










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.43 supl.1 Pretoria Jul. 2013
Effects of dietary energy content and provision of β-adrenergic agonist on the production of feedlot lambs
T.S. BrandI, II, #; M.P. GenisII; L.C. HoffmanII; W.F.J. van de VyverII; C. van der WaltI; G.F. JordaanII
IElsenburg Institute for Animal Production, Depart. of Agriculture: Western Cape, Private Bag X1, Elsenburg, 7607
IIDepartment of Animal Sciences, Stellenbosch University, Private Bag X1, Matieland, 7602
ABSTRACT
Recent increases in mutton lamb prices have resulted in many lamb producers opting to finish more and more lambs on the farms in a feedlot system rather than marketing directly from the field. The aim of this trial was to determine the effect of dietary energy, as well as the inclusion of a β-adrenergic agonist, on the production of feedlot lambs. South African Mutton Merino lambs (108) of different sexes were weaned (± 36.4 kg) at ca. 120 days of age and were randomly divided into six experimental groups (18 lambs per group), which were adapted to each treatment. Lambs were housed in individual pens. The treatments consisted of three diets (diet 1: 11.3 MJ ME/kg food, diet: 2 12.0 MJ ME/kg food and diet 3: 12.7 MJ ME/kg) with either the inclusion or absence of a ß-adrenergic agonist at 8.5 g/ton. The experimental design was a 3 x 2 x 2 factorial design with dietary energy level, the provision of a ß-adrenergic agonist and sex as main factors. No interaction occurred between treatments and data are presented as the effect of dietary energy level and the inclusion of a stimulant on the intake, growth and feed conversion ratio (FCR). Neither the dietary energy content nor the β-adrenergic agonist had an effect on FCR. Neither one of the main factors had a significant effect on feed intake. The sex of the lambs had a significant effect on the average daily gain of the lambs with the ewes growing faster than the rams.
Keywords: South African mutton merino, energy, β-agonist.
Introduction
A feedlot is defined as a production unit where livestock are divided into different paddocks and fattened up just before they are slaughtered (Smith, 2011). The recent increase in mutton and lamb prices have resulted in many lamb producers opting to finish weaner lambs in a feedlot system rather than marketing directly from the field. Another aspect that occurred with the increase in meat prices was that sheep abattoirs have become more vertically integrated and are buying in young weaned lambs and finishing them off in their own feedlots - which are often adjacent to the abattoir. Sheep feedlotting is a relatively new industry in South Africa. Therefore very little information exists on the dietary requirements of the feedlot sheep.
Lambs are growing in importance for animal production (Sayed, 2011; Smith, 2011). Therefore it is important to design the feedlot rations to minimize the number of days on feed in feedlots and to maximize the growth rate (Notter et al., 1991). The level of the lamb's nutrition is very important for proper development and growth. It is important to formulate an intensive ration for the feedlot environment to benefit from the high rate of development of lamb in their early stages of growth (Sayed, 2011). Energy is one of the most important dietary ingredients for the lamb to utilize efficiently so as to improve the gain and productivity of the animal (Duddy, 2007; Sayed, 2011). The regulation of intake plays a central role in the performance of livestock. Although the general assumption is that the energy level of the lambs' diet determines their intake, there are other factors contributing to intake, such as different pasture types and their maturity, feed types and supplements (Poppi, 2008). Variation in intake accounts for most of the difference in live weight gain (Poppi, 2008).
Beta -adrenergic agonists (β-AA) are repartitioning agents that have been found to enhance protein accretion in cattle (Lopez-Carlos et al., 2010), though very little research has been conducted on their use in sheep. The use of β-AA (an anabolic agent) is another method, other than nutrition and selection, to enhance the growth efficiency of livestock (Fiems, 1987). The β-AA is administrated orally through the diet. Commonly β-AA increases the average daily gain (ADG) and improves feed efficiency (Mersmann, 2002; Montgomery et al., 2009; Eckermann et al., 2011), decreases adipose tissue and increases skeletal muscle (Mersmann, 1998; Lopez-Carlos et al., 2010). With the use of a β-AA the muscle weight can be increased up to 40% (Mersmann, 1998), although the weight increase can vary from muscle to muscle (Beermann, 2002). Avendano-Reyse et al. (2006) found that the use of the β-AA, Zilpaterol hydrochloride (ZH), increases the dressing percentage. The aim of the study was therefore to determine the effect of different dietary energy levels, with either the inclusion or absence of a ß-agonist in the diet, on the production parameters of weaned ram and ewe lambs.
Material and Methods
Lambs used in this trial were obtained from Langgewens Experimental Farm, Western Cape, South Africa. South African Mutton Merino (SAMM) lambs (108) of both sexes were weaned (± 36.4 kg) at ca. 120 days of age. These lambs were transported to Elsenburg Experimental Farm, Western Cape, South Africa, where they were vaccinated against pulpy kidney. The lambs were housed in pens (2 lambs/pen; the size of the pens are within the norm described by animal welfare guidelines; 117 cm x 177 cm).
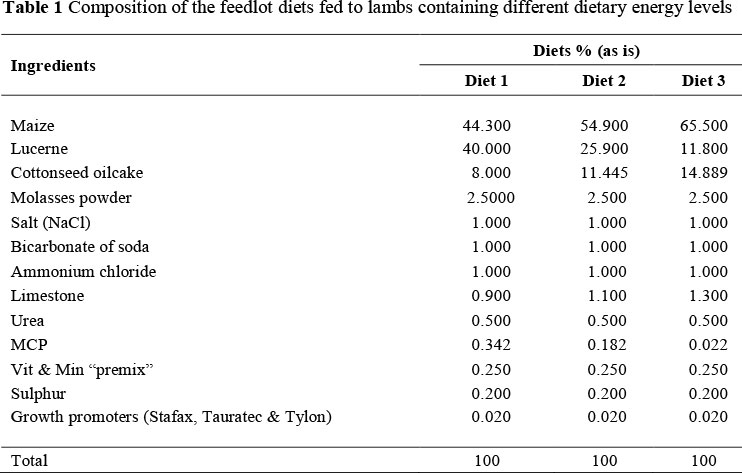
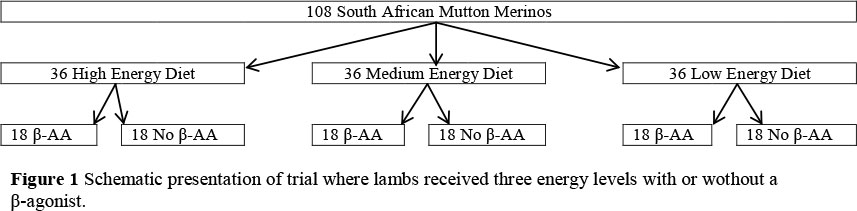
Lambs were randomly divided into six groups (18 lambs/group) and allocated to three diets (Table 1). Diet 1 (11.3 MJ ME/kg DM on an "as is" basis), diet 2 (12.0 MJ ME/kg DM on an "as is" basis) and diet 3 (12.7 MJ ME/kg DM on an "as is" basis) which included either the inclusion of a β-adrenergic agonist (β-AA) at 8.5 g/ton or not (Figure 1). The inclusion levels of the β-AA were recommended by Intervet (the company who supplies this β-AA to the beef industry). The lambs were fed for a period of approximately 6 weeks (40 days) in the feedlot.
Fresh, clean water and feed were provided ad libitum to the lambs. Lambs and feed refusals were weighed weekly. The lambs were sheared before they were slaughtered after approximately six weeks in the feedlot. Lambs were weighed prior to transport to the abattoir from the Elsenburg Experimental Farm; the lambs were slaughtered the next day. The lambs were slaughtered at a weight (± 54 kg) higher than the average commercial weight (± 40 kg) for this breed so as to determine the effect of the β-AA. A registered sheep abattoir (Roelcor, Malmesbury, Western Cape, South Africa) was used to slaughter the lambs, using standard South African techniques. The aim of this trial was to determine the effect of dietary energy as well as the inclusion of a ß-AA on the production of SAMM feedlot lambs.
The experiment consisted of a completely randomixed design with six treatmens (dietary energy level with the inlusion of a β-AA or not). The treatment design was a 3 x 2 x 2 factorial with dietary energy level (low, medium and high), the provision of a β-AA (either included or absent) and sex (rams and ewes) as the main factors.
A factorial analysis of variances was performed on the data using SAS for windows version 9.1.3 Proc GLM (SAS, 2000), whereas normality was tested with the shapiro-Wilk test (Shapiro & Wilk, 1965).
Results and Discussion
No interaction occurred between the treatments and the data are thus presented as the effect of dietary energy, inclusion of the stimulant and sex on intake, growth and FCR. The different dietary energy levels in the diets had no effect (P >0.05) on any of the production characteristics (Table 2).
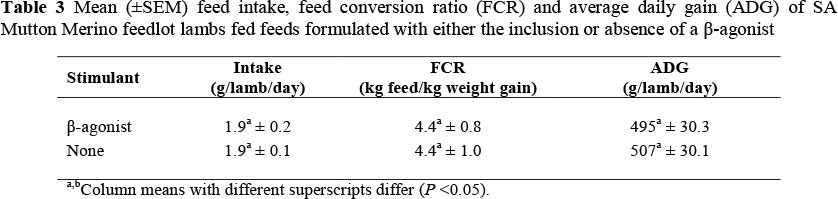
Feeding a β-agonist to livestock typically increases the ADG (average daily gain) and improves feed efficiency (Beckett et al., 2009; Lopez-Carlos et al., 2010). Neither the inclusion of the β-agonist nor the absence of the β-agonist in the diet had any effect on any of the production characteristics (feed intake, FCR or ADG) (Table 3).
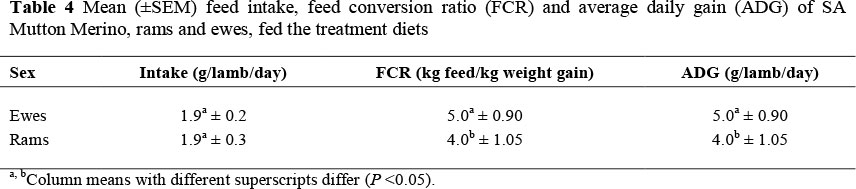
The sex of lambs is known to influence a number of production factors such as the growth rate, body composition, FCR and the meat quality (Rodriguez et al., 2008). The use of ram lambs, as meat producing animals, satisfies the consumer trend for increasing leanness (Notter et al., 1991). These ram lambs will typically be slaughtered at a younger age than their wether and ewe counterparts produced on forage-based diets. Seideman et al. (1984) found that the intact ram lamb is a more desirable meat producing animal, as it grows faster, utilizes feed better and produces a heavier carcass with less fat and leaner red meat than castrates (wethers) and ewes. In this investigation, sex affected the FCR and ADG (Table 4). Although sex had no effect on feed intake, the ram lambs showed a better (P <0.05) FCR than the ewe lambs. The ADG of the ram (459 g/lamb/day) lambs were almost 100 g/lamb/day higher (P <0.05) than that of the ewe (363 g/lamb/day) lambs.
Conclusion
There were no differences in the effect of the energy levels on the production parameters of the lambs. This may be due to the fact that the feed intakes were the same for the three energy levels, as well as for both sexes. According to the limited literature, the β-agonist should increase and improve both the ADG and the FCR of the feedlot lamb as it does for cattle. In this study no difference was found between the inclusion and the absence of the β-agonist in the diet; it could be that the ß-agonist was included at too low a concentration in the diet, or because the amount of dietary nutrients were limited. On the other hand, sex significantly affected the production parameters with the intact rams performing more efficiently than the ewes.
References
Avendano-Reyse, L., Torres-Rodriguez, V., Meraz-Murillo, F.J., Perez-Linares, C., Figueroa-Saavedra, F. & Robinson, P.H., 2006. Effect of two ß-adrenergic agonists on finishing performance, carcass characteristics, and meat quality of feedlot steers. J. Anim. Sci. 84, 3259-3265. [ Links ]
Beckett, J.L., Delmore, R.J., Duff, G.C., Yates, D.A., Allen, D.M., Lawrence, T.E. & Elam, N., 2009. Effects of zilpaterol hydrochloride on growth rates, feed conversion, and carcass traits in calf-fed Holstein steer. J. Anim. Sci. 887, 4092-4100. [ Links ]
Beermann, D.H., 2002. Beta-adrenergic receptor agonist modulation of skeletal muscle growth. J. Anim. Sci. 80, 18-23. [ Links ]
Duddy, G., Bell, A., Shands, C. & Hegarty, R., 2007. Feedlotting lambs. PrimeFacts, Profitable & Sustainable Primary Industries. Available: http://www.dpi.sw.gov.au [2012/28/08]. [ Links ]
Eckermann, S.E., Lardy, G.P., Thompson, M.M., Van Emon, M.L., Neville, B.W., Berg, P.T. & Schauer, C.S., 2011. Effects of graded levels of zeranol implants on feedlot performance, carcass characteristics, and incidence of prolapse and mortality in lambs. Sheep Research Report, North Dakota State University. pp. 7-12. [ Links ]
Fiems, L.O., 1987. Effect of beta-adrenergic agonists in animal production and their mode of action. Annals Zootechnol. 36, 271-290. [ Links ]
Lopez-Carlos, M.A., Ramirez, R.G., Aguilera-Soto, J.I., Arechiga, C.G., Mendez-Llorente, F., Rodriguez, H. & Silva, J.M., 2010. Effect of ractopamine hydrochloride and zilpaterol hydrochloride on growth, diet digestibility, intake and carcass characteristics of feedlot lambs. Livest. Sci. 131, 23-30. [ Links ]
Mersmann, J.J., 1998. Overview of the effects of ß-adrenergic receptor agonists on animal growth including mechanism of action. J. Anim. Sci. 76, 160-172. [ Links ]
Mersmann, J.J., 2002. Beta-adrenergic receptor modulation of adipocyte metabolism and growth. J. Anim. Sci. 80, 24-29. [ Links ]
Montgomery, J.L., Krehbiel, C.R., Cranston, J.J., Yates, D.A., Hutcheson, J.P., Nichols, W.T., Streeter, M.N., Bechtol, T.T., Johnson, E., TerHune, T. & Montgomery, T.H., 2009. Dietary zilpaterol hydrochloride. I. Feedlot performance and carcass traits of steers and heifers. J. Anim. Sci. 87, 1374-1383. [ Links ]
Notter, D.R., Kelly, R.F. & McClaugherty, F.S., 1991. Effects of ewe breed and management system on efficiency of lamb production: II Lamb growth, survival and carcass characteristics. J. Anim. Sci. 69, 22-33. [ Links ]
Poppi, D.P., 2008. The dilemma in models of intake regulation: mechanistic or empirical mathematical modelling in animal nutrition. In: Mathematical Modelling in Animal Nutrition. CAB International, Wallingford, UK. [ Links ]
Rodriguez, A.B., Landa, R., Bodas, R., Prieto, N., Mantecon, A.R. & Giraldez, F.J., 2008. Carcass and meat quality of Assaf milk fed lambs: Effect of rearing system and sex. Meat Sci. 80, 225-230. [ Links ]
SAS, 2000. SAS/STAT Users Guide, Version 8, First Edition, Volume 2. SAS Inst. Inc., Cary, North Carolina, USA. [ Links ]
Sayed, A.B.N., 2011. Effect of different energy levels of diets on the performance. Nutrient digestibilitiy and carcass characteristics of lambs. IJAVMS. 5, 472-476. [ Links ]
Shapiro, S.S. & Wilk, M.B., 1965. An analysis of variance test for normality (complete samples). Biometrika 52, 591-611. [ Links ]
Smith, O., 2011. Feedlot activity plan. Babil lamb feedlot. USAID, Iraq, Hawks of Iraq Company. [ Links ]
Wood, J.D., Enser, M., Fisher, A.V., Nute, G.R., Richardson, R.J. & Sheard, P.R., 2007. Manipulating meat quality and composition. Proc. Nutr. Soc. 58, 363-370. [ Links ]
Received 3 September 2012
Accepted 11 October2013
First published online 26 October 2013
# Corresponding author: tersb@elsenburg.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}