










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.44 n.1 Pretoria Jan. 2014
SHORT COMMUNICATION
Variation in individual piglet birth weights in a Large White χ Landrace sow herd
T.J. ZindoveI; E.F. DzombaII; A.T. KanengoniIII; M. ChimonyoI
IDiscipline of Animal and Poultry, University of KwaZulu-Natal, Private Bag X01, Scottsville, Pietermaritzburg 3209, South Africa
IIDiscipline of Genetics, University of KwaZulu-Natal, Private Bag X01, Scottsville, Pietermaritzburg 3209, South Africa
IIIAgricultural Research Council, Animal Breeding and Genetics, Private Bag X2, Irene 0062, South Africa
ABSTRACT
Within-litter birth weight variation in pigs is increasingly becoming important in influencing the profitability of pig enterprises. The objective of the study was to characterize within-litter birth weight variation in piglets from Large White χ Landrace sows. The study was conducted using records from 1 768 litters collected between January 1998 and September 2010 from a pig herd in South Africa. The number of piglets born alive (NBA) ranged from three to 18. The mean within-litter birth weight coefficient of variation (CVBWT) was 17.6% and ranged from 0.47% to 50.7%. The distribution of CVBWT in the herd was positively skewed. CVBWT increased as NBA increased. Multiparous sows farrowed litters with higher CVBWT than gilts. To enhance profitability of pig enterprises, the selection for increased NBA should be followed by selection for decreased CVBWT.
Keywords: Coefficient of variation, litter weight at birth, number of piglets born alive, parity
Number of piglets born alive (NBA) has been a major component of sow productivity, and genetic improvement programmes linked to it have been given priority (Kim et al., 2005). The number of teats a sow has limits the number of piglets it can nurture to weaning. When a sow farrows more piglets in a litter than its number of teats, the excess piglets have to be fostered (Canario et al., 2010). To date, sows from most breeds exhibit NBA of over 12 (Umesiobi, 2009). Kim et al. (2005) reported average teat numbers of 14 in Large White and Landrace sows. Therefore, the success of continued selection for litter size is likely to be limited by the number of teats.
Since improvement for NBA is approaching its optimum level, the next challenge could be to maximize piglet survival and growth rates. Milligan et al. (2002) suggested that one reason for low survival rates and a reduced average litter weight at weaning in large litters is the unfavourable positive correlation between NBA and within-litter birth weight variation. Light piglets have a reduced likelihood of surviving to weaning age and have reduced growth rates and weights at slaughter compared with their litter mates (Wolf et al., 2007). Uniform piglets at birth are likely to produce uniform pigs at weaning and at marketing (Fix et al., 2010). In addition, production of uniform piglets at birth reduces the incidences of foster-mothering, which is commonly practised to reduce variation among litter mates. A study by Straw et al. (1998) has shown that cross-fostering is adequate to achieve high survival rates owing to its negative effects such as reduced growth rates. Large weight variation at birth also requires the use of more pens, thereby increasing the cost of housing and cleaning, and complicating management requirements. Within-litter birth weight variation, defined as the distribution of individual weights within a litter, has not been given much attention despite its recognized impact on pig enterprises (Wolf et al., 2007). Factors affecting the variability in piglet birth weights are complex and poorly understood. These factors include nutrition and the age of the sow, breed of boar and genetic selection of gilts (Chimonyo et al., 2006). To improve the homogeneity of litters at birth, the relative importance of these factors needs to be explored. The objective of this study was to characterize within-litter birth weight variation in pigs, and determine factors that influence birth weight variation. It was hypothesized that non-genetic factors influence within-litter birth weight variation.
Data were collected from a pig herd at the Agricultural Research Council (ARC) farm at Irene, South Africa. The farm is located at 25°34'0" S and 28°22'0" E and lies 1523 m above sea level. An approximate mean annual rainfall of 715 mm is received with mean annual temperature of 17.3 °C. The temperature in the hot humid season (November to January) averages 23 °C, while the cold dry season (May to July) has an average temperature of 15 °C.
Data used in the study included 20 741 piglets from 1 816 litter records produced from January 1998 until September 2010. The records consisted of piglet identity, parity number, farrowing date, farrowing month, farrowing year, NBA and individual piglet birth weights. From these records, mean birth weight (MBWT), litter weight at birth (LWT), within-litter birth weight coefficient of variation (CVBWT), minimum birth weight (MinBWT) and maximum birth weight (MaxBWT) were calculated. Records of litters with piglets fostered in or out were excluded from the analyses. Litters with fewer than three piglets were assumed to have piglets fostered out and were excluded from the analyses. Data from 48 litters were deleted in this way, leaving 1 768 litters available for analyses.
The distribution of birth weights within the litter was described by several quantities. The arithmetic mean represented the average birth weight. PROC UNIVARIATE (SAS, 2009) was used to examine the distribution of CVBWT and MBWT and frequency distributions. Skewness and kurtosis were derived to describe the deviation of the distribution of CVBWT and MBWT between litters from the (symmetric) normal distribution.
The effect of parity, month of farrowing, year of farrowing, their interactions and relevant covariates on NBA, LWT, MBWT, MinBWT, MaxBWT and CVBWT was determined using generalized linear model procedures for repeated measures (SAS, 2009) to allow for repeated records of the sow. The relationship between CVBWT and NBA was plotted using Proc Gplot (SAS, 2009).
Summary statistics of studied traits are shown in Table 1. A skewness value of zero indicates a normal distribution (Milligan et al., 2002). The negative skewness value for NBA indicated that the majority of the NBA values were above the litter mean. The CVBWT and MBWT were positively skewed.
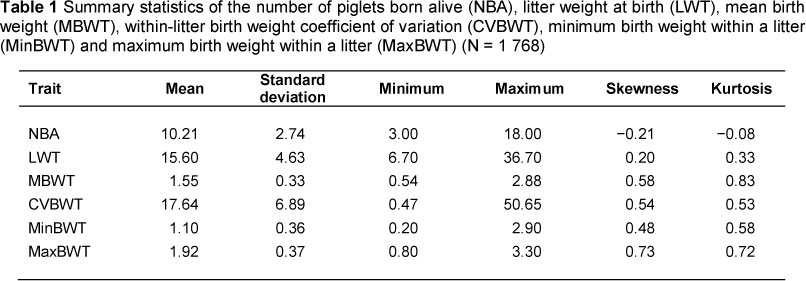
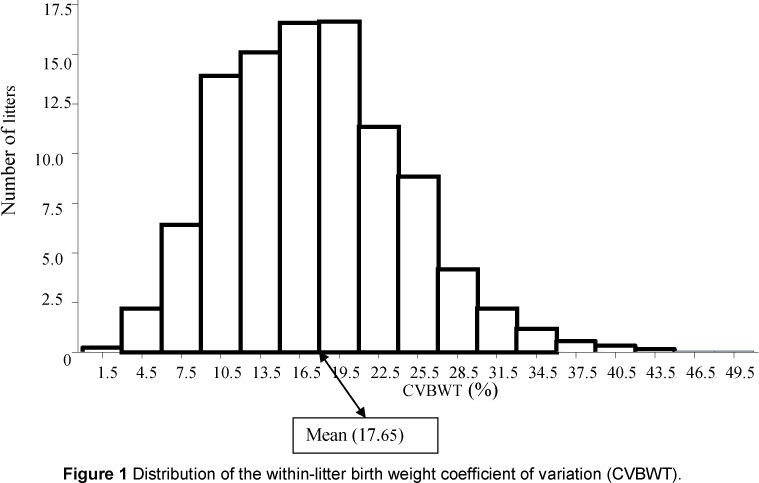
CVBWT ranged from 0.47% to 50.7%, reflecting considerable differences among litters over the 13-year period. The range of CVBWT reported in this study was larger than that reported by Wolf et al. (2008). This difference could be owing to variations in herd structures and the minimum NBA used for analysis. The CVBWT mean of 17.7% was on the lower end of the typical range of 18% to 25% reported by Le Dividich (1999). The finding that a considerable number of litters had CVBWT higher than the herd's mean CVBWT (Figure 1) implies that there is scope for improvement of within-litter birth weight homogeneity. The reported range and mean for CVBWT was higher than values reported by Wolf et al. (2008). This deviation could be ascribed to differences in environmental conditions and herd composition. The observed phenotypic increase in MBWT with year (Figure 2) shows that there was a remarkable improvement in MBWT. The reduction in NBA over the period under study could be owing to the negative correlation between NBA and MBWT. When breeders select for improved MBWT, NBA will be decreased. Year effects on MBWT and NBA can be the result of other factors such as a change in managerial strategies (e.g. differences in quality of feed and feeding regimens) by year and changes in parity distribution within the herd. Absence of year effects on CVBWT could be evidence that there was no change in within-litter variability in weights during the period under study. This could be because the trait was not being selected for over the period under study.
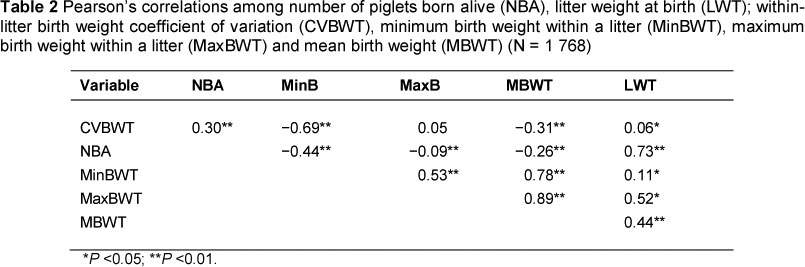
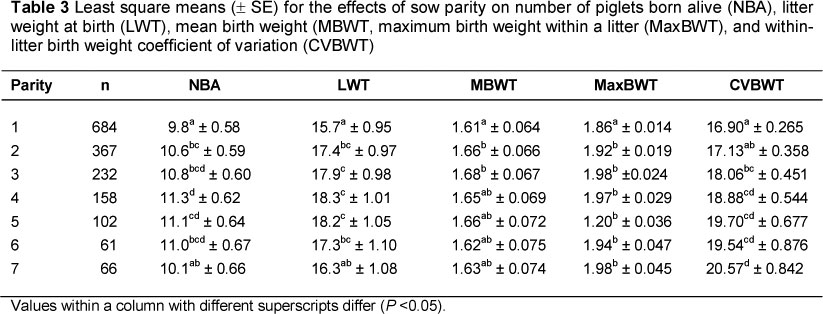
Table 2 shows that there is a positive correlation between NBA and CVBWT. This observation supports reports by Canario et al. (2010), who suggested that variation in birth weight of litter mates could be owing to differences in litter size and uterine capacity, size of the placenta and interactions among these factors. The NBA for gilts was lower (P <0.05) than that for multiparous sows, except for those greater than parity 6 (Table 3). This finding agrees with results of Fernandez et al. (2008).
Gilts and young sows have lower ovulation rates than mature sows (Cole et al., 1994) thus resulting in smaller litter sizes in primiparous sows (Table 3). The reduction in NBA in older sows (with parities greater than 6) can be attributed to high incidences of farrowing problems that lead to higher piglet mortalities, which reduce NBA. Piglets from primiparous sows were more uniform than piglets from older sows, this result being related to the effect of parity on NBA (Milligan et al., 2002). Contrary to the suggestion that the effect of parity on CVBWT is related to a parity effect on NBA, there was no significant difference in CVBWT of first and second parity sows in the present study, despite a significant difference in NBA between the two parities. This would suggest that litter heterogeneity is partly influenced by parity. The dissension can also be because of the exclusion of litters with fewer than three piglets from the analysis. This eliminated mainly first parity sows, since NBA increases with sow parity (Fernandez et al., 2008). Primiparous gilts and sows in their late parities had lower MBWT than sows in parities 2 to 5 (Table 3). This result agrees with findings by Milligan et al. (2002). Gilts produce piglets of low birth weights because they are still physiologically immature and hence have to partition nutrients between their own nutrient requirements and those of the foetuses (Campos et al., 2011). Age-associated physiological deterioration in sows in their late parities results in less efficient utilisation of feed to provide nutrition to foetuses resulting in low MBWT (Mungate et al., 1999).
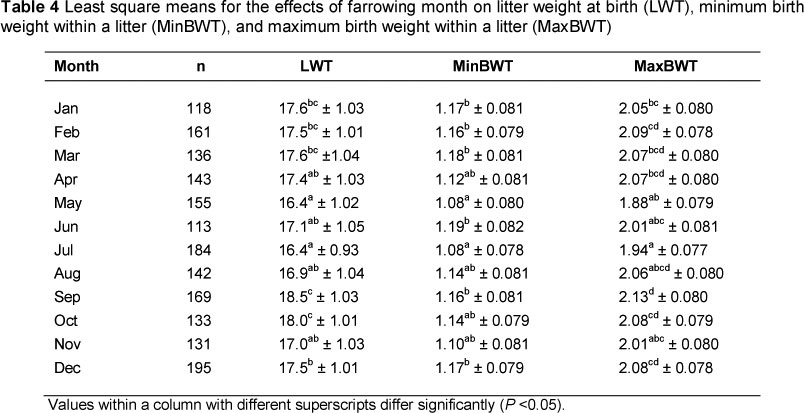
As shown in Table 4, the heaviest litters were born in September and October (P <0.05), while the lightest litters were recorded during the cool-dry months (May to August) (P <0.05). Seasonal variation in MaxBWT and MinBWT can be related to variation of LWT with season, since there is a strong positive correlation between LWT and both MaxBWT and MinBWT. These findings confirm results from other studies that indicate that sows produce not only smaller litters, but also lighter piglets during hot or warm seasons (Tummaruk & Khatiworavage, 2011).
In conclusion, large NBA was associated with an increased within-litter birth weight variation. Parity also influenced CVBWT, and within-litter birth weight variation was lowest in primiparous sows. Future research in selection programmes should attempt to reduce within-litter birth weight variation in large litters. However, CVBWT is a ratio, and its selection can be complicated owing to antagonistic correlated response in its component traits such as NBA and MBWT. Thus, when selecting for CVBWT, there is need to consider the economic values of selection responses from the component traits and possibly selection index procedures.
References
Canario, L., Lundgren, H., Haandlykken, M. & Rydhmer, L., 2010. Genetics of growth in piglets and the association with homogeneity of weight within litters. J. Anim. Sci. 88, 1240-1247. [ Links ]
Chimonyo, M., Dzama, K. & Bhebhe, E., 2006. Genetic determination of individual birth weight, litter weight and litter size in Mukota pigs. Livest. Sci. 105, 67-69. [ Links ]
Cole, D.J.A., Wiseman, J. & Variety, M.A., 1994. Principles of pig science. British cataloguing in publication data, Nottingham, UK. [ Links ]
Fernandez, A., Rodriganez, J., Zuzuarregui, J., Rodriguez, M.C. & Silio, L., 2008. Genetic parameters for litter size and weight at different parities in Iberian pigs. Span. J. Agric. Res. 6, 98-106. [ Links ]
Fix, J.S., Cassady, J.P., Holl, J.W., Herring, W.O., Culbertson, M.S. & See, M.T., 2010. Effect of piglet birth weight on survival and quality of commercial market swine. Livest. Sci. 123 (1-3), 98-106. [ Links ]
Kim, J.S., Jin, D.I., Lee, J.H., Son, D.S., Lee, S.H., Yi, Y.J. & Park, C.S., 2005. Effects of teat number on litter size in gilts. Anim. Reprod. Sci. 90, 111-116. [ Links ]
Milligan, B.N., Fraser, D. & Kramer, D.L., 2002. Within-litter birth weight variation in the domestic pig and its relation to pre-weaning survival, weight gain, and variation in weaning weights. Livest. Prod. Sci. 76, 181-191. [ Links ]
Mungate, F., Dzama, K., Mandisonza, K. & Shoniwa, A., 1999. Some non-genetic factors affecting commercial pig production in Zimbabwe. S. Afr. J. Anim. Sci. 29, 164-173. [ Links ]
Straw, B.E., Burgi, E.J., Dewey, C.E. & Duran, C.O., 1998. Effects of extensive crossfostering on performance of pigs on a farm. J. Am. Vet. Med. Assoc. 212, 855-856. [ Links ]
Tummaruk, P. & Khatiworavage, C., 2011. Seasonal effect on the litter size at birth in sows differed among parity groups. Proc. 5th Asian Pig Veterinary Society Congress 7-9 March, Pattaya, Thailand. [ Links ]
Umesiobi, D.O., 2009. Vitamin E supplementation to sows and effects on fertility rate and subsequent body development of their weanling piglets. J. Agric. Rural Dev. Trop. 110, 155-168. [ Links ]
Wolf, J., Zakova, E. & Groeneveld, E., 2008. Within-litter variation of birth weight in hyper prolific Czech Large White sows and its relation to litter size traits, stillborn piglets and losses until weaning. Livest. Sci. 115, 195-205. [ Links ]
 Correspondence:
Correspondence:
M. Chimonyo
chimonyo@ukzn.ac.za
Received 18 January 2012
Accepted 14 February 2014
First published online 23 March 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}