










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.42 no.1 Pretoria ene. 2012
Assessment of inbreeding depression for functional herd life in the South African Jersey breed based on level and rate of inbreeding
J. du ToitI, II, #; J.B. van WykII; A. MaiwasheIII
IAgricultural Research Council, Animal Production Institute, Private Bag X5013, Stellenbosch, 7599, South Africa
IIDepartment of Animal, Wildlife and Grassland Sciences, University of the Free State, P.O. Box 339, Bloemfontein, 9300, South Africa
IIIAgricultural Research Council, Animal Production Institute, Private Bag X2, Irene, 0062, South Africa
ABSTRACT
The objective of this study was to investigate the effect of inbreeding depression on functional herd life in the South African Jersey population based on individual level and rate of inbreeding. A pedigree file of the South African Jersey breed (n = 912 638) was obtained from the Integrated Registration and Genetic Information System (INTERGIS). The data included registered, grade and imported animals. The percentages of animals in the pedigree file with two, one and zero parents unknown were 22%, 18% and 60%, respectively. The inbreeding coefficient for each animal (Fi) and the rate of individual inbreeding (DFi) as an alternative measure of inbreeding that is adjusted for the depth of known pedigree were calculated. The effect of inbreeding on functional herd life in each of the first three lactations was estimated, using a single-trait sire model on data collected from 1985 to 2003. Three analyses for survival in each of the first three lactations were conducted. In the first analysis, in addition to fixed and random effects, an individual inbreeding coefficient (Fi) was fitted as a linear covariate. In the second analysis, the inbreeding coefficient was included as a discrete variable with the following classes of inbreeding: 0 < F ≤ 3.125, 3.125 < F ≤ 6.25, 6.25 < F ≤ 12.5 and F > 12.5. In the third analysis, the individual rate of inbreeding (DFi) was included in the model as a linear covariate. The level of inbreeding in the SA Jersey population showed a gradual increase for the period 1985 to 1994, while the period 1995 to 2003 showed a rapid increase. The current mean level of inbreeding (for the year 2010) is 4.85% with a minimum and maximum of 0 and 31.34%, respectively. The rate of inbreeding showed a gradual increase from 0.36% to 0.43% between 1985 and 2003. The average rate of inbreeding is currently (for the year 2010) at 0.55%. There was a significant unfavourable relationship between inbreeding and functional herd life in the first and second lactations. The effect of inbreeding was more pronounced in the second lactation for both measures of inbreeding. Based on the current level of inbreeding, the reduction in functional herd life in the first lactation can be estimated as 0.68%. The corresponding estimate for the second lactation is 1.70%. The results from the current study indicate that the current level or rate of inbreeding has reached levels that are detrimental to functional herd life. Therefore, individual inbreeding coefficients should be considered in addition to genetic merit when breeding decisions are made by Jersey breeders.
Keywords: Inbreeding, longevity, survival, dairy cattle
Introduction
Inbreeding will occur in any population of finite size unless specific measures are taken to avoid the mating of related individuals. Therefore, genetic improvement programmes should balance genetic gain with increases in inbreeding levels (Miglior et al, 1992). Increased inbreeding is usually considered undesirable because it leads to a decrease in genetic variation within a population, and to a reduction in performance in traits associated with fitness (e.g. health, fertility, survival). While the impact of inbreeding in large populations is negligible, its effect in a typical livestock population, where selective breeding is practised, cannot be ignored (Maiwashe et at., 2006). In practice, an increase in the coefficient of relationship among individuals results in increased difficulty in the selection of unrelated mates (Thompson et at., 2000a; b). It is also important to maintain genetic diversity within a breed to ensure that future animals can respond to selection and changes in the environment. The net effect of inbreeding in a selection programme will depend on the magnitude of the selection response relative to the possible depression and the rate of accumulation of inbreeding (Van Wyk et at., 2009).
Research shows that there has been a general increase in the level of inbreeding in dairy cattle populations in various countries. In the Holstein population in the United States, Young & Seykora (1996) obtained rates of inbreeding of 0.11% per year for the period 1960 to 1990. Thompson et at. (2000a; b) reported average rates of inbreeding of 0.04% in the 1970s, with an average yearly increase of 0.12% in the 1980s. It continued to increase by 0.20% per year from 1990 to 1998 for the same population. The Canadian Dairy Network reported that the rate of inbreeding increased by 0.25% per year for the period 1990 to 2000 (http://www.cdn.ca). Kearney et at. (2004) reported a rate of inbreeding of 0.03% per year from 1968 to 1991 and an increased rate of 0.17% per year for the period 1992 to 2002 in the United Kingdom Holstein population.
In a study on registered South African Ayrshire, Guernsey, Holstein and Jersey breeds, Maiwashe et at. (2006) reported that for most of the period 1960 to 1975, at least 70% of the population were foundation animals. A rapid increase in the proportion of animals with both parents known occurred in the period 1975 to 1985. In 2003 the proportion of animals with both parents known increased to 90% with an average level of inbreeding of 3.05% and a corresponding maximum inbreeding coefficient of 34.6% for the registered Jersey breed. In the current study, consisting of registered, grade and imported Jersey animals, only 60% of both parents are known.
The detrimental impact of inbreeding on performance traits, especially those related to fitness, has been widely recognized and is a result of the reduction in heterozygosity as inbreeding accumulates (Falconer & Mackay, 1996). Inbreeding depression is a reduction of mean phenotypic values in animals that have been inbred. If selection and inbreeding are applied simultaneously, deleterious alleles can possibly have their frequency diminished in the population (Norberg & Sorensen, 2007). The effect depends not only on the actual level of inbreeding, but also on the rate at which inbreeding is increasing. Van Wyk et at. (2009) suggested that slow inbreeding (1.53% per generation over 19 generations) allows natural selection to operate and to remove the less adapted animals. Less inbreeding depression would then be expected among the individuals who accumulated the inbreeding over a larger number of generations for any given level of inbreeding. For example, animals with the same individual inbreeding coefficients could have different inbreeding depression effects owing to differences in the number of generations in their own pedigree. González-Recio et at. (2007) developed an alternative method of fitting inbreeding coefficients that accounted for the depth of known pedigree. This coefficient corrects the cumulative inbreeding coefficient regarding the pedigree depth of the individual and is an indicator of the increment in inbreeding, regardless of the number of generations known in its pedigree (González-Recio et at., 2007).
While the detrimental effect of inbreeding on production traits in dairy cattle is well documented, little information is available on the effect of inbreeding on longevity of cows (Thompson et at., 2000a; Caraviello et at., 2003; Sewalem et at., 2006). Smith et at. (1998) showed that an increase of 1 per cent in inbreeding results in a decrease of 13.1 days of productive life. Cows in the Canadian population that are 10% inbred versus 5% had reduced longevity of 65 days (CDN, 2008). Mostert (2011) reported that every 1% increase in inbreeding in the South African Jersey breed is associated with a decrease of 15.42 kg, 0.64 kg and 0.59 kg in 305-day milk, butterfat and protein production, respectively. Rokouei et at. (2010) reported that cows with high levels of inbreeding were at higher relative risk of being culled. The objective of this study was to investigate inbreeding depression on functional herd life in the South African Jersey population based on individual level and rate of inbreeding.
Materials and Methods
A pedigree file of the South African Jersey breed (n = 912 638) was obtained from INTERGIS. The data were checked for integrity (e.g. animals were ordered chronologically and no animals appeared as both sire and dam) using the Animal Breeder's Tool Kit (ABTK, Golden et at., 1992). The data included registered, grade and imported animals. Pedigree information of imported ancestors was retrieved from the INTERBULL database. These pedigrees were traced as far back as 1935. The percentages of animals in the pedigree file with two, one and zero parents unknown were 22%, 18% and 60%, respectively
The inbreeding coefficient for each animal (Fi) in the pedigree was calculated using the ainv tool of the ABTK. The ainv tool implements Meuwissen & Luo's (1992) algorithm to compute the inbreeding coefficients. Individual inbreeding coefficients were used to compute the individual rate of inbreeding (DFi) according to the methodology described by Gonzalez-Recio et al. (2007) and modified by Gutierrez et al. (2009). The DFi is an alternative measure of inbreeding, which is adjusted for the depth of known pedigree. Therefore, the individual rate of inbreeding makes it possible to distinguish between two animals with the same inbreeding coefficient, but different numbers of generations in which inbreeding was accumulated (González-Recio et al., 2007; Gutierrez et al., 2009). The individual rate of inbreeding was calculated as
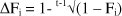
where t is the number of known equivalent generations for the ith individual. The t was calculated using the ENDOG v4.3 computer program (Gutiérrez & Goyache, 2005).
The effect of inbreeding on functional herd life in each of the first three lactations was estimated using a single-trait sire model. The general form of the model fitted was as follows:

where y is a vector of observations for productive herd life in each lactation, b is a vector of fixed effects of herd-year, registry status x herd size change x season of calving (rhs), age at calving (linear and quadratic), protein within rhs, protein and fat yield deviations (linear, quadratic and cubic), s is a vector of random sire effects, e is a vector of random residuals. The X and Z are the incidence matrices relating fixed and random sire effects to observations, respectively. The random effects were assumed to be distributed as follows:
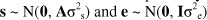
where σ2s and σ2e are the sire and residual variances, respectively. The A and I are the numerator relationship and the identity matrices, respectively. The sire and residual variances used in this study were calculated using heritability estimates reported by Du Toit et al. (2009).
Three analyses for functional herd life in each of the first three lactations were carried out using variations of Equation 1. In the first analysis, an individual inbreeding coefficient (Fi) was fitted as a linear covariate in addition to the fixed and random effects in Equation 1. In the second analysis, the inbreeding coefficient was included as a discrete variable with the following classes of inbreeding (%): 0 < F ≤ 3.125, 3.125 < F ≤ 6.25, 6.25 < F ≤ 12.5 and F > 12.5, following the approach of Sewalem et al. (2006). In the third analysis, the individual rate of inbreeding (DFi) was added to the model as a linear covariate. The individual rate of inbreeding was considered in the current study, because it automatically accounts for differences in pedigree depth.
The MTDFREML software package was used to estimate the effect of inbreeding on functional herd life and the corresponding approximate standard errors (Boldman et al., 1995).
Results and Discussion
A summary of the data with details regarding the numbers of animals, mean level of inbreeding (%) by birth year cohort, and the proportion of animals for each class of inbreeding is shown in Table 1. In the 1985 - 1989 birth year cohort 71.0% of the population were not inbred, while only 15.6% of the animals were not inbred in the 2000 - 2003 cohort. There was a gradual increase in the level of inbreeding for the period 1985 to 1994, while the period 1995 to 2003 showed a rapid increase. The average level of inbreeding for the period 2000 to 2003 was 2.91%. This occurrence might be associated with superior sires being used more extensively, or possibly the incidence of more complete pedigree information.
Thompson et al. (2000a; b) reported average inbreeding coefficients of 4.2% and 4.6% for the American Holstein and Jersey populations, respectively. Sorensen et al. (2005) reported average inbreeding levels of 3.9% and 3.4% in 2003 in the Danish Holstein and Jersey breeds, respectively.
More recently, a slightly lower inbreeding level of 1.9% was reported for the Israeli Holstein population (Weller & Ezra, 2005). Among the five major breeds in Canada, all have an average inbreeding level of between 5.50% (Ayrshire) and 6.64% (Guernsey) for registered animals born in 2009 (CDN, 2010). The Ayrshire and Jersey breeds have controlled the rate of increase in the average inbreeding level best since 2000, at -0.04% and +0.07% per year respectively, with the Holstein at +0.08% per year.
Similar results were obtained in a study by Maiwashe et al. (2006). The Jersey breed exhibited the highest annual rate of inbreeding (0.07%), followed by the Holstein (0.06%), and the Ayrshire and the Guernsey breeds, both with (0.05%) Differences in the levels of inbreeding in the various populations could be due to different base years.
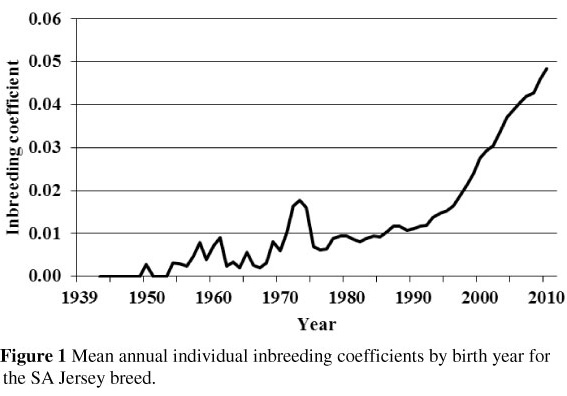
Figure 1 shows the mean annual inbreeding coefficients for Jersey animals born between 1939 and 2010. The average level of inbreeding for the Jersey breed is currently (2010) at 4.85% with a minimum and maximum of 0 and 31.34%, respectively. The rapid increase in the level of inbreeding in the last 18 years is worth noting.
Thompson et al. (2000a; b) reported average inbreeding coefficients of 4.2% and 4.6% for the American Holstein and Jersey populations, respectively. Sorensen et al. (2005) reported average inbreeding levels of 3.9% and 3.4% in 2003 in the Danish Holstein and Jersey breeds, respectively.
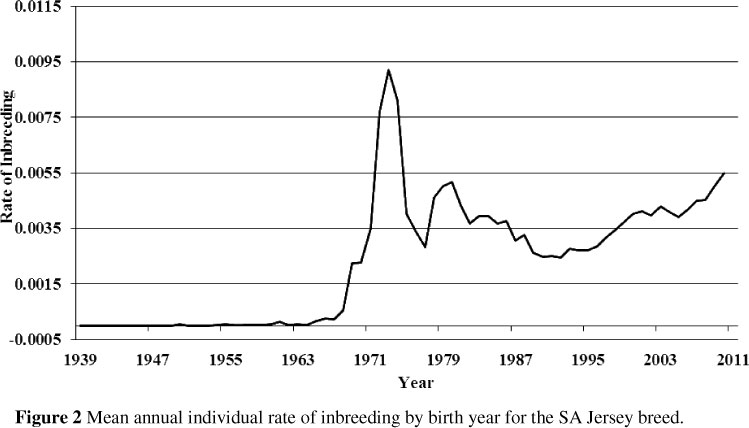
The trend of the annual individual rate of inbreeding for SA Jersey animals born between 1940 and 2010 is presented in Figure 2. The sharp increase in the rate of inbreeding to approximately 0.92% per year in the early 1970s is noticeable. According to Van Niekerk (personal communication, Jersey SA) this incidence might be the result of suspension of importing semen during that period. In general, estimates of the rate of inbreeding obtained in this study are still lower than the critical level of 0.5% per year suggested for animal breeding programmes (Nicholas, 1989). However, estimates of the rate of inbreeding of the SA Jersey breed have escalated since 2004 and are currently (2010) alarmingly high (0.55%).
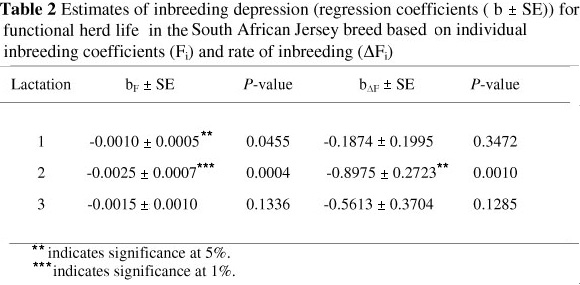
The results in Table 2 indicate that there was a significant (P <0.05) negative relationship between inbreeding and functional herd life for the SA Jersey breed in the first and second lactation, resulting in a decreased survival among more inbred cows. The effect of inbreeding was more pronounced in the second lactation for both measures of inbreeding. A negative but non-significant (P >0.05) effect of inbreeding was observed in the third lactation. This result could be owing to the culling of low-producing highly inbred cows during the first two lactations.
The effect of inbreeding was expressed as a percentage of the mean to allow comparison with similar studies. The results of the effect of inbreeding on functional herd life of the SA Jersey breed expressed as a percentage of the mean are presented in Table 3. The mean survival in each lactation is also presented. The effect of inbreeding, expressed as a percentage of the mean, was -0.14% for the first lactation, indicating that a 1 per cent increase in inbreeding is associated with a reduction in mean survival of 0.14%. The current mean level of inbreeding (for 2010) in this study is 4.85%. Based on this, the reduction in survival in the first lactation can be estimated as 0.68%. The corresponding estimate for the second lactation is 1.70%.

Miglior et al. (1992) also observed a significant reduction of -0.91% of the mean on productive life associated with a percentage increase in inbreeding in the Canadian Jersey population. Thompson et al. (2000a; b) reported a decline in survival as the level of inbreeding increases. Inconsistencies in the magnitude of estimates of the effect of inbreeding across studies could be because of the different rates and levels of inbreeding in different populations. For example, the level of inbreeding in the Canadian Jersey population in 2009 was higher than the current level of inbreeding in the South African Jersey population (5.92% vs. 4.85%).
The two measures of inbreeding (Fi and ΔFi) used to estimate regression coefficients in the current analyses both detected a decline in functional herd life with increased inbreeding. Although the individual inbreeding coefficient is the commonest parameter used in most inbreeding studies, it cannot account for the depth of known pedigree. The rate of inbreeding (ΔFi) is an alternative parameter that relates the increase of the individual inbreeding of animals in the population, accounting for the amount of known pedigree (González-Recio et al., 2007). This property is an advantage of ΔFi over Fi when pedigree is incomplete. The higher level of inbreeding depression of functional herd life obtained when using ΔFi in comparison with Fi (Table 2) is in accordance with the results of González-Recio et al. (2007) in female fertility and calving ease in Spanish dairy cattle. From the results of a study with Spanish horses, Gómez et al. (2009) recommended the use of the individual increase in inbreeding coefficient (ΔFi) instead of the individual inbreeding coefficient (Fi) owing to the better fit with data and the special property whereby individual inbreeding coefficients are adjusted for the known pedigree depth.
The effect of inbreeding was also estimated, considering inbreeding as a class variable. This analysis was conducted to gain better understanding of the dynamics of the effect of inbreeding as inbreeding increases. The results presented in Table 4 are in general agreement with the results from the regression analysis (Table 2).
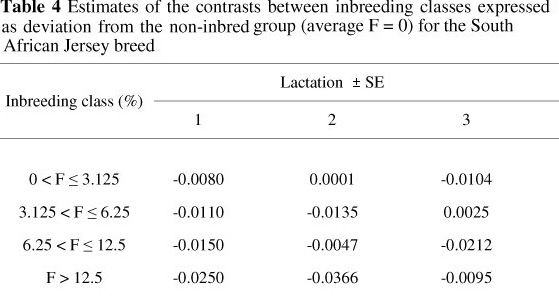
When inbreeding was treated as a class variable, animals with low inbreeding coefficients of between 0 and 3.13%, were not significantly (P >0.05) affected, compared with non-inbred animals (Fi = 0). However, as inbreeding increased from 3.13% to 6.25% and higher, a more pronounced decline in functional herd life was observed in all lactations.
As shown in Table 4, a negative trend was observed as inbreeding increased in lactation 1. In contrast, no clear pattern was observed with lactations 2 and 3.
Conclusions
The effect of inbreeding on functional herd life was assessed for South African Jersey cattle using two measures of inbreeding. Significant negative effects of inbreeding on functional herd life were observed in the first and second lactation. However, no significant association was observed between inbreeding and functional herd life in the third lactation. The results from the current study indicate that the current levels or rate of inbreeding have reached the point that they are detrimental to functional herd life. Therefore, individual inbreeding coefficients should be considered when breeding decisions are made by the Jersey breeders.
Acknowledgments
This study formed part of the first author's doctoral thesis, which benefited from funding from the South African National Research Foundation. Assistance with the pedigree file from B.E. Mostert is greatly appreciated.
References
Boldman, K.G., Kriese, L.A., Van Vleck, L.D., Van Tassell, C.P. & Kachman, S.D., 1995. A manual for use of MTDFREML. A set of programs to obtain estimates of variances and covariances. [Draft], USDA-ARS, Lincoln, N.E., USA. [ Links ]
Canadian Dairy Network (CDN), 2008. Subject: Quantifying Inbreeding Depression. http://www.cdn.ca. Accessed March, 2008. [ Links ]
Canadian Dairy Network (CDN), 2010. Subject: Inbreeding Update. http://www.cdn.ca. Accessed: Aug. 2010. [ Links ]
Caraviello, D.Z., Weigel, K.A. & Gianola, D., 2003. Analysis of the relationship between type traits, inbreeding and functional survival in Jersey cattle using a Weibull proportional hazards model. J. Dairy Sci. 86, 2984-2989. [ Links ]
Du Toit, J., Van Wyk, J.B. & Maiwashe, A., 2009. Genetic parameter estimates for functional herd life for the South African Jersey breed using a multiple trait linear model. S. Afr. J. Anim. Sci. 39, 40-44. [ Links ]
Falconer, D.S. & Mackay, T.F.C., 1996. Introduction to Quantitative genetics. Longman Group, Essex, UK. [ Links ]
Golden, B.L., Snelling, W.M. & Mallinckrodt, C.H., 1992. Animal breeder's tool kit user's guide and reference manual. Colorado State Univ. Agric. Exp. Sta. Tech. Bull. LTB 92-2. [ Links ]
Gómez, M.D., Valera, M., Molina, A., Gutierrez, J.P. & Goyache, F., 2009. Assessment of inbreeding depression for body measurements in Spanish Purebred (Andalusian) horses. Livest. Sci. 122, 149155. [ Links ]
González-Recio, O., López de Maturana, E. & Gutierrez, J.P., 2007. Inbreeding depression on female fertility and calving ease in Spanish dairy cattle. J. Dairy Sci. 90, 744-5752. [ Links ]
Gutierrez, J.P. & Goyache, F., 2005. A note on ENDOG: a computer program for analysing pedigree information. J. Anim. Breed. Genet. 122, 172-176. [ Links ]
Gutiérrez, J.P., Cervantes, I. & Goyache, F., 2009. Improving the estimation of realized effective population sizes in farm animals. J. Anim. Breed. Genet. 126, 327-332 [ Links ]
Kearney, J.F., Wall, E., Villanueva, B. & Coffey, M.P., 2004. Inbreeding trends and application of optimized selection in the UK Holstein population. J. Dairy Sci. 87, 3503-3509. [ Links ]
Maiwashe, A., Nephawe, K.A., Van der Westhuizen, R.R., Mostert, B.E. & Theron, H.E., 2006. Rate of inbreeding and effective population size in four major South African dairy cattle breeds. S. Afr. J. Anim. Sci. 36, 50-57. [ Links ]
Meuwissen, T.H.E. & Luo, Z., 1992. Computing inbreeding coefficients in large populations. Genet. Sel. Evol. 24, 305-313. [ Links ]
Miglior, F., Szkotnicki, B. & Burnside, E.B., 1992. Analysis of levels of inbreeding and inbreeding depression in Jersey cattle. J. Dairy Sci. 75, 1112-1118. [ Links ]
Mostert, B.E., 2011. Inbreeding facts of the South African Jersey breed. SA Jersey J. 60 (1), 40-45. [ Links ]
Nicholas, F.W., 1989. Evolution and Animal Breeding. Eds Hill, W.G. & Mackay, T.F.C., CAB Intl, Wallingford, UK. [ Links ]
Norberg, E. & Sorensen, A.C., 2007. Inbreeding trend and inbreeding depression in the Danish populations of Texel, Shropshire and Oxford Down. J. Anim. Sci. 85, 299-304. [ Links ]
Rokouei, M., Vaez Torshizi, R., Moradi Shahrbabak, M., Sargolzaei, M. & S0rensen, C., 2010. Monitoring inbreeding trends and inbreeding depression for economically important traits of Holstein cattle in Iran. J. Dairy Sci. 93, 3294-3302 [ Links ]
Sewalem, A., Kistemaker, G.J., Miglior, F. & Van Doormaal, B.J., 2006. Analysis of inbreeding and its relationship with functional longevity in Canadian dairy cattle. J. Dairy Sci. 89, 2210-2216. Smith, L.A., Cassell, B.G. & Pearson, R.E., 1998. The effects of inbreeding on the lifetime performance of dairy cattle. J. Dairy Sci. 81, 2729-2737. [ Links ] [ Links ]
Sorensen, A.C., Sorensen, M.K. & Berg, P., 2005. Inbreeding in Danish dairy cattle populations. J. Dairy Sci. 88, 1865-1872. [ Links ]
Thompson, J.R., Everett, R.W. & Hammerschmidt, N.L., 2000a. Effects of inbreeding on production and survival in Holsteins. J. Dairy Sci. 83, 1856-1864. Thompson, J.R., Everett, R.W. & Wolfe, C.W., 2000b. Effects of inbreeding on production and survival in Jerseys. J. Dairy Sci. 83, 2131-2138. [ Links ] [ Links ]
Van Wyk, J.B., Fair, M.D. & Cloete, S.W.P., 2009. Case study: The effect of inbreeding on the production and reproduction traits in the Elsenburg Dormer sheep stud. Livest. Sci. 120, 218-224. [ Links ]
Weller, J.L. & Ezra, E., 2005. Analysis of inbreeding in the Israeli Holstein dairy cattle population. Proc. 2005 Interbull meeting, Uppsala, Sweden, June 2-4, 85-87. [ Links ]
Young, C.W. & Seykora, A.J., 1996. Estimates of inbreeding and relationship among registered Holstein females in the United States. J. Dairy Sci. 79, 502-508. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence.
See: http://creativecommons.org/licenses/by/2.5/za Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science.
# Corresponding author: dtoitj@arc.agric.za


{kind=link}
{kind=link}