 49, 35-47. [ Links ]
Rege, J.E.O., 2001. Defining livestock breeds in the context of community based management of farm Animal Genetic Resources. In: Proc. Workshop on Community Based Management of Animal Genetic Resources. Mbabane, Swaziland. May 7-11. FAO Corporate Repository. [ Links ]
Robertson, A., 1959. The sampling variance of the genetic correlation coefficient. Biometrics 15, 469-485. [ Links ]
Scholtz, M.M., 1988. Selection possibilities of hardy beef breeds in Africa: The Nguni Example. In: Proceedings of the 3rd World Congress on Sheep and Beef Cattle Breeding. Paris, France. [ Links ]
South Africa Livestock Breeding. 2004. Picasso Headline Pty Ltd. Cape Town, South Africa. [ Links ]
Strydom, P.E., Naude, R., Smith, M.F., Kotze, A., Scholtz, M.M. & Van Wyk, J.B., 2001. Relationships between production and product traits in subpopulations of Bonsmara and Nguni cattle. S. Afr. J. Anim. Sci. 31, 181-194. [ Links ]
Van Vleck, L.D., 1993. Selection Index and Introduction to a Mixed Model Methods. CRC Press. Boca Raton, Florida, USA. [ Links ]
Received 3 May 2013
Accepted 8 May 2014
First published online 14 June 2014
# Corresponding author: norman@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}