










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.44 n.1 Pretoria Jan. 2014
SHORT COMMUNICATION
The Tankwa Karoo National Park feral goat population: A unique genetic resource
A. KotzéI, II; J.P. GroblerII; E. van Marle-KösterIII; T. JonkerIV; D.L. DaltonI, II
INational Zoological Gardens of South Africa, P.O. Box 754, Pretoria 0001, South Africa
IIDepartment of Genetics, University of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa
IIIDepartment of Animal & Wildlife Sciences, University of Pretoria, Private Bag X20, Hatfield, Pretoria 0028, South Africa
IVNorthern Cape Department of Agriculture, Land Reform and Rural Development, Private Bag X5018, Kimberley 8300, South Africa
ABSTRACT
The feral goats from Tankwa Karoo National Park in the Northern Cape, South Africa, constitute a potentially unique goat population, which dates back to the early 1900s, but is now at risk of extinction. A total of 66 feral goats from Tankwa Park and former Tankwa goats, now kept on a private farm were genotyped, using eight microsatellite markers. The data were compared with genotypic data of selected commercial breeds (Angora, Boer and Saanen dairy goats). Analysis of population structure using Bayesian and frequency-based methods suggests some uniqueness in the Tankwa populations. This uniqueness may reflect decades of random drift, but could also reflect alleles for adaptation to a harsh environment resulting from natural selection. These results are the first for the Tankwa goat and provide essential information for compiling a strategy for conservation and breeding of this genetic resource.
Keywords: Conservation, genetic variability, goat biodiversity, Tankwa feral goats
South Africa has a rich variety of farm animal genetic resources, used at different levels of commercial production. These local animals often include indigenous types with low production potential, which are nevertheless well adapted to the different climatic regions of South Africa compared with imported European breeds (Pieters, 2007; Van Marle-Köster et al., 2008). Several studies have been conducted to genetically characterize South African indigenous farm animal breeds and local types (Buduram, 2004; Kotze et al., 2004; Pieters, 2007; Van Marle-Koster et al., 2008; Soma et al., 2012).
There are a number of examples worldwide where domesticated animals have turned wild and adapted to and survived in harsh climatic and environmental conditions. Examples include the wild horses of the Namib desert (Cothran et al., 2001), feral cattle on Chirikof Island (McNeil et al., 2007) and feral goats in numerous localities (Campbell & Donlan, 2005). A similar situation was highlighted in South Africa, when South African National Parks (SANParks) initiated the removal of feral goats from Tankwa Karoo National Park. The Northern Cape Department of Agriculture, Land Reform & Rural Development (NC-DALRRD) realized that these animals might represent unique genetic material, which has been exposed to intense natural selection over the years. The interest by the department was backed by its commitment to the goat industry in the Northern Cape and the Commercialisation of Goats Programme, which then spawned the Tankwa Goat conservation framework.
It is estimated that the goats at Tankwa have been free roaming for more than 50 years, with a population size that varied between 100 and 300. There is a high degree of variation in the colour and coat appearance in the Tankwa goat, as indicated in Figure 1. The coat tends to be longer than the typical South African Boer goat and other indigenous types, and the primary coat colours are black, red, white and grey with mixtures of spotted, dappled and tri-colour variations (Jonker personal observation). Preliminary measurements taken on captured goats by NC-DALRRD indicate a respiratory type with long legs and long horns, especially in males. Although the collation of information on the phenotypic characteristics is ongoing, departmental officials have confirmed that Tankwa feral goats are well adapted to surviving under harsh grazing conditions, are parasite resilient, have good mothering abilities, and survive high predation pressures (Jonker personal observation). The aim of this study was to investigate the possible genetic uniqueness of the Tankwa feral goats in support of the conservation framework.
Twenty-six Tankwa goats captured from Tankwa Karoo National Park were included in this study (designated Tan-TP). Samples were also taken from 40 Tankwa goats captured in another area of the Roggeveld mountains and relocated to a private farm near Calvinia approximately 20 years ago (designated former Tankwa or Tan-F). Blood (5 mL) was collected from each animal and stored at -20 °C until DNA extraction was performed. As representative of other goat breeds, 40 blood samples from Saanen dairy goats were collected from a private owner in the Middelpos region in the Northern Cape. Furthermore, genotypic data generated for Boer and Angora goat populations at the Department of Animal and Wildlife Sciences, University of Pretoria, were included for comparison.
DNA was extracted from blood samples with the Qiagen© DNA isolation kit, following the manufacturer's protocol. Eight microsatellite loci were selected from the ISAG and FAO recommended lists for genetic diversity studies, namely ILStS087, MCM527, SRCRSP5, SRCRSP8, BM1329, SRCRSP24, SRCRSP23 and OarFcb20. Amplification was carried out in a 15 µL final volume containing 1 x PCR buffer, 1.5 - 2.5 mM MgCl2, 200 µM of each dNTP, 10 pmol of each of the forward and reverse primer, 1 unit (U) Promega GoTaq® DNA polymerase (Promega Corporation, Madison, WI, USA) and 50 ng genomic DNA template. PCR conditions were 5 min at 95 °C, 30 cycles for 30 sec at 95 °C, 30 sec at 50 - 65 °C and 30 sec at 72 °C, followed by 72 °C for 20 min. PCR products were run against GenescanTM 500 LIZTM internal size standard on an ABI 3130 genetic analyzer (Applied Biosystems, Inc., Foster City, CA). Samples were genotyped with GeneMapper v. 4.0 (Applied Biosystems).
Genetic diversity was estimated as unbiased heterozygosity (Hz; Nei, 1987), observed heterozygosity (Ho) and the mean number of alleles per locus (A), using MSTOOLKIT (Park, 2001). To account for unequal sample sizes, genetic diversity was calculated as allelic richness (RS) using FSTAT version 2.9.3.2 software (Goudet, 2001). Conformation of genotypes to expected Hardy-Weinberg equilibrium (HWE) was calculated using ARLEQUIN version 3.1 software (Excoffier et al., 2005).
The genetic relationships between goat populations were inferred via Baysian cluster analysis using STRUCTURE version 2.33 (Pritchard et al., 2000). A model was used that assumes admixture, but with uncorrelated allele frequencies. STRUCTURE was run for five replicates from K = 1 - 8, with a run-length of 200 000 MCMC repetitions, following a burn-in period of 100 000 iterations. The AK statistic of Evanno et al. (2005) was then calculated to further investigate results from structure, using STRUCTURE Harvester software (Earl et al., 2011). Genetic differentiation among populations was quantified using Fst (Wright, 1965), as implemented in ARLEQUIN software. This software was also used to implement a hierarchical investigation of the overall distribution of genetic diversity, through an analysis of molecular variance (AMOVA).
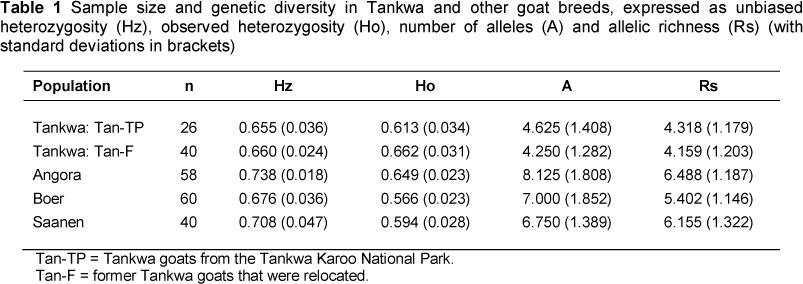
Results of molecular and statistical analysis showed that all eight loci were polymorphic, with the number of alleles per marker varying from 7 to 15 over all the populations. No unique alleles were observed in the Tankwa populations. The levels of genetic diversity in the two Tankwa goat populations were similar and comparable with the values found in other breeds, but were on the lower end of the range (Table 1). For example, Hz values in the Tan-TP and Tan-F populations were 0.655 and 0.660 compared with values of 0.709 to 0.738 in the other goat breeds. Similarly, Rs values (a measure adjusted for sample size) were 4.318 and 4.159 in the Tankwa animals, compared with 5.402 to 6.488 in other breeds. Diversity values for the Angora breed were the highest in all instances. The diversity values should be seen in the correct context, however, since the values for the Tankwa populations are based on individuals from relatively small and isolated populations, whereas values for the other breeds are representative of the breeds, rather than individual populations. For example, in a study on a single population of Angora feral goats from Faure Island, an average of 3.9 alleles was reported (Balasingham et al., 1999), compared with 8.125 for pooled Angora animals in the current study. Values for the individual Tankwa populations can thus be seen to be comparatively high.
The number of loci that deviated significantly (P <0.05) from HWE ranged from 1 to 5 per population. The Tankwa populations showed the fewest deviations (n = 2 in Tan-TP; n = 1 in Tan-F), with the highest number of deviations observed in the Angora breed (n = 5) and slightly lower numbers (n = 4) in the Boer and Saanen breeds. There was no pattern of deviations from HWE to suggest that any specific locus consistently deviates from HWE in goats, with the exception of the locus SRCRSP5, where significant deviations were observed in 4/5 groups. Although the Tankwa Tan-TP and Tan-F populations superficially have fewer deviations from HWE than the established goat breeds, the Tankwa are actual populations, whereas the other breeds are represented by grouped samples. Grouped samples are unlikely to conform to the conditions for maintenance of HWE, and deviations may not be representative of the retention of HWE in individual populations. Nevertheless, the Tankwa goats have been subjected to natural selection, which would contribute to the maintenance of HWE, compared with artificial selection in commercial breeds.
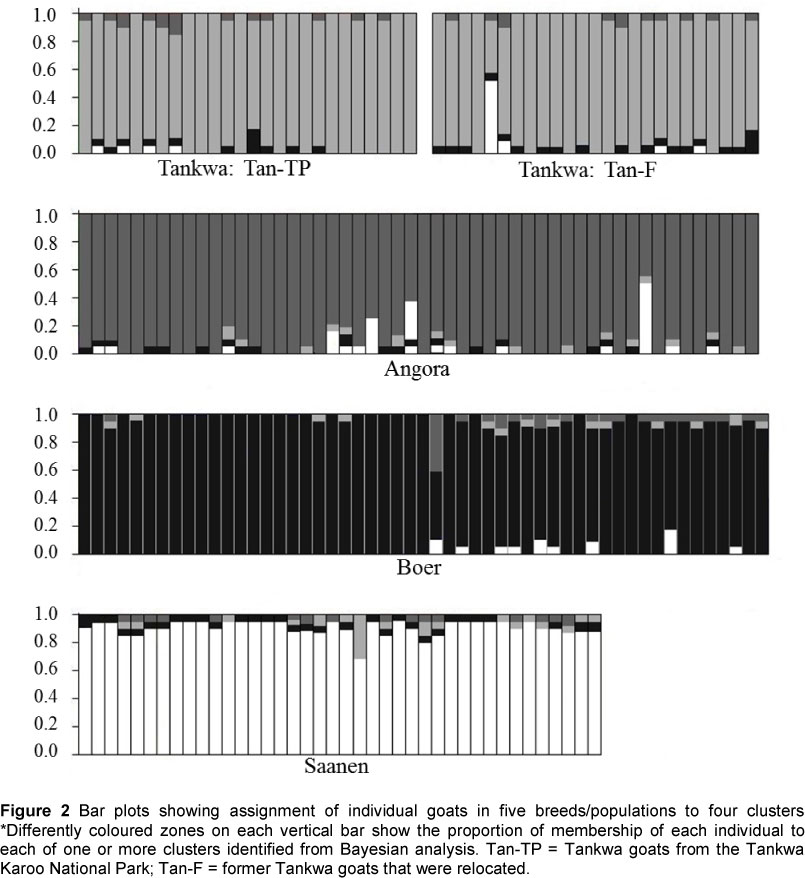
The Bayesian and frequency-based measures of population structure and differentiation suggested considerable uniqueness in the Tankwa breed. First, results from STRUCTURE showed strong support for a true population structure, consisting of four clusters, with the ΔK approach of Evanno et al. (2006) indicating a value of K = 4 as the most likely outcome. Also, the mean of -Ln(K) values for K = 4 occurred at a point where -Ln(Pr) values plateau, and with little deviation among replicate runs. The proportion of membership of each individual to the four clusters is shown in Figure 2. At this K value, the Tan-TP and Tan-F animals grouped together, with the Angora, Boer and Saanen dairy goats breeds each contained in distinct clusters. There is an indication of a shared genetic background or introgression from Boer and Saanen breeds in a limited number of individual Tankwa animals. In the former Tankwa goats that are now being farmed as a commercial population, possible introgression from Boer and Saanen individuals was found, which is plausible. For the goats from Tankwa Karoo National Park, introgression from Boer and Saanen goats was also observed. This was expected in the Boer goat, if the region where these Tankwa goats occur is considered. Introgression from Saanen dairy goats is less easily explained and the apparent introgression of Saanen goat genetic material into the Tankwa populations may indicate ancient shared ancestry rather than recent events. Indeed, the Angora and Boer breeds show a signature of introgression from the Saanen dairy goat breed, supporting the hypothesis of a lack of recent admixture.
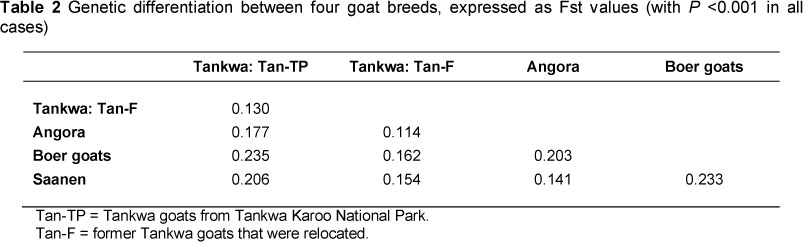
Results from Fst support the hypothesis of some uniqueness in the Tankwa populations, with a lower Fst value between the two Tankwa populations compared with the differentiation between these populations and most of the remaining breeds (Table 2). The Fst value between goats from the two nominally Tankwa populations was 0.130, compared with larger values of 0.177 to 0.235 between Tankwa Park goats and representative Angora, Boer goat and Saanen dairy goat populations.
An AMOVA with all breeds/populations treated as separate groups suggested considerable variation among breeds, with 19.7% of variation attributed to differences among breeds and 80.3% to within-breed diversity. Because almost 20% of overall variation in these goats resides between breeds, this suggests that there may be a significant reservoir of diversity for directed selection of goat breeds.
Bayesian and frequency-based statistical approaches to population structure analysis suggest some uniqueness in the Tankwa breed. A significant proportion of this assumed uniqueness no doubt reflects simple drift resulting from decades of isolation. However, there may be an adaptive component to the uniqueness, resulting from an extended period of natural selection to the particular environmental conditions. Since the Tankwa has not been studied from a production perspective, it is not known what specific production advantages the breed may have. However, because these animals are highly adapted to a life without human intervention in a harsh environment, this suggests that the breed may offer exciting possibilities for commercialization.
This study was the first attempt to provide genetic information on the feral Tankwa goat from South Africa. Results suggest a relatively high variability and detectable genetic differentiation compared with other breeds. These results serve as baseline information on the Tankwa goat and may guide further investigations into the conservation and breeding of these animals to ensure not only survival, but also utilization.
Acknowledgements
We would like to thank the staff of Veterinary Services and Research from the Northern Cape Department of Agriculture, in particular Deon Kriel for providing samples, and Anri van Wyk from the National Zoo for technical assistance and DNA profiling.
References
Balasingham, T.G., Robinson, N.A. & McGregor, B.A., 1999. Implications for the conservation of genetic diversity in mohair goats from a comparison of a relic island population with breeds farmed in Australia. Aust. J. Exp. Agric. 39, 411-418. [ Links ]
Bourdon, R.M., 2000. Understanding Animal Breeding. Prentice Hall Inc., New Jersey. [ Links ]
Buduram, P., 2004. Genetic characterization of Southern African sheep breeds using DNA markers. Unpublished M.Sc. dissertation, University of the Free State, South Africa. [ Links ]
Campbell, K. & Donlan, C.J., 2005. Feral goat eradications on islands. Conserv. Biol. 19, 1362-1374. [ Links ]
Cothran, E.G., Van Dyk, E. & Van der Merwe, F.J., 2001. Genetic variation in the feral horses of the Namib Desert, Namibia. J. S. Afr. Vet. Assoc. 72, 18-22. [ Links ]
Earl, D.A. & Von Holdt, B.M., 2012. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Con. Gen. Res. 4, 359-361. [ Links ]
Evanno, G., Regnaut, S. & Goudet, J., 2005. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14, 2611-2620. [ Links ]
Excoffier, L., Laval, G. & Schneider, S., 2005. Arlequin, version 3.0: An Integrated Software Package for Population Genetics Data Analysis. Computational and Molecular Population Genetics Laboratory (CMPG), Institute of Zoology, University of Berne, Switzerland. [ Links ]
Goudet, J., 2001. FSTAT, a Program to Estimate and Test Gene Diversities and Fixation Indices. Version 293. Institut de Zoologie et d'Ecologie Animale, Universite de Lausanne, Lucerne, Switzerland. [ Links ]
Groeneveld, L.F., Lenstra, J.A., Eding, H., Toro, M.A., Scherf, B., Pilling, D., Negrini, R., Finlay, E.K., Jianlin, H., Groeneveld, E. & Weigend, S.: GLOBALDIV Consortium, 2010. Genetic diversity in farm animals: A review. Anim. Genet. 1, 6-31. [ Links ]
Kotze, A., Swart, H., Grobler, J.P. & Nemaangani, A., 2004. A genetic profile of the Kalahari Red goat breed from southern Africa. S. Afr. J. Anim. Sci. 34 (Suppl. 1), 10-12. [ Links ]
McNeil, M.D., Cronin, M.A., Blackburn, H.D., Richards, C.M., Lockwood, D.R. & Alexander, L.J., 2007.
Genetic relationships between feral cattle from Chirikof Island, Alaska and other breeds. Anim. Gen. 38, 193-197.
Nei, M., 1987. Molecular Evolutionary Genetics. Columbia University Press, New York. 512 p. [ Links ]
Park, S.D.E., 2001. Trypanotolerance in West African cattle and the population genetic effects of selection. Unpublished PhD. thesis, Dublin, University of Dublin, Ireland. [ Links ]
Pieters, A., 2007. Genetic characterization of commercial goat populations in South Africa. Unpublished MSc. dissertation, University of Pretoria, South Africa. [ Links ]
Pritchard, J.K., Stephens, M. & Donnelly, P., 2000. Inference of population structure using multilocus genotype data. Genetics 155, 945-959. [ Links ]
Rosenberg, N., Burke, T., Elo, K., Feldman, M.W., Freidlin, P.J., Groenen, M.A.M., Hillel, J., Maki-Tanila, A., Tixier-Boichard, M., Vignal, A., Wimmers, K. & Weigend, S., 2001. Empirical evaluation of genetic clustering methods using multilocus genotypes from 20 Chicken breeds. Genetics 159, 699-713. [ Links ]
Soma, P., Kotze, A., Grobler, J.P. & Van Wyk, J.P., 2012. South African sheep breeds: population genetic structure and conservation implications. Small Rumin. Res. 103, 112-119. [ Links ]
Van Marle-Köster, E., Hefer, C.A., Nel, L.H. & Groenen, M.A.M., 2008. Genetic diversity and population structure of locally adapted South African chicken lines: Implications for conservation. S. Afr. J. Anim. Sci. 38, 271-281. [ Links ]
Wright, S., 1965. The interpretation of population structure by F-statistics with special regard to systems of mating. Evolution 19, 395-420. [ Links ]
 Correspondence:
Correspondence:
A. Kotzé
antoinette@nzg.ac.za
Received 23 April 2013
Accepted 9 January 2014
First published online 13 February 2014


{kind=link}
{kind=link}
{kind=link}
{kind=link}