










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.42 no.4 Pretoria Jan. 2012
Effect of RRR-α-tocopherol succinate on the meat quality and antioxidative status in broilers
Xuhui ZhangI, II; Gaiqin WangI; Yanmin ZhouI; Tian WangI, #
ICollege of Animal Science & Technology, Nanjing Agricultural University, Nanjing 210095, P.R. China
IICollege of Forest Resource & Environment, Nanjing Forestry University, Nanjing 210037, P.R. China
ABSTRACT
The objective of this study was to compare the effects of two esters of α-tocopherol, all-rac-α-tocopherol acetate (DL-a-TOA) and RRR-a-tocopherol succinate (D-α-TOS), on meat quality and the antioxidative status in chicks. A total of 320 day-old Arbor Acres broiler chicks were randomly allocated to 4 treatments, each consisting of 8 pens of 10 chicks per pen. Birds in the control group received the basal diet supplemented with 30 mg DL-a-TOA/kg diet. In the other treatments the diet was supplemented with D-a-TOS at 15 mg/kg, 30 mg/kg or 60 mg/kg (TOS1, TOS2 and TOS3 treatments), respectively. The trial lasted 42 days. Positive correlations existed between dietary D-a-TOS levels and plasma and hepatic α-tocopherol concentrations, and a negative correlation with malonaldehyde (MDA) concentrations. In comparison with the control group, 30 mg/kg and 60 mg/kg of dietary D-a-TOS supplementation resulted in an increase in glutathione peroxidase (GSH-Px) activity and glutathione (GSH) content of the breast and thigh muscle and total superoxide dismutase (T-SOD) activity in the thigh muscle. Furthermore, the muscle MDA and hepatic reactive oxygen species (ROS) levels were reduced. As for meat quality, 48 h drip loss and shear force of breast and leg muscle were lower in broilers in the TOS1, TOS2 treatments and also the cooking loss in leg muscle. The study suggests that 30 mg/kg to 60 mg D-a-TOS/kg of the diet could enhance the antioxidant capacity of broiler meat, and its water-holding capacity and tenderness, in association with the reduction in lipid peroxidation as measured as a decrease in MDA and ROS concentrations.
Keywords: Meat quality, lipid peroxidation, antioxidative capacity, broiler chicks
Introduction
Lipid peroxidation in meat during post mortem ageing has been associated with a deterioration in the flavour, colour, odour, quality and nutritive value of the meat (Luciano et al., 2009; Karami et al., 2011). It has been reported that the oxidative stability of muscle tissue depends on its anti-/pro-oxidant balance (Cardenia et al., 2011), and therefore dietary supplementation with antioxidants might increase the oxidative stability of meat lipids (Eder et al., 2005; Guo et al., 2006a; Ao et al., 2011). Although synthetic antioxidants, such as butylated hydroxytoluene and butyl hydroxy anisol, have been used extensively in the meat industry, consumers' concerns about the safety and toxicity of these products have initiated a search for natural sources of antioxidants (Nuala et al., 2006).
Vitamin E is a major chain-breaking antioxidant involved in preventing the development of free radicals in cell membranes in vivo and post mortem (Dufrasne et al., 2000; Onibi et al., 2000; Sahin et al., 2002; Boler et al., 2009). Since vitamin E cannot be synthesized by the animal body, its presence in body tissues such as adipose and muscle is a reflection of dietary availability (Jensen et al., 1998). Published research has shown that dietary vitamin E delays the onset of lipid oxidation in pork and poultry meat (Asghar et al., 1991; Monahan et al., 1994; Guo et al., 2006b), and the feeding of high levels of vitamin E (300 - 700 mg/kg) resulted in reduced lipid oxidation, as measured by thiobarbituric acid reactive substances (TBARs) (Jensen et al., 1997; Corino et al., 1999). Furthermore, it has been reported that supranutritional supplementation of vitamin E to finishing livestock can improve the colour and lipid stability through accumulation in muscles. The beneficial effect of dietary vitamin E supplementation by enhancing the stability of lipids in muscle foods has been reported extensively for poultry, beef cattle, veal calves and pigs (Gray et al., 1996; Jensen et al., 1998).
It is general practice to include vitamin E in the form of all-rac-a-tocopherol acetate (DL-α-TOA) in poultry feeds (Villaverde et al., 2008) to maximize the oxidative stability of the meat. However, the main limitation in vitamin E treatment is acute supplementation because marked lipophilicity hampers tissue distribution and therefore cellular bioavailability. Tocopherol esters may have advantages for processing, storage and absorption because of their greater stability and water solubility compared with tocopherol alone (Neuzil et al., 2007). They are usually rapidly converted to the natural forms by intestinal or epidermal esterases in the gastrointestinal tract, and thus can be considered to be pro-vitamins, ultimately performing the same functions in the body as natural a-tocopherol (Zingg, 2007).
RRR-α-tocopherol succinate (D-α-TOS) is a new hydrophilic, synthetic analogue of vitamin E with special properties. In addition, the succinate ester of RRR-α-tocopherol is the most stable form at room temperature (Jensen et al., 1997). Early work demonstrated that as a RRR-a-tocopherol analogue, D-a-TOS not only possesses the physiological active functions of D-a-tocopherol, but it also has distinct medicinal value through immunomodulation and cytoprotection with its hydrophilic properties (Fariss et al., 2001; Zhang et al., 2001). Interestingly, it was also found that D-α-TOS, but not D-a-tocopherol and DL-α-TOA, appears to be far more effective in many aspects, for example in protecting isolated hepatocytes against many oxidative challenges (Fariss et al., 1985; 1990; 1997).
More recently, the use of D-a-TOS in humans has been recommended because of its unique anti-tumour property in the vitamin E family (Brigelius-Flohé et al., 2002), and it is now receiving increased attention. Our previous investigations confirmed that D-a-TOS has an immunoenhancement effect (Zhang et al., 2009) and regulates inflammatory immune responses (Zhang et al., 2010). In light of these findings, the purpose of this study was to compare the effect of supplementing diets at 30 mg/kg with a conventional vitamin E derivative, synthetic (all-rac-) α-TOA and 15, 30 or 60 mg/kg of natural (RRR-) a-TOS, on meat quality and antioxidative status in broiler chicks.
Materials and Methods
All animals were treated humanely, as outlined in the Guide for the Care and Use of Experimental Animals (Nanjing Agricultural University, Animal Care and Use Committee). A total of 320 day-old Arbor Acres broilers, obtained from a commercial hatchery (Hewei, Anhui, China), were randomly allocated to 4 treatment groups consisting of 8 replicates of 10 birds each per treatment. The average initial body weight did not differ among the four groups. The chickens were fed a maize-soybean basal diet. Birds in the control group received the basal diet supplemented with 30 mg/kg of DL-α-TOA (control). The experimental treatments were the basal diet with D-α-TOS supplementation at 15 mg/kg (TOS1 treatment), 30 mg/kg (TOS2 treatment) or 60 mg/kg (TOS3 treatment) for 42 d.
According to the actual determination of the dietary ingredients, the basal starter and grower diets contained 7.85 and 9.23 mg a-tocopherol/kg, respectively, and the concentrations of a-tocopherol in the experimental diets were 33.2, 20.2, 34.2 and 63.2 mg/kg in the starter phase, and 35.2, 21.9, 36.7 and 65.0 mg/kg in the grower phase, respectively. The percentage of all other major ingredients remained the same across treatments. The diets were formulated according to the NRC (1994) to meet the nutrient requirements of the broiler. The birds were fed a starter diet until 21 d of age, followed by a grower diet from days 21 to 42 (Table 1). All birds were placed in 32 wire cages (200 cm x 100 cm x 80 cm, length x width x height) of 10 chickens (0.20 m2 per chick) in a 3-level battery and housed in a room maintained at a brooding temperature of 34 °C to 35 °C for 5 d. The environmental temperature then was gradually reduced by 1 °C every two days until a final temperature of 22 °C was reached. The interior of the broiler house was naturally ventilated. The light regimen was a 12-h light-dark cycle (06:00 - 18:00 light). Birds were allowed to consume both feed and water ad libitum. Fresh diets were prepared once a week and were stored in sealed bags at 4 °C.
D-α-TOS used in the experiment was provided by Spring Fruit Biological Products Co., Ltd. (Jiangsu, P R. China), had a purity of 99.9%. DL-α-TOA was provided by the Huamu Institute of Animal Science and Technology (Nanjing, P. R. China), with a purity of 50%.
To determine the concentration of a-tocopherol in the diets, 5 g feed were saponified with ethanol and KOH (50%) and extracted with diethylic-petrol ether (2 : 1) (Zaspel & Csallany, 1983).
Body weights (BW) of each chick and food consumption of each group were recorded weekly, starting at 1 day of age, and weights were recorded to the nearest 0.01 kg. Growth performance was evaluated in terms of live body weight gain (BWG), food intake (FI), and feed/gain ratio (F/G). At 21 and 42 days one bird per replicate was randomly selected and weighed after feed deprivation for 12 h. Individual blood samples were taken and serum separated by centrifugation at 3500 rpm for 15 min and at 4 °C. Serum samples were frozen at -20 °C for further analysis. After collection of blood samples, all birds were killed by exsanguination and necropsied immediately. After decapitation, livers were excised, frozen in liquid nitrogen and stored at -20 °C for a-tocopherol analysis. After that, the breast and thigh muscles taken from the left side were divided into two parts. The fore part (50 g) was used to measure pH, drip loss, cooking losses, colour and shear force; the hind part was frozen at -20 °C and used to measure malondialdehyde (MDA) concentration according to the method of Zhu et al. (2007). Hepatic muscle tissues were weighed and homogenized using an Ultra-Turrax homogenizer (Tekmar Co., Cincinnati, OH) with the whole protein extraction kit (KeyGEN, Nanjing, P. R. China) on ice and then centrifuged at 2000xg for 20 min at 4 °C. The supernatant was collected and stored at -80 °C until analysed. Its protein concentration was determined using the Bradford method.
Vitamin E was analysed as described by Kayden et al. (1973) with modifications. Briefly, samples were saponified by mixing WLH (500 µL) with a 2% pyrogallol solution (5 mL) and heated for 2 min in a 70 °C shaking water bath. The tubes were removed, and 0.25 mL of 11 N KOH was added. The tubes were heated again in a shaking 70 °C water bath for 30 min, then placed in an ice bath. Two millilitres of hexane (used to extract the vitamin E) and 0.5 mL of water were added to the saponified samples and shaken vigorously for 2 min. One millilitre of the hexane layer was transferred to a 4 mL glass test tube for analysis. Standards of 1, 2, 4, 6, 8, and 10 µg/mL of a-tocopherol were prepared at the same time. A 0.2% bathophenanthroline solution (200 µL) was added to all the samples and standards and thoroughly mixed. Two hundred microlitres of 1 mM FeCl3 were added and samples were vortexed. After 1 min, 200 µL of an H3PO4 solution was added and vortexed again. The tubes were read on a spectrophotometer at 534 nm. The standard curve was used to calculate the concentration of a-tocopherol in each sample. The concentrations of a-tocopherol were expressed per gram of liver.
The fatty acids present in the diets were separated by gas chromatography (CGC 6850 Series Gas Chromatography System, Agilent Technologies, Santa Clara, CA), using a 60-m-long, 0.25-mm-inner diameter, and 0.25µm-film methylpolysiloxane capillary column (DB-23, 50% cyanopropyl, Agilent). The running time for each sample was approximately 60 min. The gases used, were nitrogen (carrier gas), hydrogen, helium and synthetic air. The operating conditions of the chromatograph were as follows: column flux, 1.00 mL/min; linear velocity, 24 cm/s; detector temperature, 280 °C; injector temperature, 250 °C; oven temperature, 110 °C for 5 min, 110 to 215 °C (5 °C/min), 215 °C for 24 min; volume of carrier gas injected, 1.0 µL. A cleaning procedure for the chromatograph was performed at intervals between samples using isopropanol and petroleum ether. The fatty acids were quantified by interpolation of an external pattern curve (Table 1) of the fatty acid profiles found in the diets.
The isolation of fresh hepatocytes was carried out in accordance with the method developed by Kim & Takemura (2003) and modified by Zhou et al. (2006). Briefly, the fresh liver was carefully excised and transferred onto a glass petri dish, and rinsed twice with phosphate buffered saline (PBS: 136.9 mM NaCl; 5.4 mM KCl; 0.81 mM MgSO4; 0.44 mM KH2PO4; 0.33 mM Na2HPO4; 5.0 mM NHCO3, pH 7.6) without Ca2+. The liver was dissected into small pieces and the tissue was digested for 20 min at room temperature with PBS containing 0.1% collagenase (Sigma, St. Louis, USA) on a shaker. The softened liver tissue was agitated and filtered through 70-µιη nylon mesh. The resulting cell suspension was transferred to a sterilized centrifuge tube and then centrifuged three times at 90xg for 3 min in the buffer containing 1.5 mM CaCl2 at 10 °C. After the last wash, the cell pellet was re-suspended in DMEM/F12 medium (Sigma, St. Louis, USA). Cells were counted using a haemocytometer (Reichert, Buffalo, NY, USA), and those with more than 90% viability, the trypan blue exclusion method were used for the following experiment.
Accumulation of intracellular ROS was assayed using 2',7'-dichlorofluorescein deacetate (DCFH-DA), which is de-esterified to 2',7'-dichlorodihydrofluorescein (DCFH2) by cellular esterases, and then DCFH2 is oxidized by ROS to form fluorescent 2',7'-dichlorofluorescein (DCF) (LeBel et al., 1992). An increase in fluorescence intensity was used to quantify the generation of net intracellular ROS. Briefly, 500 µL hepatocytes suspension (1χ107 cells/mL) were incubated in PBS buffer with 10 µmol/L DCFH-DA in the dark for 1 h at 25 °C and the fluorescence intensity determined using a fluorescence microplate reader (Infinite M200, TECAN, Switzerland) with excitation at 488 nm and emission at 525 nm (Roos et al., 2009). All samples were assayed in triplicate.
The concentrations of MDA, a marker index of lipid peroxidation product, were determined by measuring the TBA reactive substances (TBARS), using a corresponding diagnostic kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, P.R. China) with the methodology in the TBA method, as described by Placer et al. (1966). The principle is that TBA reacts with MDA to form a stable pink colour that is measured spectrophotometrically (OD at 532 nm). MDA values were expressed as nmol/mL serum and nmol/mg protein.
Muscle samples were analysed for total superoxide dismutase (T-SOD) activity with a T-SOD Assay Kit (Institute of Biological Engineering of Nanjing Jianchen, Nanjing, China). The methodology used in the kit is the nitrite method described by Oyanagui (1984). The method is based on the fact that SOD inhibits the generation of nitrite from oxidation of hydroxylamine by superoxide anion (O2-) that is produced by the xanthine-xanthine oxidase system. Glutathione peroxidase (GSH-Px) activity was determined with a GSH-Px Assay kit (Institute of Biological Engineering of Nanjing Jianchen). The methodology used in the kit is the dithio-nitro benzene method described by Hafeman (1974). Glutathione peroxidase is an enzyme that catalyzes glutathione oxidation by oxidizing the reduced tripeptide glutathione (GSH) into oxidized glutathione. The GSH content was determined spectrophotometrically at 420 nm using 5,5'-dithiobis-2-nitobenzoic acid. Both TBARS and GSH were determined using a corresponding assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, P.R. China) according to the instructions of the manufacturer.
The ultimate pH values of the muscles were measured 45 min and 24 h post mortem, using a portable pH meter (HI9023, Hanna Instruments, Padova, Italy) equipped with an insertion glass electrode (FC 230B, Hanna Instruments). Before measurement, the pH electrode was calibrated, using three buffers with pH values of 4.01, 7.00, and 9.01. The samples were always measured at the same place. The average pH value was obtained from three repeated measurements on the same muscle samples (Wang et al., 2009).
At 24 h and 48 h post mortem, a 25 mm cork borer was used to remove a cylindrical core from each sample (n = 48) at a right angle to the muscle fibre direction (Young et al., 2004). Each core was weighed and then attached to a fish hook that was secured to the lid of an airtight container. Each sample was then placed in the airtight container (Gladware small bowl altered with fish hook, Glad, Danbury CT) to avoid evaporation. Sealed cores were stored at 4 - 6 °C for 28 h and reweighed. Drip loss was calculated as a percentage: ((initial wt - final wt)/initial wt) x 100.
The muscles samples were refrigerated overnight at 4 °C and then brought to room temperature before cooking. Samples of the muscle of each chick were weighed (F), vacuum packed in plastic bags and cooked to an internal temperature of 70 °C by immersion in a digital thermostat water bath (HH-4, Jiangbo instrument, Jiangsu, China) (Ramirez et al., 2004). End-point internal temperature was monitored with a thermometer. Cooked samples were cooled under running water for 30 min. The samples were then removed from the bags, blotted and weighed (C). Cooking losses were calculated as (F - C) x 100/F.
After measuring cooking loss, the cooked muscle was used for the shear force assay, according to Meek et al. (2000). Slices of 1 cm x 1 cm were cut perpendicular to the fibre orientation of the muscle. Ten 1 cm χ 1 cm cores about 3 cm thick were removed parallel to the fibre orientation through the thickest portion of the cooked muscle. Warner-Bratzler shear force was determined by an Instron Universal Mechanical Machine (Instron model 4411, Instron Corp., Canton, MA). A Warner-Bratzler apparatus was attached to a 50-kg load cell, and tests were performed at a cross head speed of 200 mm/min. Signals were processed with the Instron Series ninth software package.
Meat colour was measured 45 min post mortem with a chroma meter (CR-410, Minolta Co. Ltd, Suita-shi, Osaka, Japan) to measure CIE LAB values (L* measures relative lightness, a* relative redness, and b* relative yellowness). A reading was made from the surface of the sample, representing the whole surface of the muscle. A white tile (L* 92.30, a* 0.32, and b* 0.33) was used as standard.
The effects of dietary treatment were determined by the student's t-test or one-way analysis of variance (ANOVA, SAS 9.0), followed, where appropriate, by Tukey's comparison test. Percentage data and data that were identified as nonhomogeneous (Bartlett's test) were subjected to arcsine transformation before analysis. Differences among treatments were separated using polynomial orthogonal contrasts to determine linear, quadratic and cubic effects of D-a-TOS supplementation. Significant differences were accepted if P <0.05.
Results
Table 2 shows that no significant differences (P >0.05) occurred in body weight gain (BWG), (FI) and feed/gain ratio (F/G) among the treatments in the periods of 0 -21 d, 22 - 42 d and the overall period of 1 - 42 days.
Plasma a-tocopherol concentrations were enhanced 1.0- and 2.52-fold in groups TOS2 (P <0.05) and TOS3 (P <0.01), respectively, compared with group TOS1, while the hepatic a-tocopherol concentrations for group TOS3 were elevated 1.53- (P <0.05) and 0.79-fold (P <0.05), compared with groups TOS1 and TOS2, respectively (Figure 1). Furthermore, the hepatic MDA concentrations for broilers in group TOS3 were 21.2% (P = 0.001), 25.8% (P = 0.001), and 14.4% (P = 0.020) lower than concentrations in the control, TOS1 and TOS2 groups, respectively (Figure 2). Plasma MDA concentrations exerted a tendency to be decreased in the TOS3 group (P <0.01) compared with the control group. Significant positive correlations were recorded between supplemental D-a-TOS levels and a-tocopherol concentrations in plasma (R2 = 0.9940, P = 0.0089) and the liver (R2 = 0.9478, P = 0.0043), and showed a negative correlation with plasma (R2 = -0.9937, P = 0.0096) and hepatic (R2 = -0.9780, P = 0.0438) MDA concentrations.
Hepatic ROS levels in groups TOS2 and TOS3 decreased (P <0.05) by 5.71% (P = 0.027) and 16.2% (P = 0.001), respectively, compared with the control, and decreased by 7.1% (P = 0.008) and 17.4% (P = 0.001) respectively, compared with the TOS1 group at 42 d. Significant negative correlation existed between dietary supplemental D-a-TOS levels and hepatic (R2 = -0.9965, P = 0.0103) ROS levels (Figure 3).
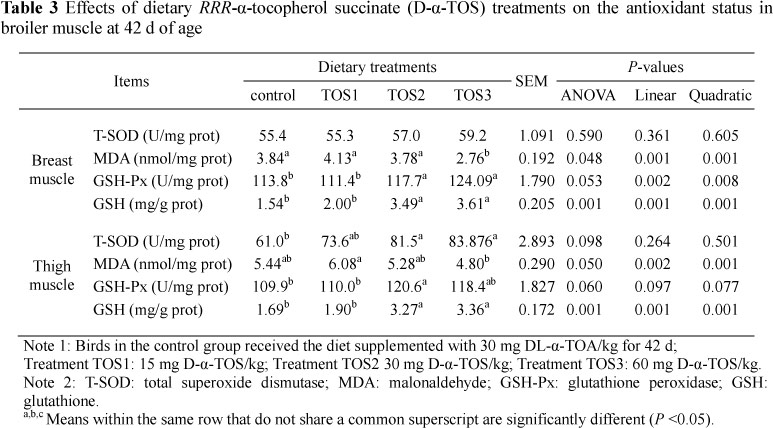
The results indicate that the T-SOD activities of thigh muscle in groups TOS2 and TOS3 exerted a 33.8% (P = 0.008) and 37.4% increase (P = 0.004), respectively, compared with the control group (Table 3). MDA concentrations of the breast muscle for group TOS3 were 28.1% (P = 0.015), 33.2% (P = 0.001), and 27.0% (P = 0.04) lower than concentrations in the control, TOS1 and TOS2 groups, respectively. Also, group TOS3 exerted the greatest effect, as it led to a decrease of 21.1% compared with the TOS1 group (P <0.05).
The GSH-Px activities and GSH contents of both breast and thigh muscles of the TOS2 and TOS3 groups were higher than in the control and TOS1 groups (P <0.05). However, corresponding measurements did not differ between the control and the TOS1 group (P >0.05).
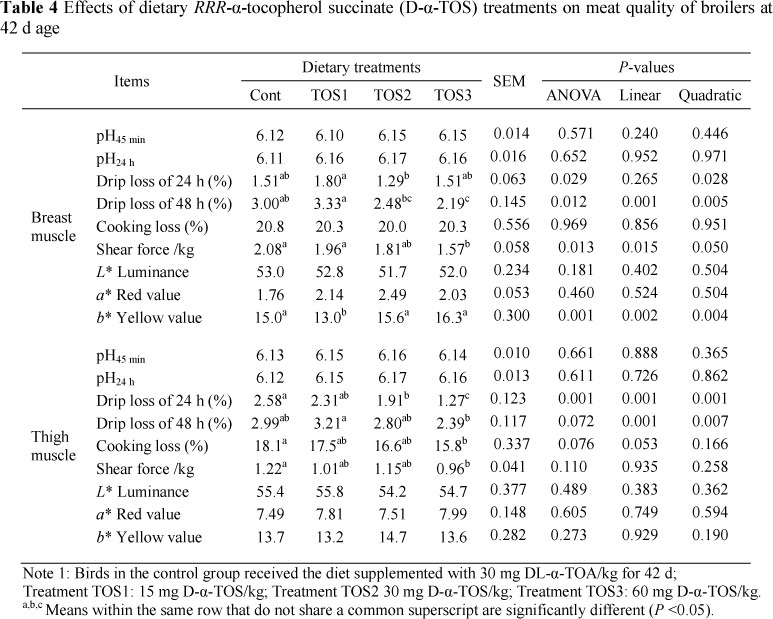
In the breast muscle, drip loss of 24 h for group TOS2 was lower (P <0.05) than in the TOS1 group (Table 4). Drip loss of 48 h for group TOS3 was lower (P <0.05) than that of the control group and TOS1 group. Also, group TOS2 exerted a decrease compared with the TOS1 group. Shear force for group TOS3 was decreased (P <0.05) in comparison with the control or TOS1 group.
In the thigh muscle, drip loss of 24 h for group TOS3 was decreased (P <0.05) by 50.78%, 45.02% and 33.51%, respectively, compared with the other three groups, and group TOS2 was lower than the control group (P <0.05). Differences in drip loss of 48 h were significant (P <0.05) between groups TOS3 and TOS1. Treatments with graded levels of D-a-TOS supplementation also brought about a 12.44% and 21.31% decrease for cooking loss and shear force in group TOS3 compared with the control group (P <0.05).
Discussions
It is generally agreed that the biological activity of RRR-a-tocopherol is greater than that of all-rac-α-tocopherol (Weiser & Vecci, 1982). Bioavailability of D-α-TOS has been investigated with aquatic animals, and it has been recorded that D-a-TOS is not readily available to juvenile red drum compared with the widely used DL-α-TOA. To date little research has been done on poultry. The bioavailability of these two a-tocopheryl esters might be species dependent, as well as influenced by the methodology for comparison. Using the commonly accepted conversion factor of 1.21 (Hidiroglou & Butler, 1990), the dietary intakes of 1 mg/kg of D-α-TOS would be expected to have similar biological activities to 1.21 mg DL-α-TOA/kg.
Compared with other meat, chicken meat is relatively abundant in PUFA, including the key n-3 fatty acids, and is easily attacked by free radicals (Asghar et al., 1990; Rhee et al., 1996). The reaction of 2-thiobarbituric acid with MDA is widely used for measuring the extent of oxidative deterioration of lipid in muscle foods (Descalzo & Sancho, 2008; Descalzo et al., 2008). Lipid oxidation results in the production of free radicals, which may lead to the oxidation of meat pigments and generation of rancid odours and flavours (Faustman & Cassens, 1989; 1990). Young et al. (2003) reported that MDA production was decreased in the pectoralis major of ascorbic acid-α-tocopherol-supplemented chickens.
It is clear that vitamin E acts as part of the cellular antioxidant systems in close cooperation with other cellular antioxidants, particularly the ascorbate and GSH systems. Lipid peroxidation may be exacerbated by dietary factors such as polyunsaturated fatty acid level. GSH is the natural intracellular ROS scavenger and is involved in the cellular defence mechanisms, thus protecting the tissue against oxidative damage, which is particularly important for the prevention of fatty acid peroxidation (Benedich, 1990).
In the present study, the increased sarcous GSH and decreased MDA concentrations at the moderate levels of D-α-TOS (30 - 60 mg/kg) in comparison with the control treatment may contribute to the potent free radical eliminating effect of D-α TOS by the modulation of the cellular free radical/antioxidant balance. Moreover, these results were consistent with the results of increased serum and hepatic a-tocopherol concentrations and hepatic ROS levels. Findings in the present study suggested that 30 mg/kg to 60 mg/kg of D-α-TOS supplementation in broilers may be needed to protect tissues against attack by lipid oxidation products.
Water-holding capacity (drip loss and cooking loss) of meat is directly related to the intramuscular lipids and moisture content of the meat and lean meat contains approximately 75% of water (Trout, 1988; Huff-Lonergan & Lonergan, 2005). Tenderness (shear force) may be the most important eating quality parameter that determines consumer acceptability (Miller et al., 2001; Kannan et al., 2002). In the present trial, the water-holding capacity and shear force were greatly improved by dietary supplementation of 30 and 60 mg/kg of D-a-TOS compared with the control group. In addition, a* (redness) tended to increase slightly (P >0.05) by dietary supplementation of 30 and 60 mg/kg of D-α-TOS. This indicated that dietary 30 and 60 D-α-TOS/kg supplementation can reduce the rate of surface discoloration in chicken meat. The similar results for vitamin E supplementation on muscle colour were determined for beef cattle (Liu et al., 1995; Mitsumoto et al., 1998) and pork (Asghar et al., 1991).
The improvement of meat quality must be related to the antioxidative status. In our study, the higher T-SOD and GSH-Px activity in the muscle of 30 mg/kg and 60 mg/kg of D-a-TOS-supplemented broilers compared with that of control broilers indicated that 30 mg/kg and 60 mg/kg of D-a-TOS enhanced antioxidant enzymatic activity of broilers. The antioxidant defences include natural and synthetic antioxidants and the antioxidant enzymes present in the biological system (Sies, 1991). Free radicals are produced during normal metabolism but can induce body damage if they are present in excessive levels. It has generally been recognized that T-SOD, GSH-Px and catalase are three main antioxidant enzymes in scavenging the oxygen free radical (McCord, 1979). Therefore, increasing activities of SOD and GSH-Px would subsequently enhance the capacity of broilers to clear out the oxygen free radicals. Consistent with the increased activity of serum T-SOD and GSH-Px, MDA concentration in the muscle was reduced by inclusion of 30 or 60 mg D-α-TOS/kg in broiler diets. Taking the ROS levels and a-tocopherol contents together, these results demonstrated that dietary 30 mg/kg and 60 mg/kg of D-a-TOS supplementation improved the antioxidant status of broiler chickens.
Conclusions
The results of the present experiment showed that dietary supplementation of 30 mg/kg and 60 mg/kg of D-a-TOS to broilers resulted in a general improvement of meat oxidative stability compared with the control diet. Specifically, antioxidant procedures, that is, 30 mg/kg and 60 mg/kg of D-α-TOS, protected chicken thigh and breast muscle from oxidative damage while improving its consumer acceptance.
Acknowledgements
This research was supported financially by a grant (Project No.2004CB117500.) from the National Basic Research Programme (973 Programme) of P. R. China. The authors gratefully acknowledged Zhu Qiufeng, Wang Longchang, Ma xia, Guo Zhenhuan, and Hou Xiang for their kind help. Thank you also to all the staff and students at the university who offered us help in this study.
References
Ao, X., Yan, L., Meng, Q.W., Zhou, T.X., Wang, J.P., Kim, H.J., Cho, J.H. & Kim, I.H., 2011. Effects of Saururus chinensis extract supplementation on growth performance, meat quality and slurry noxious gas emission in finishing pigs. Livest. Sci. 138, 187-192. [ Links ]
Asghar, A., Gray, J.I., Booren, A.M., Gomaa, E.A., Abouzied, M.M. & Miller, E.R., 1991. Effects of supranutritional dietary vitamin E levels on subcellular deposition of α-tocopherol in the muscle and on pork quality. J. Sci. Food Agric. 57, 31-41. [ Links ]
Asghar, A., Lin, C.F., Gray, J.I., Buckley, D.J., Booren, A.M. & Flegal, C.J., 1990. Effects of dietary oils and α-tocopherol supplementation on membranal lipid oxidation in broiler meat. J. Food Sci. 55, 46-50. [ Links ]
Benedich, A., 1990. Antioxidant vitamins and their function in immune response. In: Advances in Experimental Medicine and Biology, Vol. 262. Ed. Benedich, A., Plenum Press, New York, NY. pp. 35-55. [ Links ]
Boler, D.D., Gabriel, S.R., Yang, H., Balsbaugh, R., Mahan, D.C., Brewer, M.S., McKeith, F.K. & Killefer, J., 2009. Effect of different dietary levels of natural-source vitamin E in grow-finish pigs on pork quality and shelf life. Meat Sci. 83, 723-730. [ Links ]
Brigelius-Flohe', R., Kelly, F.J., Salonen, J.T., Neuzil, J., Zingg, J.M. & Azzi, A., 2002. The European perspective on vitamin E: current knowledge and future research. Am. J. Clin. Nutr. 76, 703-716. [ Links ]
Cardenia, V., Rodriguez-Estrada, M.T., Cumella, F., Sardi, L., Della, C.G. & Lercker, G., 2011. Oxidative stability of pork meat lipids as related to high-oleic sunflower oil and vitamin E diet supplementation and storage conditions. Meat Sci. 88, 271-279. [ Links ]
Corino, C., Oriani, G., Pantaleo, L., Pastorelli, G. & Salvatori, G., 1999. Influence of dietary vitamin E supplementation on "heavy pig" carcass characteristics, meat quality and vitamin E status. J. Anim. Sci. 77, 1755-1761. [ Links ]
Descalzo, A.M. & Sancho, A.M., 2008. A review of natural antioxidants and their effects on oxidative status, odor and quality of fresh beef produced in Argentina. Meat Sci. 79, 423-436. [ Links ]
Descalzo, A.M., Rossetti, L., Sancho, A.M., Garcia, P.T., Biolatto, A., Carduza, F. & Grigioni, G.M., 2008. Antioxidant consumption and development of oxidation during ageing of buffalo meat produced in Argentina. Meat Sci. 79, 582-588. [ Links ]
Dufrasne, I., Marche, C., Clinquart, A., Hornick, J.L., Van Eenaeme, C. & Istasse, L., 2000. Effects of dietary vitamin E supplementation on performance and meat characteristics in fattening bulls from Belgian Blue breed. Livest. Prod. Sci. 65, 197-201. [ Links ]
Eder, K., Muller, G., Kluge, H., Hirche, F. & Brandsch, C., 2005. Concentrations of oxysterols in meat and meat products from pigs fed diets differing in the type of fat (palm oil or soybean oil) and vitamin E concentrations. Meat Sci. 70, 15-23. [ Links ]
Fariss, M.W., 1990. Oxygen toxicity: unique cytoprotective properties of vitamin E succinate in hepatocytes.Free Radical Biol. Med. 9, 333-343. [ Links ]
Fariss, M.W., Pascoe, G.A. & Reed, D.J., 1985. Vitamin E reversal of the effect of extracellular calcium on chemically induced toxicity in hepatocytes. Science 227, 751-754. [ Links ]
Fariss, M.W., Bryson, K.F. & Tirmenstein, M.A., 1997. Role of cellular thiol status in tocopheryl hemisuccinate cytoprotection against ethyl methanesulfonate-induced toxicity. Biochem. Pharmacol. 53, 651-661. [ Links ]
Fariss, M.W., Nicholls-Grzemski, F.A., Tirmenstein, M.A. & Zhang, J.G., 2001. Enhanced antioxidant and cytoprotective abilities of vitamin E succinate is associated with a rapid uptake advantage in rat hepatocytes and mitochondria. Free Radical Biol. Med. 31, 530-541. [ Links ]
Faustman, C. & Cassens, R.G., 1989. Strategies for improving fresh meat colour. In: Proc. 35th Int. Congr.,Meat Sci. Technol., Copenhagen, Denmark. pp. 446-453. [ Links ]
Faustman, C. & Cassens, R.G., 1990. The biochemical basis for discoloration in fresh meat: A review. J. Muscle Foods 1, 217-243. [ Links ]
Gray, J.I., Gomaa, E.A. & Buckley, D.J., 1996. Oxidative quality and shelf life of meats. Meat Sci. 43, S111-S123. [ Links ]
Guo, Q., Richert, B.T., Burgess, J.R., Webel, D.M., Orr, D.E., Blair, M., Fitzner, G.E., Hall, D.D., Grant, A.L. & Gerrard, D.E., 2006a. Effects of dietary vitamin E and fat supplementation on pork quality. J. Anim. Sci. 84, 3089-3099. [ Links ]
Guo, Q., Richert, B.T., Burgess, J.R., Webel, D.M., Orr, D.E., Blair, M., Grant, A.L. & Gerrard, D.E., 2006b. Effect of dietary vitamin E supplementation and feeding period on pork quality. J. Anim. Sci. 84, 3071-3078. [ Links ]
Hafeman, D.G., 1974. Effect of dietary selenium on erythrocyte and liver glutathione peroxidase in the rat. J. Nutr. 104, 580-587. [ Links ]
Hidiroglou, M. & Butler, G., 1990. Vitamin E response in sheep dosed intraruminally or intraduodenally with various alpha-tocopherol compounds. Int. J. Vitam. Nutr. Res. 60, 331-337. [ Links ]
Huff-Lonergan, E. & Lonergan, S.M., 2005. Mechanisms of water-holding capacity of meat: The role of post-mortem biochemical and structural changes. Meat Sci. 71, 194-204. [ Links ]
Jensen, C., Guidera, J., Skovgaard, I.M., Staun, H., Skibsted, L.H., Jensen, S.K., Moller, A.J., Buckley J. & Bertelsen, G., 1997. Effects of dietary alpha-tocopheryl acetate supplementation on alphatocopherol deposition in porcine m. psoas major and m. longissimus dorsi and on drip loss, colour stability and oxidative stability of pork meat. Meat Sci. 45, 491-500. [ Links ]
Jensen, C., Lauridsen, C. & Bertelsen, G., 1998. Dietary vitamin E: Quality and storage stability of pork and poultry. Trends Food Sci. Technol. 9, 62-72. [ Links ]
Jensen, S.K., Engberg, R.M. & Hedemann, M.S., 1997. All-rac-a-tocopherol acetate is a better vitamin E source than all-rac-a-tocopherol succinate for broilers. J. Nutr. 129, 1355-1360. [ Links ]
Kannan, G., Chawan, C.B., Kouakou, B. & Gelaye, S., 2002. Influence of packaging method and storage time on shear value and mechanical strength of intramuscular connective tissue of chevon. J. Anim.Sci. 80, 2383-2389. [ Links ]
Karami, M., Alimon, A.R. & Goh, Y.M., 2011. Effect of vitamin E, Andrographis paniculata and turmeric as dietary antioxidant supplementation on lipid and color stability of goat meat. Small Rumin. Res. 97, 67-71. [ Links ]
Kayden, H.J., Chow, C.K. & Bjornson, L.K., 1973. Spectrophotometric method for determination of tocopherol in red blood cells. J. Lipid Res. 14, 533-540. [ Links ]
Kim, B.H. & Takemura, A., 2003. Culture conditions affect induction of vitellogenin synthesis by estradiol-17beta in primary cultures of tilapia hepatocytes. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 135, 231-239. [ Links ]
LeBel, C.P., Ischiopoulos, H. & Bondy, S.C., 1992. Evaluation of the probe 2',7'-dichlorofluorescin as indicator of reactive oxygen species formation and oxidative stress. Chem. Res. Toxicol. 5, 227-231. [ Links ]
Liu, Q., Lanari, M.C. & Schaefer, D.M., 1995. A review of dietary vitamin E supplementation for improvement of beef quality. J. Anim. Sci. 73, 3131-3140. [ Links ]
Luciano, G., Monahan, F.J., Vasta, V., Pennisi, P., Bella, M. & Priolo, A., 2009. Lipid and colour stability of meat from lambs fed fresh herbage or concentrate. Meat Sci. 82, 193-199. [ Links ]
McCord, J.M., 1979. Superoxide, superoxide dismutase and oxygen toxicity. Rev. Biochem. Toxicol. 1109-1124. [ Links ]
Meek, K.I., Claus, J.R., Duncan, S.E., Marriott, N.G., Solomon, M.B., Kathman, S.J. & Marini, M.E., 2000. Quality and sensory characteristics of selected post-rigor, early-deboned broiler breast meat tenderized using hydrodynamic shock waves. Poult. Sci. 79, 126-136. [ Links ]
Miller, M.F., Carr, M.F., Ramsey, C.B., Crockett, K.L. & Hoover, L.C., 2001. Consumer thresholds for establishing the value of beef tenderness. J. Anim. Sci. 79, 3062-3068. [ Links ]
Mitsumoto, M., Ozawa, S., Mitsuhashi, T. & Koide, K., 1998. Effect of dietary vitamin E supplementation for one week before slaughter on drip loss, colour and lipid stability duringdisplay in Japanese Black steer beef. Meat Sci. 49, 165-174. [ Links ]
Monahan, F.J., Asghar, A., Gray, J.I., Buckley, D.J. & Morrissey, P.A., 1994. Effect of oxidized dietary lipid and vitamin E on the colour stability of pork chops. Meat Sci. 37, 205-215. [ Links ]
Neuzil, J., Dong, L.F., Ramanathapuram, L., Hahn, T., Chladova, M., Wang, X.F., Zobalova, R., Prochazka, L., Gold, M., Freeman, R., Turanek, J., Akporiaye, E.T., Dyason, J.C. & Ralph S.J., 2007. Vitamin E analogues as a novel group of mitocans: Anti-cancer agents that act by targeting mitochondria. Mol. Aspects Med. 28, 607-645. [ Links ]
Nuala, T.M.M., Sean, A.H. & Kerry, J.P., 2006. Comparative addition of rosemary extract and additives on sensory and antioxidant properties of retail packaged beef. Int. J. Food Sci. Technol. 42, 1201-1207. [ Links ]
Onibi, G.E., Scaife, J.R., Murray, I. & Fowler, V.R., 2000. Supplementary a-tocopherol acetate in full-fat rapeseed-based diets for pigs: Effect on performance, plasma enzymes and meat drip loss. J. Sci. Food Agric. 80, 1617-1624. [ Links ]
Oyanagui, Y., 1984. Reevaluation of assay methods and establishment of kit for superoxide dismutase activity. Anal. Biochem.142, 290-296. [ Links ]
Placer, Z.A., Cushman, L.L. & Johnson, B.C., 1966. Estimation of product of lipid peroxidation (malonyl dialdehyde) in biochemical systems. Anal. Biochem. 16, 359-364. [ Links ]
Ramirez, J.A., Oliver, M.Á., Pla, M., Guerrero, L., Arino, B., Blasco, A., Pascual, M. & Gil, M., 2004. Effect of selection for growth rate on biochemical, quality and texture characteristics of meat from rabbits. Meat Sci. 67, 617-624. [ Links ]
Rhee, K.S., Anderson, L.M. & Sams, A.R., 1996. Lipid oxidation potential of beef, chicken and pork. J. Food Sci. 61, 8-12. [ Links ]
Roos, D.H., Puntel, R.L., Santos, M.M., Souza, D.O.G., Farina, M., Nogueira, C.W., Aschner, M., Burger, M.E., Barbosa, N.B.V. & Rocha, J.B.T., 2009. Guanosine and synthetic organoselenium compounds modulate methylmercury-induced oxidative stress in rat brain cortical slices: involvement of oxidative stress and glutamatergic system. Toxicol. in Vitro 23, 302-307. [ Links ]
Sahin, K., Sahin, N., Onderci, M. & Yaralioglu, S., 2002. Effects of vitamins E and A supplementation on lipid peroxidation and concentration of some mineral in broilers reared under heat stress (32 °C). Nutr. Res. 22, 723-731. [ Links ]
Sies, H., 1991. Oxidative stress: From basic research to clinical application. Am. J. Med. 91, 31-38. [ Links ]
Trout, G.T.R., 1988. Techniques for measuring water-binding capacity in muscle foods: A review of methodology. Meat Sci. 23, 235-252. [ Links ]
Villaverde, C., Baucells, M.D., Manzanilla, E.G. & Barroeta, A.C., 2008. High levels of dietary unsaturated fat decrease α-tocopherol content of whole body, liver, and plasma of chickens without variations in intestinal apparent absorption. Poult. Sci. 87, 497-505. [ Links ]
Wang, K.H., Shi, S.R., Dou, T.C. & Sun, H.J., 2009. Effect of a free-range raising system on growth performance, carcass yield, and meat quality of slow-growing chicken. Poult. Sci. 88, 2219-2223. [ Links ]
Weiser, H. & Vecci, M., 1982. Stereoisomers of α-tocopheryl acetate. II. Biopotencies of all eight stereoisomers, individually or in mixtures, as determined by rat resorption-gestation tests. Int. J. Vitam. Nutr. Res. 52, 351-370. [ Links ]
Young, J.F., Stagsted, J., Jensen, S.K., Karlsson, A.H. & Henckel, P., 2003. Ascorbic acid, α-tocopherol, and oregano supplements reduce stress-induced deterioration of chicken meat quality. Poult. Sci. 82, 1343-1351. [ Links ]
Young, J.F., Karlsson, A.H. & Henckel, P., 2004. Water-holding capacity in chicken breast muscle is enhanced by pyruvate and reduced by creatine supplements. Poult Sci. 83, 400-405. [ Links ]
Zaspel, B.J. & Csallany, A.S., 1983. Determination of alpha-tocopherol in tissues and plasma by high-performance liquid chromatography. Anal. Biochem. 130, 146-150. [ Links ]
Zhang, J.G., Nicholls-Grzemski, F.A., Tirmenstein, M.A. & Fariss, M.W., 2001. Vitamin E succinate protects hepatocytes against the toxic effect of reactive oxygen species generated at mitochondrial complexes I and III by alkylating agents. Chemico-Biol. Interact. 138, 267-284. [ Links ]
Zhang, X.H., Zhong, X., Zhou, Y.M., Du, H.M. & Wang, T., 2009. Effect of RRR-α-tocopherol succinate on the growth and immunity in Broilers. Poult. Sci. 88, 959-966. [ Links ]
Zhang, X.H., Zhong, X., Zhou, Y.M., Wang, G.Q., Du, H.M. & Wang, T., 2010. Dietary RRR-α-tocopherol succinate attenuates LPS-induced inflammatory cytokines secretion in broiler chicks. Br. J. Nutr. 2010, 1, 1-10. [ Links ]
Zhou, B.S., Liu, C.S., Wang, J.X., Lam, P.K. & Wu, R.S., 2006. Primary cultured cells as sensitive in vitro model for assessment of toxicants-comparison to hepatocytes and gill epithelia. Aquat. Toxicol. 80, 109-118. [ Links ]
Zhu, Y.M., Miao, J.F., Fan, H.J., Zou, S.X. & Chen, W.H., 2007. Protective effect of CpG-DNA against mastitis induced by Staphylococcus aureus infection in a rat model. Int. Immunopharmacol. 7, 435-443. [ Links ]
Zingg, J.M., 2007. Vitamin E: An overview of major research directions. Mol. Aspects Med. 28, 400-422. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. See: http://creativecommons.org/licenses/by/2.5/za Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science.
# Corresponding author: tianwangnjau@163.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}