










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.42 n.3 Pretoria Jan. 2012
Effect of dietary starch source on milk production and composition of lactating Holstein cows
GH.R. MosaviI; F. FatahniaI, #; H.R. Mirzaei AlamoutiII; A.A. MehrabiI; H. Darmani KohiIII
IDepartment of Animal Science, University of Ilam, Ilam, Iran
IIDepartment of Animal Science, University of Zanjan, Zanjan, Iran
IIIDepartment of Animal Science, University of Guilan, Rasht, Iran
ABSTRACT
The objective of this study was to evaluate the effects of four sources of starch on milk production and composition, nutrient digestion and blood metabolites of lactating Holstein cows. Four multiparous Holstein cows (708 ± 70 kg of body weight; 83 ± 9 days in milk) were used in a 4 χ 4 Latin square design with 21-d periods. The concentrate portion of the diet contained wheat, barley, maize or potato as the primary source of starch. Intake of dry matter (DM) ranged from 18.7 kg/d to 19 kg/d, and was similar among treatments. Milk production was higher in cows fed the wheat-based diet compared with other diets while the milk fat concentration of the cows fed the maize-based diet was the highest. Milk protein concentration was unaffected by the source of dietary starch. Cows fed the potato-based diet had a lower milk protein, lactose and solid-non-fat yield. Milk yield/kg of DM intake and net energy for lactation (NEL)/NEL intake were higher in cows fed wheat-, barley- or maize-based diets compared with those fed the potato-based diet. Feed nitrogen efficiency was higher in cows fed the maize-based diet compared with the other experimental diets. Total tract apparent digestibility of organic matter, crude protein and ether extract were higher in cows fed the wheat- or maize-based diets compared with those fed barley- or potato-based diets; however, total tract apparent digestibility of neutral detergent fibre and acid detergent fibre were higher in cows fed the wheat-based diet compared with those fed the potato-based diet. These results showed that improved production performance in cows fed the wheat-based diet appeared to be because of greater nutrient digestibility and greater nutrient utilization efficiency. Furthermore, potato starch is not superior to grain starch as a readily available energy source for lactating dairy cows.
Keywords: Nutrient digestibility, nutrient efficiency, plasma metabolites, wheat, barley, potato, maize
Introduction
The increase in milk yield of dairy cows that results from their genetic improvement requires the use of large amounts of concentrates that are rich in energy and crude protein (CP) to meet their nutrients requirements (Cabrita et al., 2009). Dietary carbohydrate is composed of neutral detergent fibre (NDF) and non-fibre fractions, which collectively compose 65% to 75% of the diets of lactating dairy cattle. Non-fibre carbohydrates (NFC) may provide 30 to 45% of the diet on a dry matter (DM) basis (Hall et al., 2010). Root crops such as potato have been used in dairy rations, but have been replaced by grains and maize silage because of labour costs (Eriksson et al., 2004). Dairy cow diets usually contain barley, maize and wheat as the main carbohydrate sources because they are cost-effective sources of digestible energy. Starch is the major nutrient providing energy from these cereal grains and potato. These cereal grains and potato differ in their starch content, with wheat containing (DM basis) 77% starch, maize 72%, barley 58% (Huntington, 1997) and potato 63% (Wang et al., 2009). Differences also exist among these starch sources in their rates and extents of ruminal starch degradation, with 32%/h for wheat starch, 2%/h for maize starch, 29%/h for barley starch, and 5%/h for potato starch being digested in the rumen (Monteils et al., 2002; Wang et al., 2009). The larger granules of more enzyme-resistant B-type crystalline starch in potato compared to the smaller granules of grain, generally with A-type crystalline starch (Tester et al., 2006) could explain the difference in their ruminal fermentation patterns. Raw potato starch could not be expected to be superior to grain starch as a readily available energy source, but it could reduce diurnal fluctuations in energy supply and limit the problems caused by the incorporation of a large amount of wheat or barley starch in the diet. In principle, the rate and extent of fermentation of dietary carbohydrates (especially starch) in the rumen are important parameters that determine nutrient supply to the animal (Hall, 2004). Greater dietary concentration of nonstructural carbohydrates increases the utilization of ruminal ammonia-N for microbial protein synthesis (Nocek & Tamminga, 1991). Increasing ruminally available energy concentration of diets for dairy cows has the potential to enhance milk production through increased metabolizable nutrient supply (Gozho & Mutsvangwa, 2008). Most studies on the effect of starch source have been conducted with grains (barley, maize, millet, oat, sorghum or wheat), but rarely with potato. The study of different sources of starch using in sacco technique (Monteils et al., 2002; Wang et al., 2009) showed that the ruminal degradation of potato starch is slower than that of wheat or barley, but faster than maize starch. These observations have been confirmed by enzymatic tests with ruminal fluid (Con, 1991). Owing to differences in digestion characteristics and fermentation products, the starch source of the diet has the potential to alter feed intake, milk production and milk composition. Therefore, the objective of the study was to evaluate the effect of dietary inclusion of barley, maize, wheat or potatoes as a principle source of starch on milk production and composition, nitrogen and energy efficiency, nutrient digestibility and blood metabolites of lactating dairy cows.
Materials and Methods
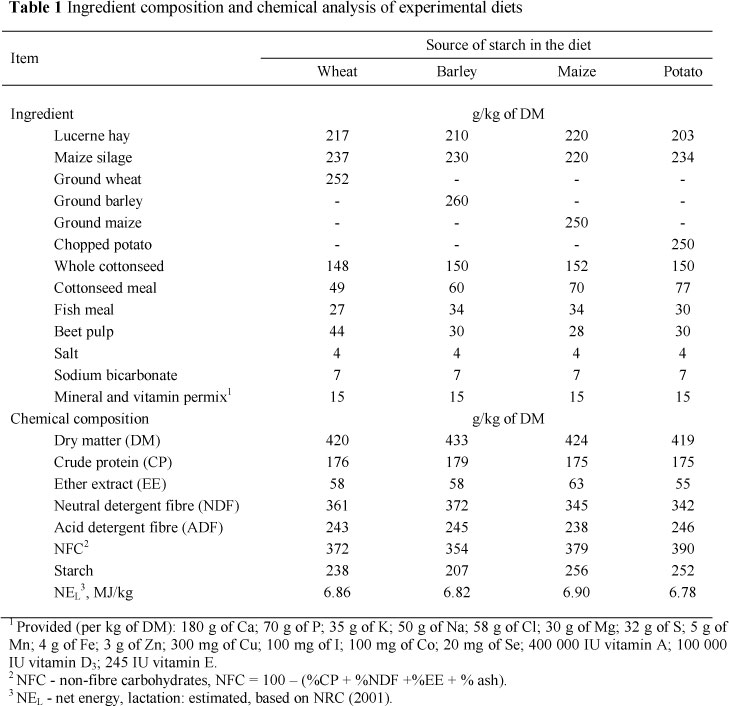
Four multiparous Holstein cows (708 ± 70 kg of BW; 83 ± 9 days in milk; 30 ± 1.3 kg of milk/d) were used in a 4 χ 4 Latin square design. Each experimental period consisted of 14 days for adaptation to the diets and 7 days for daily data collection. The cows were housed in individual tie stalls and had free access to drinking water throughout the trial. The four dietary treatments examined, consisted of different sources of starch, with the concentrate portion of the diet containing barley, maize, wheat or potato. All cereal grains were coarsely ground, and potato was chopped. The ingredient composition and chemical analysis of the experimental diets are shown in Table 1. The experimental diets were formulated according to the NRC (2001) recommendations, and fed individually twice daily at 08:00 and 16:00 as total mixed ration (TMR) for ad libitum intake and adjusted for 100 g orts/kg as fed. The cows were milked three times daily at 08:30, 16:00 and 24:00.
During the 7-d sample and data collection period, individual cow feed intake was recorded daily. Samples of diets and orts were collected daily, frozen at -20 °C, and composited per cow for each experimental period. Composite samples of orts were based on daily amounts of orts for each cow. After the experiment, pooled samples of the diets (TMR) and orts were dried at 60 °C for 48 h, ground through a 1mm screen using a Wiley mill, and analysed for DM, organic matter (OM), ether extract (EE), CP (AOAC, 1990), acid detergent fibre (ADF) (Van Soest et al., 1991) and NDF with heat-stable α-amylase and sodium sulphite (Van Soest et al., 1991). The starch concentration of the experimental diets (TMR) was analysed by an enzymatic method (Karkalas, 1985). Faecal grab samples were collected for six consecutive days at 08:00 on d 15, 10:00 on d 16, 12:00 on d 17, 14:00 on d 18, 16:00 on d 19, and 18:00 on d 20 of each experimental period. Faecal samples were dried at 55 °C in a forced draft oven for 72 h, then ground through a 1 -mm screen using a Wiley mill. Equal DM from each faecal subsample was mixed to obtain a single composite for each sampled cow during each period and analysed for OM, CP, EE, ADF and NDF, using the methods already described for pooled TMR and orts samples. Total tract nutrient digestibility was determined by acid insoluble ash (AIA) as an internal marker (Van Keulen & Young, 1977). Milk production was recorded during the 7-d data collection period. On days 16 and 17 of each experimental period, milk samples were collected from the six consecutive milking into plastic vials that contained a preservative (dichromate potassium, K2Cr2O7). Milk samples were then pooled daily, based on milk yield, and kept at room temperature for determination of protein, fat and lactose concentrations (Milko-Scan 133B Foss Electric, Denmark). On d 19 of each experimental period, blood samples were drawn before the morning feeding from the jugular vein into vacutainer tubes containing heparin. The plasma was separated by centrifugation (2,500 χ g for 20 min) and total cholesterol (kit no. 10-508; ZiestChem Diagnostics Co., Tehran, Iran), high-density lipoprotein (HDL) cholesterol (kit no. 10-507; ZiestChem Diagnostics Co., Tehran, Iran), triglycerides (kit no. 10-525; ZiestChem Diagnostics Co., Tehran, Iran ), and glucose (kit no. 10-505; ZiestChem Diagnostics Co., Tehran, Iran) concentrations were analysed by colorimetric methods. Low-density lipoprotein (LDL) cholesterol was calculated by difference between total cholesterol and high-density lipoprotein (HDL) cholesterol.
All statistical analyses were performed using PROC MIXED of SAS (1999). Data on DM intake, milk production, milk composition, nutrient digestibility, faecal excretion, nitrogen and energy efficiency, and blood metabolites were analyzed as a 4 χ 4 Latin square design using the following model:
Yijk = µ + Ti + Cj + Pk + eijk, where Yijk is the dependent variable, µ is the overall mean, Ti is the fixed effect of treatment, Cj is the random effect of cow, Pk is the fixed effect of period, and eijk is the random residual error. Treatment effects were declared significance at P <0.05.
Results and Discussion
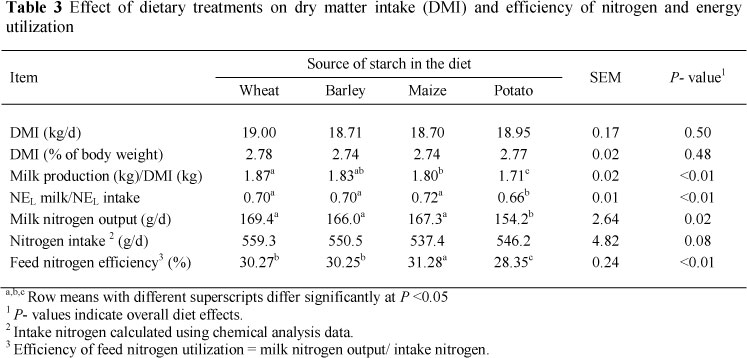
Actual milk production was significantly higher (P <0.05) in cows fed the wheat-based diet compared with those fed the other diets. On average, cows fed barley-, maize-, and potato-based diets produced 1.4, 1.74, and 3.54 kg/d less milk, respectively, compared with those fed a wheat-based diet (Table 2). The potato-based diet produced significantly less milk than the other three diets, but there was no significant difference when replacing maize with barley. Fat corrected milk (FCM) and energy corrected milk (ECM) were higher in cows fed wheat-, or maize-based diets compared with those fed barley- or potato-based diets (Table 2). Similarly, Silveira et al. (2007) reported higher 4% FCM in cows fed maize-based diet compared with those fed barley-based diet, but, in a study by Gozho & Mutsvangwa (2008), 3.5% FCM was higher in cows fed barley- and maize-based diets compared with a wheat-based diet.
In studies of Cabrita et al. (2009), with maize and wheat, and Silveira et al. (2007), with maize and barley as the main sources of dietary starch, actual milk production was increased for cows fed the maize-containing diets compared with those fed wheat- or barley-containing diets. However, Gozho & Mutsvangwa (2008) with barley, maize, wheat and oats, Khorasani et al. (2001), with maize and barley, and Jurjanz et al. (1998), with wheat and potato peelings, did not observe any effect of starch source on milk production. Responses of lactating cows to different cereal grains depend on the level of dietary inclusion, the basal ration, physical processing of the cereal grains, the composition of a given batch of cereal grain, and the level of dietary intake (Khorasani et al, 2001). Although in our experiment DM intake was not affected by experimental treatments, the higher milk production in cows fed the wheat-based diet was attributed to greater total tract digestibility of DM (Table 4) and higher efficiency for milk production, defined as milk production/DMI (Table 3), although not for NEL milk/NEL intake. The reduction in milk production for cows fed potato-based diet was attributed to a lower total tract digestibility of DM and higher faecal excretion of DM (Table 4). However, in the current experiment, actual milk production was significantly higher for cows fed the wheat-based diet compared with those fed the maize-based diet, their similar 4% FCM production can be explained by higher milk fat content of cows fed maize-based diet.
In the current study, feeding the maize-based diet resulted in a higher milk fat concentration (P <0.05) compared with the feeding of the wheat-, barley-, or potato-based diets (Table 2). A higher fat concentration in milk from cows fed maize compared with barley or wheat diets has been reported previously (Gozho & Mutsvangwa, 2008; Cabrita et al, 2009). However, other research observed no effect of starch source on milk fat concentration (Khorasani et al., 2001; Silveira et al, 2007). Jurjanz et al. (1998) did not observe any dietary effects on milk fat concentration with diets containing potato peelings or wheat as the principal source of starch at low and medium starch concentration, but milk fat concentration was higher when potato peelings were fed at the higher starch concentration. Observed differences in milk fat concentration undoubtedly reflect differences in patterns of ruminal fermentation (Moren, 1986) and in site and the extent of digestion of these starch sources (Okine & Kennelly, 1994). Slower degradation of maize starch compared with wheat, barley or potato starches in the rumen could enhance a higher delivery of precursors (acetate and butyrate) of milk fat synthesis to the udder. The rumen is the major site of starch digestion in cattle (Taniguchi et al., 1995). In vivo studies (Sauvant et al, 1994; Sauvant, 1997) showed a decreased acetate + butyrate to propionate ratio for cows fed diets containing starch that was degraded quickly in the rumen. In situations where the diet promotes lower ruminal pH, the biohydrogenation pathway for the saturation of 18-carbon polyunsaturated FA can be perturbed, with a resultant increase in C18:1 trans-FA. Some of these trans-FA have been shown to be potent inhibitors of milk fat synthesis (Griinari et al., 1998).
Dietary treatment did not affect the protein concentration in milk (P >0.05), but wheat-, barley-, or maize-based diets increased (P <0.05) milk protein yield compared with the potato-based diet (Table 2). Similarly, previous research (Khorasani et al., 2001; Silveira et al., 2007; Cabrita et al., 2009) found no differences in milk protein concentration of cows fed diets containing rapidly degradable starch compared with slowly degradable starch. Gozho & Mutsvangwa (2008), investigating wheat, barley, maize and oats as principal sources of starch, found that milk protein concentration was higher in cows fed a maize-based diet compared with those fed an oats-based diet. However, they observed no differences in milk protein concentration in cows fed wheat-, barley-, or maize-based diets.
Milk lactose concentration did not differ (P >0.05) among dietary treatments (Table 2). Similarly, other research observed no effect of starch source on milk lactose concentration (Khorasani et al., 2001; Gozho & Mutsvangwa, 2008; Cabrita et al., 2009).
Dry matter intake was not affected (P >0.05) by dietary source of starch (Table 3). Cabrita et al. (2009) reported similar DM intakes in cows fed maize-based diets compared with wheat-based diets. Gozho & Mutsvangwa (2008) did not observe any effect of dietary starch source on DM intake in cows fed wheat-, barley-, maize- or oats- based diets. Jurjanz et al. (1998) did not observe any dietary effects on DM intake of dairy cows fed diets containing wheat or potato peelings as the principal sources of starch. In contrast, Silveira et al. (2007) reported higher DM intakes in cows fed a maize-based diet compared with those fed a barley-based diet. A review by Cabrita et al. (2006) found contradictory effects of changing starch fermentability on DM intake of lactating dairy cows. With diets containing rapidly degradable starch, a decrease in rumen pH and an increase in volatile fatty acid production may decrease DM intake by regulatory mechanisms. Inconsistent effects of starch source on DM intake can be attributed to the starch concentration of the diet (DeVisser et al., 1990), and forage particle length (Rode & Satter, 1988). Diets containing higher concentrations of forage NDF promote chewing, salivation and high rumen pH (Mertens, 1997), which mask some effects caused by differences in fermentability of diets in the rumen.
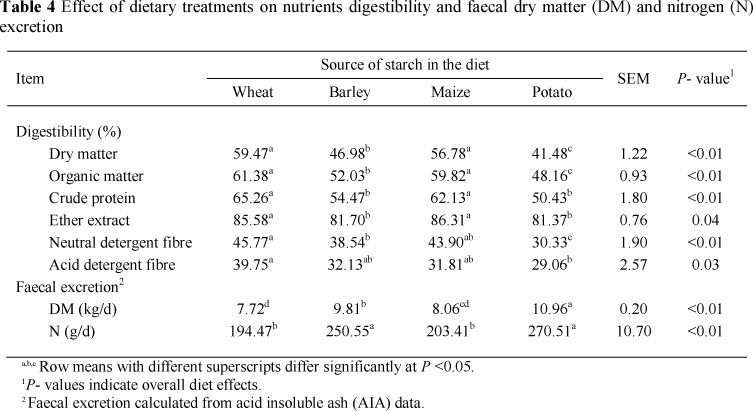
Efficiency for milk production, defined either as milk yield/DM intake or as NEL milk/NEL intake, was significantly (P >0.05) higher in cows fed wheat-, barley- or maize-based diets compared with the potato-based diet (Table 3). Contrary to the results of this study, Silveira et al. (2007) reported a higher NEL milk/NEL intake in cows fed barley-based diets than for cows fed maize-based diets, but milk yield/DM intake was not affected by grain source. It is generally recognized that the efficiency of utilization of energy from starch is greater when it is digested in the small intestine and absorbed as glucose rather than when starch is fermented in the rumen and the propionate fraction is converted to glucose in the liver (Reynolds, 2006). However, the effects of this higher energetic efficiency on production response are equivocal. Theurer et al. (1999) concluded that higher ruminal starch digestion increases milk production. In the current study, lower efficiency for milk production in cows fed potato-based diet could be attributed to lower total tract digestibility of DM and higher faecal DM excretion (Table 4). Nitrogen intake (Table 3) was similar across diets, in part because DM intake (Table 3) was not affected by starch source and also because the diets were formulated to be isonitrogenous. Feed nitrogen efficiency was lowest in cows fed the potato-based diet, intermediate in those fed the barley-, or wheat-based diets, and greatest in cows fed the maize-based diet (Table 3), reflecting the lower total tract digestibility of CP and the higher faecal nitrogen excretion (Table 4) in cows fed the potato-based diet. In the current study, the lower feed nitrogen efficiency in cows fed the potato-based diet might be partly explained by lower microbial protein synthesis and higher ammonia concentration in the rumen. Surber & Bowman (1998) speculated that increased ruminal and post ruminal digestibility of barley starch, compared with maize starch, may result in a greater energy yield and an improved feed conversion. They reported 17% greater microbial nitrogen synthesis for steers fed barley than for those fed maize. In our experiment, the lower starch content in barley-based diet (20.7% of DM) and consequently the lower available energy for microbial protein synthesis could partly explain the lower feed nitrogen efficiency when compared with the maize-based diet. Theurer et al. (1999) showed that a decrease in total starch or rumen degradable starch intake was associated with a decrease in microbial nitrogen flow in dairy cows. Considering that DM intake was not affected by dietary treatments (Table 3) and the lower starch content of barley, the barley-based diet was expected to result a lower starch intake when compared with the maize-based diet.
Total tract digestibility of DM, OM, CP and EE was higher in cows fed the wheat- or maize-based diets than those fed the barley- or potato-based diets (Table 4). Total tract ADF and NDF digestibility were higher in cows fed the wheat-based diet than cows fed the barley-, maize- or potato-based diets (Table 4). Similarly, Silveira et al. (2007) reported higher DM, OM and EE digestibility for cows fed a maize-based diet compared with those fed a barley-based diet. However, CP and NDF digestibility were not affected by experimental diets. In another comparative study with wheat, barley, maize and oats as principal sources of dietary starch, digestibility of DM, OM, and NDF was not affected by dietary treatments, but CP digestibility was higher in cows fed oats-based diet compared with those fed wheat-, barley- or maize-based diets (Gozho & Mutsvangwa, 2008). In the current study, the lower CP digestibility in cows fed the barley- or potato-based diets compared with those fed the wheat-, or maize-based diets might be related to the lower extent of ruminal starch degradation and higher undigested starch reaching the hindgut. Higher levels of undigested starch reaching the hindgut might have promoted more bacterial protein synthesis (0rskov et al., 1970). Because there is no mechanism for hindgut enzymatic digestion of the resultant bacterial protein, it is voided in the faeces, thus reducing total tract CP digestibility (0rskov et al., 1970). In addition, differences in CP digestibility of experimental diets may be due to differences in the digestibility of the cereal grains protein (McAllister et al, 1993).
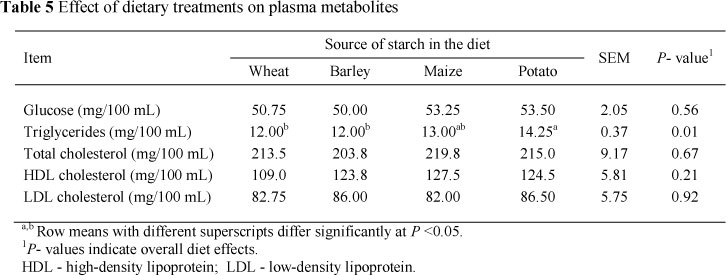
There were no differences in plasma glucose, total cholesterol, HDL-cholesterol and LDL-cholesterol concentrations between dietary starch sources. However, plasma triglyceride concentrations were higher in cows fed the potato-based diets compared with the wheat-, barley-, or maize-based diets (Table 5). We expected the lower plasma triglyceride concentrations for cows fed the potato-based diet compared with the other diets (especially the wheat- and maize-based diets) because of the lower EE digestibility (Table 4). Contrary to expectations, plasma triglycerides concentration was higher in cows fed the potato-based diet. Similar plasma glucose concentrations were reported in studies of Cabrita et al. (2009) with maize and wheat and Silveira et al. (2007) with maize and barley as the main sources of dietary starch.
Conclusions
This study showed that feeding diets containing wheat, barley, maize or potato resulted in similar DM intake. Cows fed the wheat-based diet had a higher milk production and efficiency of milk production. Milk fat concentrations were higher for cows fed the maize-based diet resulting in no differences in FCM, ECM and NEL milk/NEL intake for the wheat, barley and maize diets, with potato diets recording lower values. Milk protein concentration was not affected by starch source. Feeding maize-based diets increased feed nitrogen efficiency. According to the results of this study, feeding cereal grains (especially wheat) can improve nutrient digestibility, nutrient utilization efficiency and milk responses to a greater extent than potato. Potato starch is not superior to grain starch as a readily available energy source for lactating dairy cows and can be used in specific situations (lower cost of potato compared with cereal grains).
References
AOAC, 1990. Official Methods of Analysis. 1, 15th ed. Association of Official Analytical Chemists, Washington, D.C., USA. [ Links ]
Cabrita, A.R.J., Dewhurst, R.J., Abreu, J.M.F. & Fonseca, A.J.M., 2006. Evaluation of the effects of synchronizing the availability of N and energy on rumen function and production responses of dairy cows - A review. Anim. Res. 55, 1-24. [ Links ]
Cabrita, A.R.J., Vale, J.M.P., Bessa, R.J.B., Dewhurst, R.J. & Fonseca, A.J.M., 2009. Effects of dietary starch source and buffers on milk responses and rumen fatty acid biohydrogenation in dairy cows fed maize-based diets. Anim. Feed Sci. Technol. 152, 267-277. [ Links ]
Con, J.W., 1991. Degradation of starch in feed concentrates by enzymes, rumen fluid and rumen enzymes. J. Sci. Food Agric. 54, 23-34. [ Links ]
DeVisser, H., Van Der Togt, P.L. & Tamminga. S., 1990. Structural and non-structural carbohydrates in concentrate supplements of silage-based dairy cow rations: 1. Feed intake and milk production. Neth. J. Agric. Sci. 38, 487-498. [ Links ]
Eriksson, T., Murphy, M., Ciszuk, P. & Burstedt, E., 2004. Nitrogen balance, microbial protein production, and milk production in dairy cows fed fodder beets and potato, or barley. J. Dairy Sci. 87, 1057-1070. [ Links ]
Gozho, G.N. & Mutsvangwa. T., 2008. Influence of carbohydrate source on ruminal fermentation characteristics, performance, and microbial protein synthesis in dairy cows. J. Dairy Sci. 91, 2726-2735. [ Links ]
Griinari, J.M., Dwyer, D.A., McGuier, M.A., Bauman, D.E., Palmquist, D.L. & Nurmela, K.V., 1998. Trans- octadecenoic acids and milk fat depression in lactating dairy cows. J. Dairy Sci. 81, 1251-1261. [ Links ]
Hall, M.B., 2004. Short communication: Effect of carbohydrate fermentation rate on estimates of mass fermented and milk response. J. Dairy Sci. 87, 1455-1456. [ Links ]
Hall, M.B., Larson, C.C. & Wilcox, C.J., 2010. Carbohydrate source and protein degradability alter lactation, ruminal, and blood measures. J. Dairy Sci. 93, 311-322. [ Links ]
Huntington, G.B., 1997. Starch utilization by ruminants: from basics to the bunk. J. Anim. Sci. 5, 852-867. [ Links ]
Jurjanz, S., Colin-Schoellen, O., Gardeur, J.N. & Laurent, F., 1998. Alteration of milk fat by variation in the source and amount of starch in a total mixed diet fed to dairy cows. J. Dairy Sci. 81, 2924-2933. [ Links ]
Karkalas, J., 1985. An improved enzymatic method for the determination of native and modified starch. J. Sci. Food Agric. 36, 1019-1027. [ Links ]
Khorasani, G.R., Okine, E.K. & Kennelly, J.J., 2001. Effects of substituting barley grain with corn on ruminal fermentation characteristics, milk yield and milk composition of Holstein cows. J. Dairy Sci. 84, 2760-2769. [ Links ]
McAllister, T.A., Phillippe, R.C., Rode, L.M. & Cheng, K.J., 1993. Effect of the protein matrix on the digestion of cereal grains by ruminal microorganisms. J. Anim. Sci. 71, 205-212. [ Links ]
Mertens, D.R., 1997. Creating a system for meeting the fiber requirements of dairy cows. J. Dairy Sci. 80, 1463-1481. [ Links ]
Monteils, V., Jurjanz, S., Colin-Schoellen, O., Blanchart, G. & Laurent, F., 2002. Kinetics of ruminal degradation of wheat and potato starches in total mixed rations. J. Anim. Sci. 80, 235-241. [ Links ]
Moren, J.B., 1986. Cereal grains in complete diets for dairy cows: A comparison of rolled barley, wheat and oats and other method of processing oats. Anim. Prod. 43, 27-36. [ Links ]
NRC, 2001. Nutrient Requirments of Dairy Cattle. 7th ed. National Academy of Sciences., Washington, D.C., USA. [ Links ]
Nocek, J.E. & Tamminga, S., 1991. Site of digestion of starch in the gastrointestinal tract of dairy cows and its effects on milk yield and composition. J. Dairy Sci. 74, 3598-3629. [ Links ]
Okine, E. & Kennelly. J.J., 1994. From fiber to starch: The evaluation of the cow. Adv. Dairy Technol. 6, 187-198. [ Links ]
0rskov, E.R., Fraser, R.C., Mason, V.C. & Mann, S.O., 1970. Influence of starch digestion in the large intestine of sheep on caecal fermentation, caecal microflora and faecal nitrogen excretion. Br. J. Nutr. 24, 671-682. [ Links ]
Reynolds, C.K., 2006. Production and metabolic effects of site of starch digestion in dairy cattle. Anim. Feed Sci. Technol. 130, 78-94. [ Links ]
Rode, L.M. & Satter, L.D., 1988. Effect of amount and length of alfalfa hay in diets containing barley or corn on site of digestion and rumen microbial protein synthesis in dairy cows. Can. J. Anim. Sci. 68, 445-454. [ Links ]
SAS, 1999. Statistical Analysis Systems user's guide Version 8.01. SAS Institute Inc., Raleigh, N.C., USA. [ Links ]
Sauvant, D., 1997. Consequences digestives et zootechniques des variations de la vitesse de digestion de lamidon chez les ruminants. Inst. Natl. Rech. Agron. Prod. Anim. 10, 287-300. (in French) [ Links ]
Sauvant, D., Chapoutot, P. & Archimede, H., 1994. La digestion des amidons par les ruminants et ses consequences. Inst. Natl. Rech. Agron. Prod. Anim. 7, 115-124. (in French) [ Links ]
Silveira, C., Oba, M., Beauchemin, K.A. & Helm, J., 2007. Effect of grains differing in expected ruminal fermentability on the productivity of lactating dairy cows. J. Dairy Sci. 90, 2852-2859. [ Links ]
Surber, L.M.M. & Bowman, J.G.L., 1998. Monensin effects on digestion of corn or barley high concentrate diets. J. Anim. Sci. 76, 1945-1954. [ Links ]
Tanaguchi, K., Huntington, G.B. & Glenn, B.P., 1995. Net nutrient flux by visceral tissues of beef steers given abomasal and ruminal infusions of casein and starch. J. Anim. Sci. 73, 236-249. [ Links ]
Tester, R. F., Qi, X. & Kaekalas, J., 2006. Hydrolysis of native starches with amylases. Anim. Feed Sci. Technol. 130, 39-54. [ Links ]
Theurer, C.B., Huber, J.T., Delgado-Elorduy, A. & Wanderley, R., 1999. Invited review: summary of steam- flaking corn or sorghum grain for lactating dairy cows. J. Dairy Sci. 82, 1950-1959. [ Links ]
Van Keulen, J. & Young, B.A., 1977. Evaluation of acid - insoluble ash as a natural marker in ruminant digestibility studies. J. Anim. Sci. 44, 282-287. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3593-3597. [ Links ]
Wang, M., Jiang, J.Z., Tan, L., Tang, S.X., Sun, Z.H. & Han, H.F., 2009. In situ ruminal crude protein and starch degradation of three classes of feedstuffs in goats. J. Appl. Anim. Res. 36, 23-28. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. See: http://creativecommons.org/licenses/by/2.5/za Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science.
# Corresponding author: ffatahnia@yahoo.com">ffatahnia@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}