










Servicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares en Google
Similares en Google
Compartir

 Permalink
PermalinkSouth African Journal of Animal Science
versión On-line ISSN 2221-4062
versión impresa ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 no.4 Pretoria ene. 2011
Effect of concentrate supplementation during early lactation on plasma IGF-I concentrations and reproductive performance of dairy cows
F.Y. ObeseI,#; T.E. StirlingIII; R.C. StockdaleIV; K.L. MacmillanIII;, A.R. EganII; S. HumphrysV
IDepartment of Animal Science, College of Agriculture and Consumer Sciences, University of Ghana, P.O. Box LG 226, Legon, Ghana
IISchool of Agriculture and Food Systems, University of Melbourne, VIC. 3031, Australia
IIISchool of Veterinary Science, University of Melbourne, Werribee, VIC. 3030, Australia
IVDepartment of Primary Industries, 120 Cooma Road, Kyabram, VIC. 3620, Australia
VPrimegro (Pty) Ltd., Thebarton, SA. 5001, Australia
ABSTRACT
An experiment was conducted to assess the effects of body condition score (BCS) at calving and level of feed supplementation during early lactation on the Insulin-like growth Factor-I (IGF-I) and reproductive performance in 72 Holstein cows in a pasture-based system following seasonally-synchronized calving. At the start of the seasonal breeding, plasma IGF-I concentrations in cows that had recommenced ovarian activity with detected oestrus were compared with those in anoestrus. The cows (n = 72) were managed over a period of five months to calve in a BCS of 4, 5 or 6 - using a scale from 1 (thin) to 8 (obese). The cows were maintained on pastures and supplemented during early lactation with either 1 or 6 kg grain per day. The BCS at calving did not affect plasma IGF-I concentrations. The plasma IGF-I concentration at week 10 was elevated at the higher level of grain supplementation (87.8 vs. 72.1 ng/mL). Higher conception rate following the first insemination of cows calving with a BCS of 6 (71% vs. 46% and 37% for a BCS of 4 and 5, respectively) was associated with more cows in that group conceiving during the first three weeks of the artificial insemination (AI) programme (75% vs. 46% and 46% for a BCS of 4 and 5, respectively). At weeks 0 and 10, cows that commenced ovarian activity by the start of the intensive AI programme, recorded higher plasma IGF-I concentrations than anoestrous cows. These findings indicate that diet can affect the plasma IGF-I levels, which in turn may be involved in affecting the resumption of cyclic ovarian activity and conception in dairy cows.
Keywords: BCS, conception, dairy cows, IGF-I, postpartum period
Introduction
The nutritional status before calving and during the postpartum period, as well as body condition score (BCS) at calving have been shown to affect reproductive performance in cattle (Montiel & Ahuja, 2005; Roche et al., 2009). A loss in BCS at calving and during the postpartum period has also been associated with delays in the resumption of ovarian activity (Shrestha et al., 2005), decreased conception rates and prolonged calving-to-conception intervals in dairy cattle (Gillund et al., 2001). Prolonged periods of a negative energy balance during the early weeks of lactation and a delay in achieving a positive energy balance have contributed to periods of anovulation following calving and long intervals of postpartum anoestrus (Beam & Butler 1999; Butler, 2000).
The insulin-like growth factor-I (IGF-I) is a potential mediator of the nutritional effects on reproduction (Zulu et al., 2002; Velazquez et al., 2008). The IGF-I concentrations in the peripheral blood of lactating dairy cows have been related directly to their energy status, with higher concentrations being positively associated with BCS (Beam & Butler, 1999) and nutrient intake (Thissen et al., 1994). The plasma concentrations of IGF-I in postpartum cattle have also been correlated with reproductive performance (Patton et al., 2007; Falkenberg et al., 2008). Thus, increased IGF-I plasma concentrations during early lactation have been associated with the earlier resumption of ovarian function in dairy cattle fed a total mixed ration (TMR) (Thatcher et al., 1996).
The typical seasonal calving system in Victoria, Australia, has been predominantly pasture-based, with the diets being supplemented with cereal grains to balance the diets in order to meet nutritional requirements, and to enhance the milk yield above that which is achieved by pasture feeding alone. The dry matter intake of cows, managed in a pasture-based system, is often less than in the intensive systems in which cows are fed a TMR. Pasture-based diets may, thus, provide insufficient nutrients to sustain a high milk production that can be achieved together with the genetic potential of the modern dairy cow (Kolver & Muller, 1998). Any deficiencies are then likely to be exacerbated during early lactation, when disparities between feed requirements and intake are at the highest. Plasma concentrations of IGF-I may be affected, especially in situations where feed supplementation is inadequate during early lactation. Seasonally-synchronized calving patterns, typical of pasture-based management systems in Victoria as well as New Zealand and Ireland, are adversely affected by prolonged periods of anoestrus, as well as lower conception rates typically associated with a negative energy balance during early lactation. The information is, however, limited by the interactions between the effects of BCS at calving and the different levels of concentrate feeding during early lactation, on circulating concentrations of IGF-I, together with the reproductive performance in dairy cows managed within these pasture-based systems.
The objective of this study was to evaluate the effects of the body condition score (BCS) at calving and the level of grain supplementation during early lactation on: (i) the plasma IGF-I concentrations and (ii) reproductive performance relevant to maintaining a seasonally compact calving pattern. Consequently, the concentrations of IGF-I in cows that had recommenced cyclic ovarian activity and those that were in anoestrus, were investigated, as well as the relationships between IGF-I concentrations and the subsequent reproductive performance.
Materials and Methods
This study was conducted from June to November 2001 (winter to early summer), at the Department of Primary Industries, Kyabram Dairy Centre in the Northern Irrigation District of Victoria, Australia. The project protocol was approved by the Centre's Research Ethics Committee.
Seventy-two multiparous Holstein cows were allocated to three groups and differentially fed from April 2001 to achieve a target BCS, one month before calving, of 4 (3.5 - 4.5), 5 (4.6 - 5.5), or 6 (5.6 - 6.5) units on an 8-point scale (Earle, 1976). The cows were randomly allocated to the groups prior to being differentially fed on the basis of live weight and BCS (March), the expected calving date (August) and according to milk production and composition during the first three months of the previous lactation. The mean (± SEM) live weight and BCS, recorded in early March 2001 (autumn), was 549 ± 64.2 kg and 4.4 ± 0.44, respectively. The cows were fed as three separate groups according to the BCS targets, until the end of lactation, while being supplemented with crushed barley grain and maize silage. When the pastures were no longer adequate, the cows were fed a TMR of maize silage, crushed barley grain, canola meal and barley straw. Cows were fed different quantities of the same TMR after being dried off, to either maintain or gain body condition. Each cow was individually fed a TMR of approximately 1.6% of her live weight to provide in maintenance requirements, without altering the BCS during the last three to four weeks prior to the expected calving date. They were fed their total diet as a single daily meal.
After calving, the cows grazed annual pastures at a pasture allowance level of 35 kg DM/cow/day for five weeks. Metabolizable energy (ME), crude protein (CP) and neutral detergent fibre (NDF) concentrations of the pasture on offer averaged 11.2 MJ, 182 g and 393 g/kg DM, respectively. The cows in each BCS group were offered either 1 or 6 kg DM of the pelleted concentrates per day (Stockdale, 2004). Treatments were set out in a 2 x 3 factorial design, with two levels of grain supplementation (pelleted wheat; 1 or 6 kg fed individually in stalls, twice daily at 06:30 and 15:30, after milking) and the cows divided into one of three BCS groups (4, 5 or 6). Each group comprised 12 cows, fed their allotted diets until the onset of the seasonal, synchronized breeding programme which commenced, on average, 70 days postpartum.
Artificial insemination (AI) commenced on a predetermined date, recorded as the mating starting date (MSD; 70 days postpartum) in this specific seasonally-synchronized calving system. The AI programme then continued for six weeks; after which the bulls were placed in each herd for a further eight-week period (Padula & Macmillan, 2002; Stockdale, 2004). Cows in each herd were monitored for oestrus before and following the MSD, from calving to MSD, using tail paint combined with visual observations on the pastures at least four times per day. Any cow not observed in oestrus by 10 days before the MSD, was subjected to veterinary examination. Cows classified as being in anoestrus, were treated with an intravaginal progesterone-releasing device (CIDR®, Genetics Australia, Bacchus Marsh, Victoria) for eight days, with oestradiol benzoate being administered at the time of device insertion (2 mg i.m.) and 24 h following removal (1 mg i.m.) (Eagles et al., 2002). The cycling or anoestrous status of the cows was based on oestrous detection and confirmed by using milk progesterone profiles. The cows were classified as having ovulated when at least one weekly milk sample with a progesterone concentration of >3.5 ng/mL, had been recorded. None of the weekly milk samples from cows classified as being in anoestrus, had milk progesterone concentrations greater than 3.5 ng/mL by the MSD. In the absence of oestrus, as confirmed by visual observation, the day of ovulation was estimated as having occurred three days prior to the milk sample collection date. Pregnancy was further confirmed by using transrectal ultrasonography, on days 30 and 37 after insemination.
The daily intake of the concentrate was recorded for all the cows and the daily pasture intake during the trial, estimated by using the sward sampling technique (Stockdale & King, 1983). Individual milk yields were recorded weekly and the cows were weighed weekly during the dry period and every day during the study period, immediately after morning milking. The BCS of each cow was assessed weekly, using the 8-point score (Earle, 1976).
Blood was sampled from a coccygeal vein into a 10 mL evacuated, heparinised tube (BD Vacutainer Systems, Plymouth, UK), within 12 h of calving, and thereafter at 1, 5 and 10 weeks of lactation. The blood samples were immediately placed on ice after collection, and centrifuged at 1800 g for 15 min at 4 ºC within 10 min of collection. Plasma was stored at -20 ºC, until assayed for IGF-I. Milk samples, collected weekly, were also centrifuged and the "fat-free" milk stored at -20 ºC until assayed for progesterone.
Total plasma IGF-I concentrations were determined by using a DSL-10-2800 IGF-I ELISA kit (Diagnostic Systems Laboratories Inc, Webster, TX, USA) which had been validated for the IGF-I radioimmunoassay (RIA) of bovine plasma samples in which insulin-like growth factor binding-proteins (IGFBP's) had been dissociated and extracted from IGF-I, using a HPLC (Obese et al., 2008). The intra-assay and inter-assay CV's of the IGF-I ELISA were <6% and <8%, respectively. The minimum detection limit of this assay was 10 ng/mL. Milk progesterone concentrations were determined with the aid of an RIA kit (Spectria Veterinary Progesterone RIA kit, Orion Diagnostica, Espoo, Finland). The inter- and intra-assay CV's for milk pools of 1.19, 2.81 and 5.33 ng/mL were 13.3% and 8.5%, 15.3% and 9.7%, and 16.5% and 11.8%, respectively. The sensitivity of the milk progesterone assays was 0.49 ng/mL.
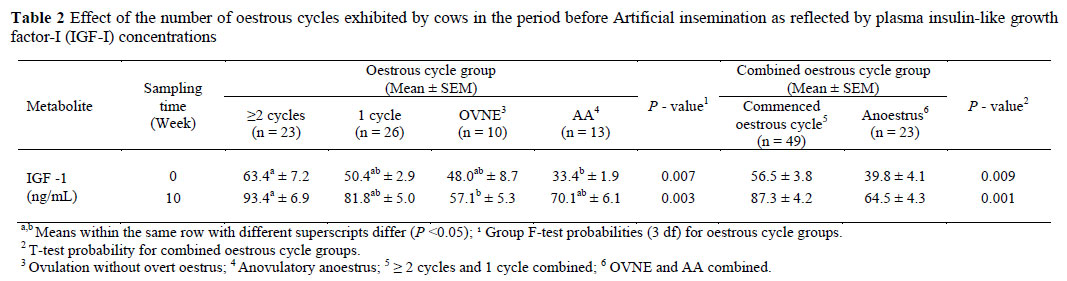
Data for the plasma IGF-I concentration determinations were classified as cows that had resumed cycling, and those that were in anoestrus by the MSD, irrespective of their BCS at calving. In the period before the AI programme commenced, 23 of the 72 cows had exhibited >2 oestrous cycles, 26 had exhibited only 1 cycle and 10 cows had ovulated, but have not displayed overt signs of oestrus (OVNE), while 13 cows were anovulatory (AA). The effects of BCS and level of grain concentrate feeding on the plasma concentration of IGF-I and reproductive parameters were analysed separately on week 0 (calving) and week 10 post-calving, using the ANOVA in SPSS v.11.5 (SPSS, 2002). At week 0 the model for analysis had a BCS at three levels (4, 5 and 6) as the main variable (One-way ANOVA). Cows in the various BCS groups received the grain supplements after calving. The interaction between the BCS and the supplement was not significant for any of the outcome variables and therefore excluded from the model in the analysis for week 10. The model for analysis on week 10 included the main factors, namely the BCS at three levels (4, 5, 6) and supplementation at two levels (1 and 6 kg) (two-way ANOVA). The effects of the number of oestrous cycles exhibited by the cows before the MSD on plasma IGF-I concentrations, were analysed on weeks 0 and 10 by an ANOVA using SPSS v.11.5 (SPSS, 2002). The model had oestrous cycle effects at 4 levels (> 2 cycles, 1 cycle, OVNE, and AA). The four oestrous cycle groups were divided into two main groups namely, cows that had exhibited oestrous cycles and were detected in oestrus (>2 and 1 oestrous cycle) and anoestrous cows (OVNE and AA) and the differences in plasma concentrations of IGF-I between these two groups of cows, evaluated by using an independent sample t-test. The conception rate following the first insemination, pregnancy rates at three weeks and six weeks and the non-pregnancy rates in the 3 BCS and 4 oestrous cycle groups were analysed using Fisher's Exact Test.
Results
No effect of the calving BCS on the plasma IGF-I concentration at either week 0 (calving) or week 10 postpartum, was recorded. The IGF-I concentrations (± SEM) recorded at calving were 50.6 ± 5.3, 52.5 ± 5.3 and 50.5 ± 5.3 ng/mL, while those for week 10 were 80.4 ± 5.8, 80.6 ± 5.8 and 79.0 ± 5.8 ng/mL for the BCS groups 4, 5 and 6, respectively. Higher levels of grain supplementation (6 kg/cow/day) increased (P <0.05) plasma IGF-I concentration, compared to the lower level (1 kg/cow/day) at week 10 (87.8 ± 4.7 vs. 72.1 ng/mL).
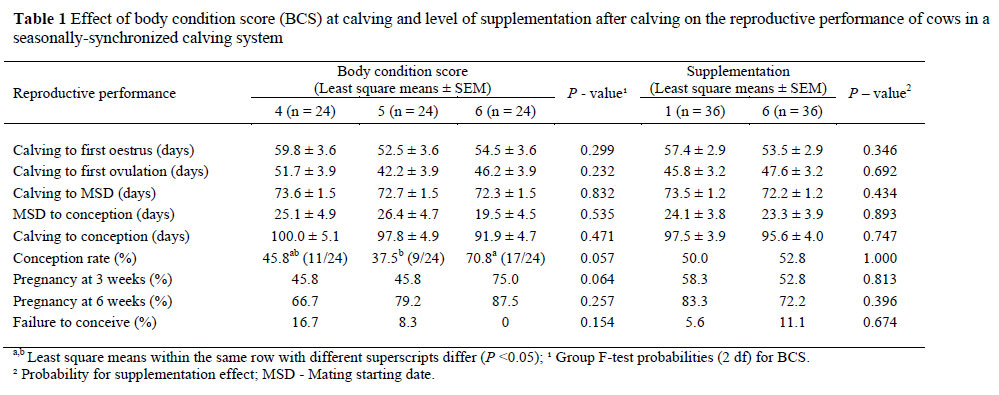
The conception rate at the first AI was the only reproductive parameter affected (P <0.05) by the BCS at calving (Table 1). The conception rates were higher in cows with a BCS of 6, compared to those with a BCS of 5. Similar conception rates were recorded for cows in the BCS 4 group than for those in the BCS 5 or 6 groups.
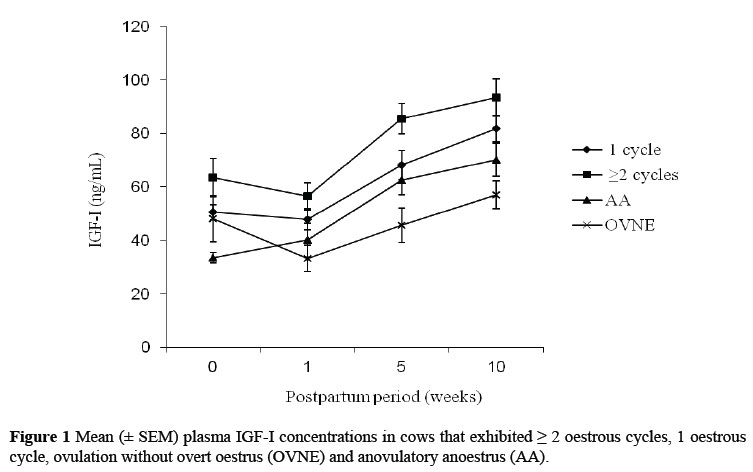
Plasma IGF-I concentrations were low in every group at calving and week 1 (ranging from 33.4 ± 1.9 to 63.4 ± 7.2 ng/mL at calving and 33.3 ± 4.9 to 56.5 ± 5.1 ng/mL at week 1) ( Figure 1). There was a steady increase from week 1 to week 10, when the group mean ranged from 57.1 ± 5.3 to 93.4 ± 6.9 ng/mL. A higher mean plasma IGF-I concentration was recorded for cows with multiple oestrous cycles, before the MSD compared to the AA (anovulatory) cows at week 0 (calving) (P <0.05) and OVNE (ovulation without overt signs of oestrus) cows at week 10 (P <0.05) (Table 2). The results from the independent sample t-test showed that the plasma IGF-I concentrations were higher in cows that had commenced their oestrous cycles and been detected in oestrus, compared to the anoestrous cows at week 0 (P <0.01) and week 10 (P <0.01) (Table 2).
The cows with 1 or >2 cycles demonstrated shorter (P <0.05) intervals from calving to first oestrus, compared to the OVNE or AA cows (Table 3). The average calving time to the first ovulation interval, for all cows, was 46.7 ± 2.2 days, while the interval from calving to first ovulation was shorter (P <0.05) in cows with >2 cycles, 1 cycle or OVNE cows, compared to the AA cows (Table 3). The interval from calving to the MSD recorded was also shorter (P <0.05) for the OVNE cows, compared to the cows with >2 cycles (Table 3). Days to conception, conception rate at first insemination, pregnancy rates and non-pregnant rates did not differ between the groups.
Discussion
This study showed that calving BCS did not influence the concentration of IGF-I in the blood. The level of supplementation following calving, however, affected plasma IGF-I concentrations. In addition, significant relationships were recorded between plasma IGF-I concentration and the cyclic status, at 10 weeks post-calving, and for the interval from calving to first ovulation. These findings indicate that diet can affect plasma IGF-I levels, which in turn may be involved in affecting the length of the postpartum anoestrous period. The plasma IGF-I concentrations recorded were also less at calving than at week 10 postpartum. These concentrations are generally low at calving and in early lactation, when many dairy cows experience a negative energy balance, but then increase as lactation progresses and the cows move towards a more positive energy balance (Lucy, 2000). The higher level (6 kg) of grain supplementation was associated with an increased plasma concentration of IGF-I at week 10. This supports the hypothesis that nutrient intake (protein and energy) partly influences blood IGF-I concentrations (Thissen et al., 1994).
Neither the calving BCS nor the level of supplementation influenced any of the reproductive variables measured, apart from the conception rate at first AI. The cows with the higher calving BCS achieved the highest pregnancy rates six weeks after the MSD (87.5% and 79.2% vs. 66.7%) (Table 3). A positive relationship between the BCS and 6-week pregnancy rate was also recorded in 123 seasonally-calving herds in Southern Australia, where the average pregnancy rate at six weeks after AI, was 62% (Morton, 2004). Other studies involving pasture-based systems have also demonstrated a positive effect on the calving BCS and on the postpartum anoestrous period and reproductive performance (Buckley et al., 2003; Miekle et al., 2004; Roche et al., 2007).
The cows that had commenced oestrous cycles with detected oestrus (>2 cycles and 1 cycle combined) before the MSD, had higher plasma IGF-I concentrations, compared to the anoestrous cows (OVNE and AA combined). These findings are in agreement with other reports for Holstein-Friesian cows in pasture-based systems (Miekle et al., 2004; McDougall et al., 2005). Generally, the plasma IGF-I concentrations for cows with single or multiple oestrous cycles, in this study, were higher at calving and also throughout the entire study period compared to the cows that were anovulatory (AA), or those that ovulated, but failed to display overt signs of oestrus sufficient to be noticed by the herdsmen (OVNE) (Figure. 1). This finding suggests that ovarian follicular development, sufficient to result in oestrus and ovulation, may be associated with an early increase in circulating IGF-I concentration postpartum. Circulating IGF-I thus seems to stimulate ovarian function by acting synergistically with gonadotropins to promote follicular growth, differentiation and steroidogenesis of the ovarian cells (Lucy, 2000). Low levels of circulating IGF-I concentrations may thus contribute to reduced follicular responsiveness to a given level of LH support, low oestradiol synthesis, and anovulation of the dominant ovarian follicles in the postpartum cows experiencing a negative energy balance, especially during early lactation (Beam & Butler 1999; Diskin et al., 2003; Roche, 2006).
Conclusions
Plasma IGF-I concentration was not affected by BCS at calving. The level of cereal grain concentrate supplementation, however, influenced the plasma levels of IGF-I. The differences in plasma IGF-I concentrations were associated with postpartum ovarian activity. The cows with lower plasma IGF-I concentrations, during the early postpartum period, were at greater risk of experiencing extended intervals from calving to first ovulation, and of not showing overt signs of oestrus at that ovulation. A greater understanding of the role of IGF-I in postpartum ovarian activity, independent of variations in concentration associated with the feeding of an energy-rich supplement is needed. This could aid in achieving a better pre-calving and post-calving nutritional management programme, for improved herd reproductive performance by minimizing the incidence of postpartum anoestrus in pasture-based herds with seasonally-synchronized calving patterns.
Acknowledgements
The authors are grateful to Meg Jenkins of the Kyabram Dairy Centre for the collection of milk and plasma samples. Appreciation is expressed to the Department of Primary Industries, University of Melbourne, Dairy Australia and Primegro Ltd. (Adelaide) for financially supporting the project.
References
Beam, S.W. & Butler, W.R., 1999. Effects of energy balance on follicular development and first ovulation in postpartum dairy cows. J. Reprod. Fertil. (Suppl.) 54, 411- 424. [ Links ]
Buckley, F., O'Sullivan, K., Mee, J.F., Evans, R.D. & Dillon, P., 2003. Relationships among milk yield, body condition, cow weight, and reproduction in spring-calving Holstein-Friesians. J. Dairy Sci. 86, 2308-2319. [ Links ]
Butler, W.R., 2000. Nutritional interactions with reproductive performance in dairy cattle. Anim. Reprod. Sci. 60-61, 449-457. [ Links ]
Diskin, M.G., Mackey, D.R., Roche, J.F. & Sreenan, J.M., 2003. Effects of nutrition and metabolic status on circulating hormones and ovarian follicle development in cattle. Anim. Reprod. Sci. 78, 345-370. [ Links ]
Eagles, V.E., Nation, D.P. & Macmillan, K.L., 2002. New protocols for effectively treating anoestrous cows in seasonally calving dairy herds. Proc. Dairy Res. Conf, the University of Melbourne, Australia. pp. 75-79. [ Links ]
Earle, D.F., 1976. A guide to scoring dairy cow condition. J. Agric. (Victoria) 74, 228-231. [ Links ]
Falkenberg, U., Haertel, J., Rotter, K., Iwersen, M., Arndt, G. & Heuwieser, W., 2008. Relationships between the concentration of IGF-1 in serum in dairy cows in early lactation and reproductive performance and milk yield. J. Dairy Sci. 91, 3862-3868. [ Links ]
Gillund, P., Reksen, O., Grohn, Y.T. & Karlberg, K., 2001. Body condition related to ketosis and reproductive performance in Norwegian dairy cattle. J. Dairy Sci. 84, 1390-1396. [ Links ]
Kolver, E.S. & Muller, L.D., 1998. Performance and nutrient intake of high producing Holstein cows consuming pasture or a total mixed ration. J. Dairy Sci. 81, 1403-1411. [ Links ]
Lucy, M.C., 2000. Regulation of ovarian follicular growth by somatotropin and insulin- like growth factors in cattle. J. Dairy Sci. 83, 1635-1647. [ Links ]
McDougall, S., Blache, D. & Rhodes, F.M., 2005. Factors affecting conception and expression of oestrus in anoestrous cows treated with progesterone and oestradiol benzoate. Anim. Reprod. Sci. 88, 203-214. [ Links ]
Miekle, A., Kulcsar, M., Chilliard, Y., Febel, H., Delavaud, C., Cavestany, D. & Chilibroste, P., 2004. Effects of parity and body condition at parturition on endocrine and reproductive parameters of the cow. Reprod. 127, 727-737. [ Links ]
Montiel, F. & Ahuja, C., 2005. Body condition and suckling as factors influencing the duration of postpartum anestrus in cattle: a review. Anim. Reprod. Sci. 85, 1-26. [ Links ]
Morton, J., 2004. Determinants of reproductive performance of dairy cows in commercial herds in Australia. PhD thesis, the University of Melbourne. Australia. [ Links ]
Obese, F.Y., Humphrys, S., Macmillan, K.L. & Egan, A.R., 2008. Measuring concentrations of insulin-like growth factor-I with an enzyme-linked immunosorbent antibody assay in plasma samples from Holstein cows. J. Dairy Sci. 91,160-168. [ Links ]
Padula, A.M. & Macmillan, K.L., 2002. Reproductive responses of early postpartum dairy cattle to continuous treatment with a GnRH agonist (deslorelin) for 28 days to delay the resumption of ovulation. Anim. Reprod. Sci. 70, 23-36. [ Links ]
Patton, J.D., Kenny, A., McNamara, S., Mee, J.F., O'Mara, F.P., Diskin, M.G. & Murphy, J., 2007. Relationships among milk production, energy balance, plasma analytes, and reproduction in Holstein-Friesian cows. J. Dairy Sci. 90, 649-658. [ Links ]
Roche, J.F., 2006. The effect of nutritional management of the dairy cow on reproductive efficiency. Anim. Reprod. Sci. 96, 282-296. [ Links ]
Roche, J.R., Macdonald, K.A., Burke, C.R., Lee, J.M. & Berry, D.P., 2007. Associations among body condition score, body weight, and reproductive performance in seasonal-calving dairy cattle. J. Dairy Sci. 90, 376-391. [ Links ]
Roche, J.R., Friggens., N.C., Kay, J.K., Fisher, M.W., Stafford, K.J. & Berry, D.P., 2009. Invited review: Body condition score and its association with dairy cow productivity, health, and welfare. J. Dairy Sci. 92, 5769-5801. [ Links ]
Shrestha, H.K., Nakao, T., Suzuki, T., Akita, M. & Higaki, T., 2005. Relationships between body condition score, body weight, and some nutritional parameters in plasma and resumption of ovarian cyclicity postpartum during pre-service period in high-producing dairy cows in a subtropical region in Japan. Theriogenology 2005, 855-866. [ Links ]
SPSS Inc., 2002. SPSS for Windows version 11.5. Chicago, Illinois, U.S.A. [ Links ]
Stockdale, C.R., 2004. Effects of feeding of concentrates during early lactation on the yield and composition of milk from grazing dairy cows with varying body condition score at calving. Aust. J. Exp. Agric. 44, 1-9. [ Links ]
Stockdale, C.R. & King, K.R., 1983. A comparison of two techniques used to estimate the herbage intake of lactating dairy cows in a grazing experiment. J. Agric. Sci., Camb. 100, 227-230. [ Links ]
Thatcher, W.W., de la Sota, R.L., Schmitt, E.J., Diaz, T.C., Badinga, L., Simmen, F.A., Staples, C.R. & Drost, M., 1999. Control and management of ovarian follicles in cattle to optimize fertility. Reprod. Fertil. Dev. 8, 203-217. [ Links ]
Thissen, J.P., Ketelslegers, J.M. & Underwood, L.E., 1994. Nutritional regulation of the insulin-like growth factors. Endocr. Rev. 15, 80-101. [ Links ]
Velazquez, M.A., Spicer, L.J. & Wathes, D.C., 2008. The role of endocrine insulin-like growth factor-I (IGF-I) in female bovine reproduction. Domest. Anim. Endocrinol. 35, 325-342. [ Links ]
Zulu, V.C., Nakao, T. & Sawamukai, Y., 2002. Insulin-like growth factor-I as a possible hormonal mediator of nutritional regulation of reproduction in cattle. J. Vet. Med. Sci. 64, 657-665. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. See: http://creativecommons.org/licenses/by/2.5/za
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science.
# Corresponding author: fyobese@yahoo.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}