










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.42 n.2 Pretoria Jan. 2012
SHORT COMMUNICATION
Effect of forage sources and Saccharomyces cerevisiae (Sc 47) on ruminal fermentation parameters
E. GhasemiI; M. KhorvashII, #; A. NikkhahI
IDepartment of Animal Science, University of Tehran, PO Box 3158711167-4111, Karaj, Iran
IIDepartment of Animal Science, Isfahan University of Technology, PO Box 84156, Isfahan, Iran
ABSTRACT
Diet composition has been suggested as a factor that influences the variability of responses when Saccharomyces cerevisiae (SC) is fed to ruminant animals. Diets based on lucerne hay (462 g/kg DM) and maize silage (488 g/kg DM) were fed to determine the effects of 0 or 5 g SC 47 (8x109 cfu/g) on ruminal digestion, fermentation and protozoa population. Ruminal pH, acetate, propionate, degradation rate and effective degradability were significantly affected by the forage sources. The addition of SC caused an increase in degradability of forage neutral detergent fibre (NDF), and tended to enhance degradability of total diet organic matter (OM), and the concentration of propionate and result in a decrease in protozoa numbers at 3 h post feeding. Ruminal crude protein degradation, ammonia-N concentration, acetate : propionate ratio and pH were not elicited by the addition of SC. Although the ruminal environment was significantly affected by the forage sources, no interaction between SC and forage sources occurred for ruminal digestion parameters: pH, ammonia-N and protozoa populations. However, compared with the maize silage, the SC increased the initial degradability (3 h after feeding) of forage NDF (4.6% vs. 1.7%), total diet OM (3.1% vs. 1.0%) and crude protein (CP) (5.5% vs. 0.1%) to higher proportions for lucerne hay. Moreover, acetate concentration was increased on the diet based on maize silage and decreased on the diet based on lucerne hay with supplementation of SC. Although ruminal environments were considerably altered by the forage sources, the SC exhibited a transitory effect (3 h post feeding) without overall improvement on ruminal digestion and fermentation; nevertheless, this observation was more pronounced for lucerne-based diet.
Keywords: Rumen degradability, fermentation, protozoa, forage sources
Yeast culture based on Saccharomyces cerevisiae (SC) has a beneficial effect on the rumen via shifting the ruminal microbial populations, and its effects on animal productivity have been interpreted widely in terms of its action in the rumen (Nagaraja et al., 1997). Increased bacterial count seems to be central to the action of S. cerevisiae associated with increased degradability of forages (Wallace & Newbold, 1992), altered ruminal VFA proportions (Moya et al., 2009), decreased ruminal ammonia concentration, along with increased flow of microbial protein leaving the rumen (Erasmus et al., 1992). However, most studies (Dorea & Jouany, 1998; Lila et al., 2004; Giger-Reverdin et al., 2004) reported that the effect of SC on rumen fermentation and production performance have been marginal and variable. The source and amount of yeast supplemented and interactions among yeast, diet and animal effect (e.g. lactation stage) have been regarded as causes of difference in response to added yeast (Kammalamma et al., 1996; Kung et al., 1997). Newbold et al. (1995) noted that strains of yeast differed in their ability to increase the number of viable bacteria in vitro and in vivo, and responses to yeast were highly variable and apparently influenced by the compositions of the diet. Lila et al. (2004) found variable effects of SC on ruminal fermentation when different substrates were used in vitro, but they did not investigate the interactions between yeast and substrates. In this study, two forage sources were used: lucerne hay as a leguminosae, and maize silage as a gramminaea, which have reverse rate and extent of digestion, to evaluate the influence of S. cerevisiae (Sc 47, Biosaf; Lesaffre Developments, France) on ruminal digestion, fermentation and protozoa population in vivo.
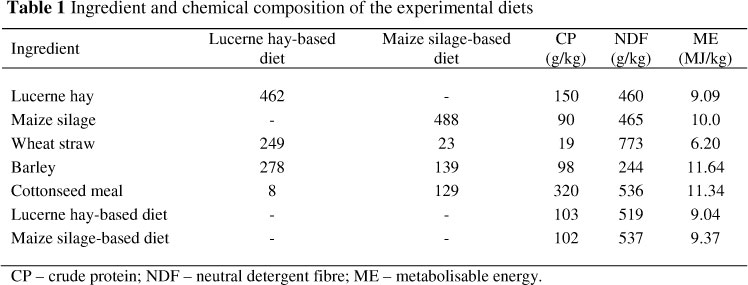
Four mature bulls (two Holstein and two Sistani; 750 kg ± 50 kg) fitted with rumen cannulas were fed the four experimental diets in a 4 χ 4 Latin square design with a 2 χ 2 factorial arrangement of treatments with 21 d in each period (14 d adaptation followed by 7 d data collection). The four treatments, differing in forage sources (lucerne hay or maize silage) and in the absence or presence of 5 g/d of SC, were i) lucerne hay-based diet (46%), no SC; ii) lucerne hay-based diet plus SC; iii) maize silage-based diet (48.8%), no SC; and iv) maize silage-based diet plus SC. Composition of feeds and total diets are given in Table 1. Bulls were restricted fed 10.5 kg DM/d (08:00 and 15:00) as total mixed rations at maintenance level. Saccharomyces cerevisiae (8x109 cfu/g) strain Sc 47 (Biosaf; Lesaffre Developments, France) was top-dressed once per day at 08:00.
Ruminal degradability of forage and total diet was measured using the in situ technique. Samples of forages and ingredients of total diet were ground to pass through a 2 mm screen to determine ruminal degradability of forage NDF and total diet OM and CP. Maize silage had previously been air-dried at room temperature (25 °C) for 48 h. Five-gram samples of lucerne hay, maize silage, the lucerne hay-based diet and the maize silage-based diet were placed in polyester bags (10 cm χ 20 cm; 50 μm pore size) and incubated in duplicate for 3, 6, 12, 24, 48 and 72 h in the rumen. Following incubation, the bags were machine-washed for 14 min, then dried in a forced-air oven at 60 °C for 48 h. Residues from the nylon bags were analysed for NDF (Van Soest et al., 1991), OM and CP (AOAC, 1990).
The degradation kinetics of NDF, OM and CP were estimated by fitting the disappearance values to the following model (McDonald, 1981):
Y= a + b (1-exp-c (t-l)) for t > L where a is the soluble fraction; b the slowly degradable fraction; c the fractional degradation rate constant at which b is degraded; L the lag time (h) and t is the time of incubation (h). Effective degradability (ED) of the feed was determined using ED = a + [bc/(c + k)], where k is the rumen outflow rate, assumed to be 0.03 (fraction/h), describing retention time at maintenance level.
Ruminal fluid samples were collected at 3 h after feeding and strained through two layers of cheesecloth to measure ruminal pH and concentration of ammonia-N and VFA. Ruminal pH was recorded immediately after sampling using a glass electrode. Approximately 0.8 mL of sulphuric acid (0.098 M) was added to a 50 mL of ruminal fluid for ammonia-N measurement by a microdiffusion method (Conway, 1950). For VFA determination by gas chromatography (model PU 4410, Philips, England), 1 mL of 2.5 M meta-phosphoric acid was added to a 4 mL of rumen fluid (Ottenstein & Bartley, 1971). Ruminal contents were collected at 3 h post feeding to enumerate total protozoa, entodiniomoroph and holotrich organisms.
Each sample was subsampled, and then 10 mL of ruminal contents was preserved by adding 10 mL of 50% formalin (Dehority, 1984).
Data on ruminal parameters as a 4 χ 4 Latin square with 2 χ 2 factorial arrangement of treatments were analysed using the GLM procedure of SAS Institute (SAS, 2001).
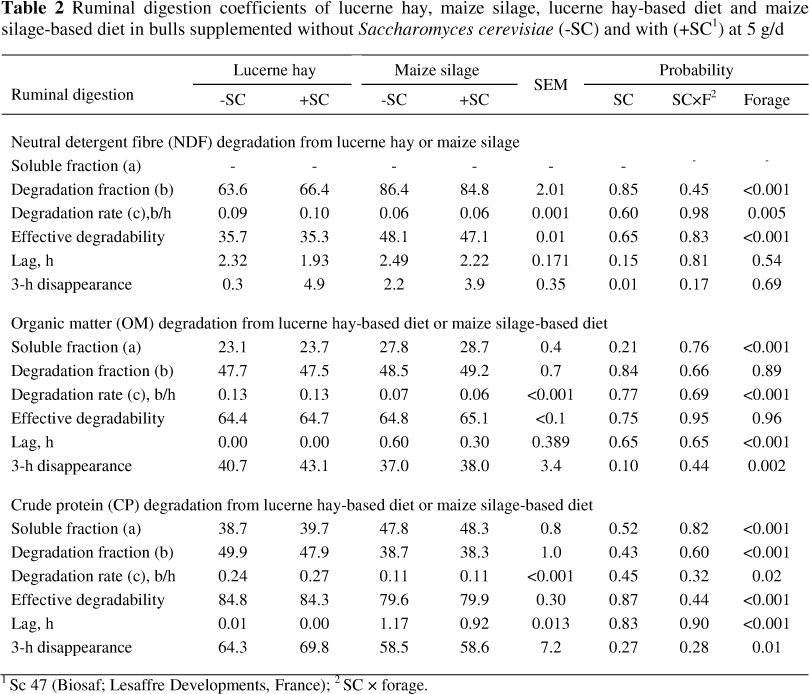
Ruminal digestion rate of lucerne hay NDF was significantly greater than maize silage (9.9%/h vs. 6.1%/h), whereas the extent of digestibility was the converse (36% vs. 48%; Table 2). In situ degradation of forages NDF (P <0.01) and total diets OM (P <0.10) were higher at initial time (3 h incubation) of incubation when SC was added to the diets. This increase in initial degradability tended to cause a slight decrease in lag time of forage NDF (P <0.15) with SC addition. No other significant effect of SC was found in degradability parameters (a, b, c or ED). No interaction was observed between SC and forage sources on ruminal digestibility of NDF, OM or CP. However, SC exerted a larger influence, although not statistically, on initial digestibility (3 h incubation) of forage NDF (4.6% vs. 1.7 %), total diet OM (3.1% vs. 1.0%) and CP (5.5% vs. 0.1%) and decrease in lag time of forage NDF (0.39 vs. 0.27 h) for lucerne hay than maize silage, respectively.
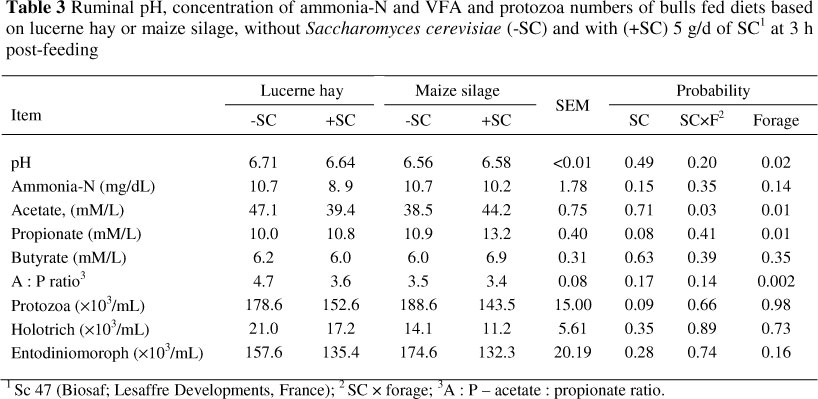
Ruminal pH, acetate and acetate to propionate ratio were higher for diet based on lucerne hay (P <0.05; Table 3). Ruminal pH was not affected by SC (P >0.05). The SC supplementation did not modify the concentration of the rumen ammonia-N, acetate and butyrate and the ratio of acetate to propionate but tended to increase propionate concentration (P <0.08) at 3 h post feeding. There were no interactions between SC and forage sources on ruminal pH, NH3 and individual VFA concentrations and protozoa counts (P >0.05).
However, interaction was significant (P <0.05) for molar concentration of acetate. The addition of SC to the diet based on lucerne hay resulted in a lower acetate concentration, while in the diet based on maize silage, that was lower when no yeast was added (P <0.05). Total counts of protozoa decreased (P <0.10) at 3 h post feeding when SC was supplemented (Table 3).
Increased initial degradability of forages in conjunction with decrease in lag time corresponds with the two basic mechanisms involved in the stimulation of ruminal bacterial growth with the addition of yeast in the rumen. One mode of yeast action reported by Newbold et al. (1996) is the respiratory activity that scavenges O2 which is toxic to anaerobic bacteria and causes inhibition of adhesion of cellulolytic bacteria to cellulose, and this peak in O2 concentration occurs at approximately the time of feeding (initial time). The other is that yeast-derived small peptides and other nutrients reduce the time required to initiate growth (lag) of the predominant ruminal cellulolytic bacteria (Girard, 1996; Callaway & Martin, 1997). The reason that this increase in initial degradability did not continue, even with the consistent rise in total bacteria counts, may support the hypothesis of the Cornell Net Carbohydrate and Protein System (CNCPS), which assumes that the rumen operates as a substrate-limited, enzyme (microbial mass) excess system (Fox et al., 2004). Therefore, little benefit in the extent of digestion may be associated with increased ruminal bacterial numbers stimulated by SC.
Yeast decreased total counts of protozoa at 3 h post feeding, but did not modify counts of holotrich and entodiniomoroph. Kumar et al. (1994) and Newbold et al. (1995) reported that S. cerevisiae did not modify protozoa population in buffalo or in sheep, respectively. The various strains of SC differing in their hydrolyzing ability may partly account for these contradictory results. A wide range of mechanisms by which S. cerevisiae might stimulate bacterial counts has been proposed (Wallace & Newbold, 1992). The decreases in numbers of protozoa suggest another possibility, because they are the primary cause of bacterial breakdown in mixed populations and may partly account for the increased bacterial numbers.
The yeast supplement used in this study altered rumen fermentation parameters, in particular for lucerne hay at 3 h post feeding. However, no overall enhancement of digestion and interactions between SC and forage sources were observed, despite ruminal environment change caused by the forage sources.
Acknowledgements
Financial support of this study was provided by the Department of Animal Science, University of Tehran. The authors acknowledge B.A. Dehority and M.T. Yokoyama for their guidance in counting protozoa and Makian Daro Co. for provision of yeast.
References
AOAC, 1990. Official Methods of Analysis, Vol. I., 15th ed. Association of Official Analytical Chemists, Arlington, Virginia. [ Links ]
Callaway, E.S. & Martin, S.A., 1997. Effects of Saccharomyces cerevisiae culture on ruminal bacteria that utilize lactate and digest cellulose. J. Dairy Sci. 80, 2035-2044. [ Links ]
Conway, E.J., 1950. Microdiffusion Analysis and Volumetric Error. 2th ed. Crosby Lockwood & Son, London, U.K. [ Links ]
Dehority, B.A., 1984. Evaluation of sampling and fixation procedures used for counting rumen protozoa. Appl. Environ. Microbiol. 48, 182-185. [ Links ]
Dorea, M. & Jouany, J.P., 1998. Effect of Saccharomyces cerevisiae culture on nutrient digestion in lactating dairy cows. J. Dairy Sci. 81, 3214-3221. [ Links ]
Erasmus, L.J., Botha, P.M. & Kistner, A., 1992. Effect of yeast culture supplement on production, rumen fermentation, and duodenal nitrogen flow in dairy cows. J. Dairy Sci. 75, 3056-3065. [ Links ]
Fox, D.G., Tedeschi, L.O., Tylutki, T.P., Russell, J.B., Van Amburgh, M.E., Chase, L.E., Pell, A.N. & Overton, T.R., 2004. The Cornell Net Carbohydrate and Protein System Model for evaluating herd nutrition and nutrient excretion. Anim. Feed Sci. Technol. 112, 29-78. [ Links ]
Giger-Reverdin, S., Sauvant, D., Tessier, J., Bertin, G. & Morand-Fehr, P., 2004. Effect of live yeast culture supplementation on rumen fermentation in lactating dairy goats. S. Afr. J. Anim. Sci. 2004, 34 (Suppl.1), 59-61. [ Links ]
Girard, I.D., 1996. Characterization of stimulatory activities from Saccharomyces cerevisiae on the growth and activities of ruminal bacteria. Ph.D. dissertation. University of Kentucky, Lexington, USA. [ Links ]
Kammalamma, U.K. & Krishnappa, P., 1996. Effect of feeding yeast culture (Yea-sacc1026) on rumen fermentation in vitro and production performance in crossbred dairy cows. Anim. Feed Sci. Technol. 57, 247-256 [ Links ]
Kumar, U., Sareen, V.K. & Singh, S., 1994. Effect of Saccharomyces cerevisiae yeast culture supplement on ruminal metabolism in buffalo calves given a high concentrate diet. Anim.Prod. 59, 209-215. [ Links ]
Kung Jr, L., Kreck, E.M. & Tung, R.S., 1997. Effects of a live yeast culture and enzymes on in vitro ruminal fermentation and milk production of dairy cows. J. Dairy Sci. 80, 2045-2051. [ Links ]
Lila, Z.A., Mohammed, N., Yasui, T., Kurokawa, Y., Kanda, S. & Itabashi, H., 2004. Effects of a twin strain of Saccharomyces cerevisiae live cells on mixed ruminal microorganism fermentation in vitro. J. Anim. Sci. 82, 1847-1854. [ Links ]
McDonald, I., 1981. A revised model for the estimation of protein degradability in the rumen. J. Agric. Sci. 96, 251-252. [ Links ]
Moya, D., Calsamiglia, S., Ferret, A., Blanch, M., Fandino, J.I., Castillejos, L. & Yoon, I., 2009. Effects of dietary changes and yeast culture (Saccharomyces cerevisiae) on rumen microbial fermentation of Holstein heifers. J. Anim. Sci. 87, 2874-2881. [ Links ]
Nagaraja, T.G., Newbold, C.J., Van Nevel, C.J. & Demeyer, D.I., 1997. Manipulation of ruminal fermentation. In: The Rumen Microbial Ecosystem (2nd ed.). Eds Hobson, P.M. & Stewart, C.W., Blackie Academic and Professional, London, England. p. 521. [ Links ]
Newbold, C.J., Wallace, R.J., Chen, X.B. & McIntosh, F.M., 1995. Different strains of Saccharomyces cerevisiae differ in their effects on ruminal bacterial number in vitro and in sheep. J. Anim. Sci. 73, 1811-1818. [ Links ]
Newbold, C.J., Wallace, R.J. & McIntosh, F.M., 1996. Mode of action of the yeast Saccharomyces cerevisiae as a feed additive for ruminants. Br. J. Nutr. 76, 249-261. [ Links ]
Ottenstein, D.M. & Bartley, D.A., 1971. Determination of rumen VFA. Anal. Chem. 43, 952-955. [ Links ]
SAS, 2001. The Statistical Analysis System for Windows, Release 8.2. SAS Institute Inc., Cary, N.C., USA. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3592. [ Links ]
Wallace, R.J. & Newbold, C.J., 1992. Probiotics for ruminants. In: Probiotics: The Scientific Basis. Ed. Fuller, R., Chapman and Hall, London, UK. pp. 317-353. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. S ee: http://creativecommons.org/licenses/by/2.5/za Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science.
# Corresponding author:khorvash@cc.iut.ac.ir


{kind=link}
{kind=link}
{kind=link}