










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 no.4 Pretoria Jan. 2011
The effect of tannin with and without polyethylene glycol on in vitro gas production and microbial enzyme activity
I.V. Nsahlai#; F.N. Fon; N.A.D. Basha
Animal and Poultry Science, SASA, University of KwaZulu-Natal, Pietermaritzburg, 3209, South Africa
ABSTRACT
The aim was to determine the effect of tannin on the nutritive value and microbial enzyme activity in vitro. The leaves and pods of the Acacia sieberiana and Acacia nilotica trees were subjected to chemical analysis and in vitro fermentation with and without polyethylene glycol (PEG; 35 mg/g). Gas production (GP), true degradability (TD) and the proteolytic and fibrolytic enzyme activities in digesta were determined. Data analyses accounted for the effects of feed type, feed fraction, PEG and their interactions. Leaves and pods had similar NDF and ADF contents in A. sieberiana but differed in both attributes in A. nilotica. The leaves had a higher protein, but lower condensed tannin (CT) contents than the pods. A. sieberiana had more CT than A. nilotica, but both had a similar maximum GP. PEG increased the maximum GP, stimulated GP from the soluble fraction and shortened the lag phase. PEG increased the TD for leaves and pods of A. sieberiana, but suppressed TD in fractions of A. nilotica. PEG also increased the activity of protease enzymes, xylanase, endo- and exocellulases, indicating that tannins depress the digestibility of feeds partly by suppressing the activity of these enzymes. Consequently, condensed tannin depressed the digestibility of feed in the rumen by affecting the activity of enzymes involved in carbohydrate and protein breakdown, the effects of which could be reduced with the use of PEG.
Keywords: Protease, xylanase activity, endo- and exocellulase activity, fibrolase, degradation kinetics, PEG
Introduction
Livestock production in tropical Africa is limited mainly by the availability and quality of feed during the dry season (Ngwa et al., 2000) when animals lose part of the weight they gained in summer. This is largely the result of a very low input in the prevailing livestock production systems which are heavily reliant on protein-deficient natural pastures. The biodiversity and quality of feed items in animal diets vary seasonally in such a way that the proportion of browse selected in the diet increases with the increasing scarcity of palatable grasses and grass fractions. Numerous studies have demonstrated that browses (Degen et al., 1995; Nsahlai et al., 1995; Ngwa et al., 2001) and some crop residues, such as bird resistant sorghum varieties (Nsahlai et al., 1998), are endowed with anti-nutritional factors such as condensed tannins (CT). Trees and shrub foliage are of importance in animal production because they do not compete with human food and can provide protein supplements, especially in the dry season. In African, particularly in goat, production systems the animals spend 75% of their time browsing on shrubs and trees. These feeds are rich in anti-nutritional factors, particularly tannins, because they reduce the intake of food, depress the breakdown rate of protein and fibre, reduce the efficiency of microbial protein synthesis (Makkar, 2003) and are poisonous (McSweeney et al., 2001). Consequently, detannification of these feeds with wood ash and storage at room temperature (Makkar & Singh, 1991; 1993), dilution, drying, extraction using organic solvents (Makkar, 2003), biodegradation by white rot fungi (Gamble et al, 1996) and binding with polyethylene glycol (PEG) and polyvinyl pyrrolidone (PVP) (Makkar et al., 1995c) have been advocated.
In our previous studies it was demonstrated that Menz sheep given oilseed cake following the consumption of tannin-rich browses (Nsahlai et al., 1999) achieved higher live-weight gains than others given diets in the reversed order. This benefit could be interpreted as being due to the interaction of feed protein and tannin and/or a consequence of depressed proteolytic activity in the rumen; both mechanisms would increase the proportion of dietary protein that bypasses the rumen undegraded. This study explores the effect of tannin-rich feeds on the kinetics of fermentation of fibre and soluble components, and on the activity of fibrolytic and proteolytic enzymes.
Materials and Methods
The leaves and pods of Acacia sieberiana and Acacia nilotica trees were harvested at pod maturity from five plants at different spots on the Ukulinga Research Farm (30º 24'S, 29º 24' E), University of KwaZulu-Natal, South Africa, in summer (February). The research farm is situated in a subtropical hinterland which is approximately 700 m above sea level. The climate is characterized by an annual rainfall of 735 mm, which falls mostly in summer between October and April. Mean annual maximum and minimum temperatures are 25.7 ºC and 8.9 ºC, respectively. Light to moderate frost occurs in winter.
Samples were transported to the laboratory and dried in a force-air oven at 40 ºC for 48 h. Dried samples were separated into two portions, each of which was ground through either a 1-mm or a 3-mm screen of the laboratory harmer mill (lKA cutting mill). Subsequently, the 1-mm fine samples were used for chemical analysis and in vitro gas production, whilst the 3-mm fine ones were used to determine the ruminal degradation parameters
Degradation was based on the nylon bag technique described by Mehrez & Ørskov (1977) on 5 g air dry sample per bag (size 9 x 16 cm and pore size 40 - 50 µm; Polymon, Switzerland). Two fistulated cows (a Jersey and a Friesland), given pasture hay (containing 32 g crude protein (CP), 690 g neutral detergent fibre (NDF) and 453 g acid detergent fibre (ADF)/kg dry matter (DM)) supplemented with lucerne (1.5 kg/head/day), served to determine the nylon bag degradation parameters and as source of rumen inoculum for the in vitro gas production assay. Following incubation, samples were withdrawn from the rumen after 3, 6, 24, 48, 72 and 96 h and washed. The 0 h bags were not incubated but washed together with other bags in 6 cycles of 5 min each in a semi-automatic washing machine (Hoovermatic Superfast), spun, dried in an oven at 60 ºC for 48 h and weighed.
A total of 1.0 ± 0.0010 g DM of each sample was weighed into a 250 mL Duran bottle for in vitro incubation. During a run, each feed sample was represented by five Duran bottles. A solution of PEG (MW 4000) was made by dissolving 3.5 g of PEG in 100 mL of distilled water. Two bottles of each sample were treated by adding 1 mL of PEG solution and mixing thoroughly.
The in vitro gas production was done in two runs following the automated technique described by Pell & Schofield (1993). A salivary buffer solution was prepared according to McDougall (1948). A 67 mL solution of the buffer was added to each sample, control or blank bottle. The bottles were kept in the incubator (39 ºC) for 1 h to allow the soaking of the substrate prior to adding the rumen fluid. Meanwhile a mixture of the rumen fluid was collected from two cows prior to morning feeding. The rumen fluid was collected and filtered through four layers of cheesecloth into a pre-warmed flask (39 ºC) that had been flushed with CO2, transported to the laboratory, and inoculation was completed by adding 33 mL of the rumen fluid to each bottle under a stream of CO2 and tightening the bottle lids. The Duran bottles were incubated in the incubator channels and pressure sensors were fitted. Pressure was logged at 20 min intervals during a 72-h incubation period. The model described by Campos et al. (2004) was fitted to gas volumes to determine the gas production kinetics following a two-step process:
GP = GPMAX.(1-exp-kGP.t); GP = GPsol /[1 +exp(2 + 4kso1 (TL - t))] + (GPMAX - GPsol /[1 +exp(2 + 4kfibre (TL -t))] where GP is the total gas volume (mL) at time t, GPMAX is the asymptotic GP, kGP is the overall rate of gas production, GPsol is the gas volume (mL) from fasting (soluble sugars and starch) and GPfibre = (GPMAX - GPsol) is the GP from slowly (cellulose and hemicellulose) degradable fractions, ksol and kfibre are the degradation rates (/h) for fasting and slowly degradable fractions, respectively, and TL is the bacteria colonization or Lag time (h).
Upon completion of each run of incubation, one Duran bottle pertaining to each of the treatments was selected at random for the determination of enzyme activity. Proteolytic activity on azo-casein (sulfanilamide-azo-casein, Sigma A2765) was measured following the method described by Kopecny & Wallace (1982). Crude protein broth was extracted, treated and concentrated following the procedure described by Fon (2006). The Bradford dye binding assay (Bradford, 1976) was used for protein determination. Reducing sugars in sampled solutions were determined using the dinitrosalicylic (DNS) method (Miller, 1960). Crystalline cellulase was assayed following the method described by Gerrit et al. (1984). The activity of carboxymethylcellulase (CMCase) was determined by following the method described by Gerrit et al. (1984). Xylanase activity was essayed according to the procedure described by Seyis & Aksoz (2005). The quantity of CP extract used, was 150 µg for endo-cellulase and xylanase and 350 µg for crystalline cellulose, as defined by Fon (2006).
Feed samples and/or nylon bag residues were analyzed for DM, CP and nitrogen (N) in extracted acid detergent fibre (ADIN), following the standard AOAC methods (AOAC, 1990). Acid detergent fibre (ADF) and NDF, using heat stable alpha-amylase, were extracted by following the methods of Van Soest et al. (1991). Condensed tannin concentration was determined following the procedure described by Makkar & Goodchild (1996).The degradation parameters of DM and N were estimated by fitting the non-linear model proposed by McDonald (1981) and modified by Dhanoa (1988), to the degradation data:
Y = W + B[1 -e -c(T-LT)]
where 'Y' is the disappearance of DM or N at time 'T', 'W' is the washing loss or solubility, 'B' the degradable part of the insoluble fraction, 'c' the fractional rate of degradation of the B fraction; and LT is the lag time. The potential degradability was calculated as A+B.
In this study two species and two fractions were considered. Consequently, a 2 x 2 factorial arrangement was applied when considering degradability. In addition, PEG (at two levels) was added to make a 2 x 2 x 2 factorial arrangement when analyzing all the other variables. All variables were analyzed using the general linear model (GLM) available in the SAS system (SAS, 2000). The statistical model accounted for the effect of species, feed fraction and their interaction (for degradability variables) and species, feed fraction, PEG and their interaction (for GP data) while correcting for the effect of run.
Results and Discussion
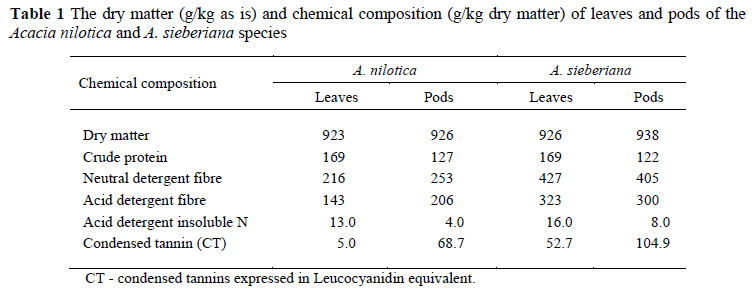
The leaves were much richer in CP than the pods, whilst the two species had a similar CP content (Table 1). Both fractions of A. nilotica had a lower content than the corresponding fractions of A. sieberiana. ADIN was higher in the leaves than in the pods. However, the pods had a higher CT concentration than the leaves. Both fractions of A. sieberiana had a higher CT content than A. nilotica. With the exception of the leaves of A. nilotica, all other feeds had more than 5% CT, which is higher than the 3% recommended beneficial level for increased "bypass" protein (Barry & McNabb, 1999), but could be suitable for reducing gastro-intestinal nematode load.
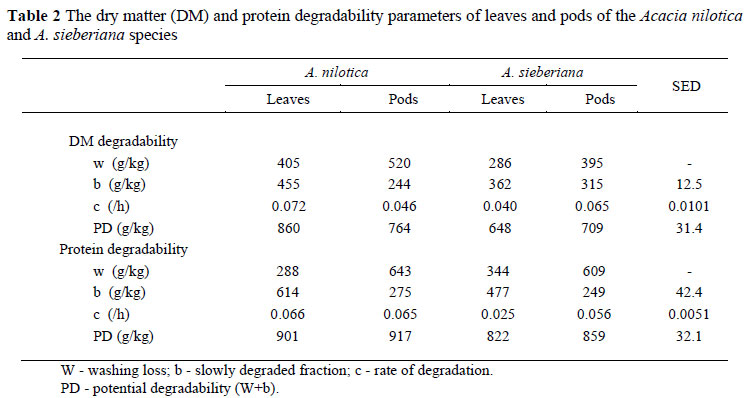
The DM and N were potentially more degradable (P <0.05 and P = 0.055) for A. nilotica than for A. sieberiana (Table 2). The dependence of N loss on DM loss has been demonstrated in numerous studies (Nsahlai et al., 1995; Kaitho et al., 1997). The observation that A. sieberiana had a lower (P <0.001) soluble fraction than A. nilotica would explain the superior potential degradability ascribed to the latter. The solubility of N and DM suggests that most N in pods are in the soluble fraction whilst most N in leaves are in the insoluble fraction. This accords with the ADIN contents. A. nilotica had a faster (P <0.05) protein degradation rate than A. sieberiana. The direction of these effects suggests that fibre and/or tannin might have reduced the rate and the potential degradability of DM and N in the rumen. However, the fact that the pods had faster protein degradation rates than the leaves is inconsistent with this interpretation and may be related to the distribution of protein and CT between seed and husk. It is possible that the full effect of CT has not been realized because of the diluting effect of the rumen environment in view of free movement of fluid into and out of a nylon bag (Nsahlai & Ummuna, 1996). The interaction effect of species by fraction on DM and N degradation rates indicated that: (1) Leaf DM degraded faster (P <0.05) than the pods for A. nilotica but slower (P <0.05) than the pods for A. sieberiana, and (2) leaf protein degraded at similar rates, like the pod protein for A. nilotica, but slower than the pod protein for A. sieberiana. These degradation parameters suggest that, with the exception of A. sieberiana the leaves have an intrinsic moderate potential to supply "bypass" protein, while the rest of the feeds are potential sources of rumen degradable protein.
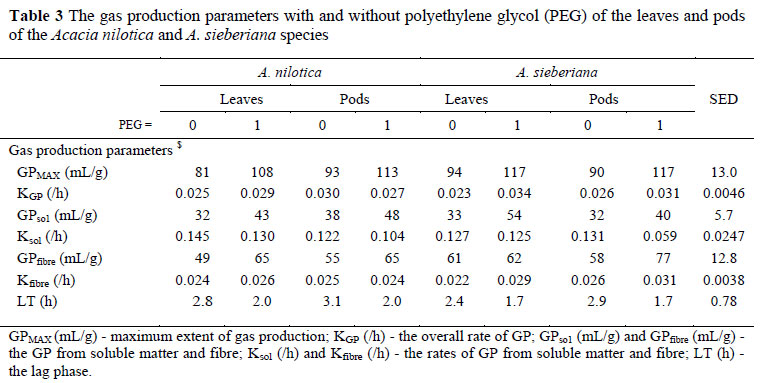
The average pH was 6.6 ± 0.103 and was similar for all treatments. Contrary to expectation the GPMAX was neither affected by species, nor by fraction (Table 3). However, treatment with PEG increased (P <0.01) GPMAX and GPso1, tended to increased (P = 0.11) GPfibre and sped up (P = 0.13) kGP, but shortened (P <0.05) the lag time. Anti-fermentation factors, e.g. CT, when released, are retained in the batch culture where they could exert a maximum effect of suppressing fermentation unlike with the nylon bag technique where CT can be washed out of the nylon bag before they have had the opportunity to form complexes with macro-molecules. It is thus surprising and moreover inconsistent, with the nylon bag technique that the effects on species and feed fraction were not manifested in the GP technique. A significant (P <0.05) effect on species by fraction interaction indicated that the leaves had a higher GPso1 than the pods for A. sieberiana, but that the trend was reversed for A. nilotica. The former observation is consistent with the tannin content in leaves and pods of A. sieberiana, but the reversed trend in A. nilotica is difficult to explain. It is possible that the richness of the leaves of A. nilotica, in catechin gallates (hydrolysable tannins), may be responsible.
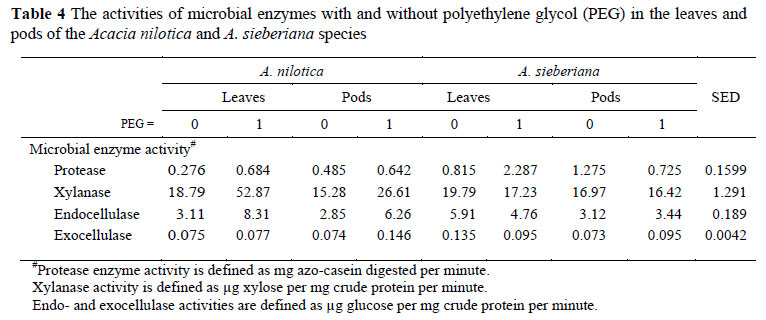
One possible strategy for increasing ruminal "bypass" protein would be to decrease the activity of protease enzymes. Unexpected from the CT contents, A. sieberiana generally elicited higher (P <0.05) protease activity than A. nilotica (Table 4). The interaction effect (Table 4) indicated that treatment with PEG increased protease activity for both fractions of A. nilotica and for the leaves of A. sieberiana, but depressed the protease activity for the pods of A. sieberiana. These results support the expectation that the use of PEG would negate the effect of tannins and increase protein hydrolysis for microbial use. The fact that the largest effect of PEG was in the leaves of A. sieberiana is suggestive of a higher reactivity of tannins in this feed. The poor response of protease activity and all the other enzymes to PEG in the pods of A. sieberiana suggests that the added PEG might not have been enough to saturate and displace the tannins from complexes with these hydrolytic enzymes. Ngwa et al. (2003) also reported a negative effect of tannins on protease activity in the pods of A. sieberiana and A. nilotica.
The effects of feed type, feed fraction, PEG and their interactions affected (P <0.00l) the activity of all fibrolases, except for crystalline cellulose (exocellulase) which was not affected by feed fraction. The PEG treatment also increased the activities of endocellulases and xylanase for both fractions of A. nilotica, but the activities of both enzymes either decreased (for leaves) or remained unchanged for the pods of A. sieberiana following PEG treatment. PEG increased the activity of crystalline cellulases in the pods of both species, had no effect on the leaves of A. nilotica but decreased the activity in the leaves of A. sieberiana. Different mechanisms proposed to explain tannin antimicrobial activity, include the inhibition of extracellular microbial enzymes, deprivation of the substrates required for microbial growth or direct action on microbial metabolism through the inhibition of oxidative phosphorylation and iron deprivation (Scalbert, 1991; Chung et al., 1998; Ngwa et al., 2003). It is perhaps a consequence of these mechanisms that PEG shortened the lag phase and increased the extent of GP from solubles.
It could be asked why the effect of PEG was not as pronounced in the kinetics of fibre digestion as in the digestion of solubles. Since fibrolytic bacteria usually became attached to the substrate, it is possible that some microbial attachment processes could have been disturbed due to the interaction of its protein component with tannins, which could have delayed attachment during colonization. However, following successful attachment, the catalytic domain is brought into close proximity with fibre which is then hydrolyzed. It is possible that this attachment mechanism reduces the chance of interaction between free condensed tannins in solution and extra-cellular fibrolytic enzymes at the site of digestion. This could explain why the positive effect of PEG on microbial enzyme activity in A. nilotica did not translate into a significantly increased gas production kinetic of fibre. We are unable to explain why PEG failed to consistently enhance fibrolytic enzyme activity in the leaves of A. sieberiana leaves.
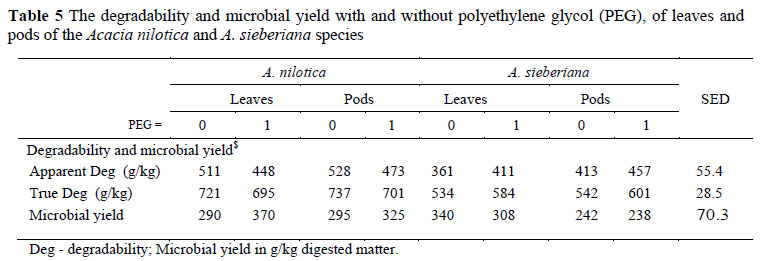
The superior true degradability of A. nilotica is consistent with the nylon bag results, which is not reflected in the GPMAX (Table 5). The fact that PEG increased the true degradability of A. sieberiana is consistent with the general effect of tannins to depress digestibility (Degen et al., 1995; Ngwa et al., 2003). The fact that PEG depressed the true degradability of A. nilotica is difficult to explain. Ironically, the increased fermentative activity following PEG treatment, as reflected in GPMAX, did not result in an increased microbial yield.
Conclusion
This study demonstrated that the negative effect of tannins on digestion is not just the result of their interaction with macro-molecules in the feeds, but also a reflection of their interference with fibrolytic and proteolytic enzymes. Consequently, neutralization of the tannin effect on PEG resulted in enhanced rates and extent of digestion, and shorter lag times preceding the onset of digestion.
References
Seyis, I. & Aksoz, N., 2005. Investigation of factors affecting xylanase activity from Trichoderma harzianum 1073 D3. 48, 187-193. [ Links ]
AOAC, 1990. Official Methods of Analysis, 15th ed. Association of Official Analytical Chemists. AOAC, Washington, USA. pp. 69-88. [ Links ]
Barry, T.N. & McNabb, W.C., 1999. The implications of condensed tannins on the nutritive value of temperate forages fed to ruminants. Br. J. Nutr. 81, 263-272. [ Links ]
Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254. [ Links ]
Campos, F.P., Sampaio, A.A.M., Bose, M.L.V., Vieira, P.F. & Sarmento, P., 2004. Evaluation of in vitro gas production of roughages and their mixtures using curves subtraction method. Anim. Feed Sci. Technol. 116, 161-172. [ Links ]
Chung, K.T., Lu, Z. & Chou, M.W., 1998. Mechanism of inhibition of tannic acid and related compounds on the growth of intestinal bacteria. Food Chem. Toxicol. 36, 1053-1060. [ Links ]
Degen, A.A., Becker, K., Makkar, H.P.S. & Borowy, N., 1995. Acacia saligna as a fodder tree for desert livestock and the interaction of its tannins with fibre fractions. J. Sci. Food Agric. 68, 65-71. [ Links ]
Dhanoa, M.S., 1988. On the analysis of dacron bag data for low degradability feeds. Grass For. Sci. 43, 441-444. [ Links ]
Fon, F.N., 2006. Fibrolytic enzyme activity of herbivore microbial ecosystems. MSc thesis, University of KwaZulu-Natal, South Africa. [ Links ]
Gamble, G.R., Akin, D.E., Makkar, H.P.S. & Becker, K., 1996. Biological degradation of tannins in Sericea lespedeza by the white rot fungi Ceriporiopsis subvermispora and Cyanthus stercoreus analyzed by solid state 13C NMR spectroscopy. Appl. Environ. Microbiol. 62, 3600-3604. [ Links ]
Gerrit, B., Marjo, F., Frank, M. & Fons, G., 1984. The cellulase of Trichoderma viride. Purification characterization and comparison of all detectable endoglucanases, exoglucanases and β-glucosidases. Eur. J. Biochem. 146, 301-308. [ Links ]
Henry, B., Nick, V., Desh, P. & Gordon, A., 1974. Purification and characterisation of two cellulases from Auxin-treated Pea Epicoltyls. J. Biol. Chem. 10, 1012-1018 [ Links ]
Kaitho, R.J., Nsahlai, I.V., Williams, B.A., Umunna, N.N., Tamminga, S. & Van Bruchem, J., 1997. Inter-relationships between palatability, rumen degradability, gas production and chemical composition in browses. Agroforestry Systems 39, 129-144. [ Links ]
Kopecny, J. & Wallace, J., 1982. Cellular location of some properties of proteolytic enzymes of rumen bacteria. Appl. Environ. Microbiol. 43, 1026-1033. [ Links ]
Makkar, H.P.S., 1995. Quantification of tannins: A laboratory manual. International Centre for Agricultural Research in the Dry Areas (ICARDA), Aleppo, Syria. p. 25. [ Links ]
Makkar, H.P.S., 2003. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rumin. Res. 49, 241-256. [ Links ]
Makkar, H.P.S. & Singh, B., 1991. Distribution of condensed tannins (proanthocyanidins) in various fraction of young and mature leaves of some oak species. Anim. Feed Sci. Technol. 32, 253-260. [ Links ]
Makkar, H.P.S. & Singh, B., 1993. Effects of storage and urea addition on detannification and in sacco dry matter digestibility of mature oak (Quercus incana) leaves. Anim. Feed Sci. Technol. 41, 247-259. [ Links ]
Makkar, H.P.S., Blummel, M. & Becker, K., 1995. Formation of complexes between polyvinyl pyrrolidone and polyethylene glycol with tannins and their implications in production and true digestibility in in vitro techniques. Br. J. Nutr. 73, 897-913. [ Links ]
McDonald, I.M., 1981. A revised model for the estimation of protein degradability in the rumen. J. Agric. Sci., Camb. 96, 251-252. [ Links ]
McSweeney, C.S., Palmer, B., McNeil, D.M. & Krause, D.O., 2001. Microbial interactions with tannins: nutritional consequences for ruminants. Anim. Feed Sci. Technol. 91, 83-93. [ Links ]
Mehrez, N.P. & Ørskov, E.R., 1977. A study of the artificial fibre bag technique for determining the digestibility of feeds in the rumen. J. Agric. Sci., Camb. 88, 645-650. [ Links ]
Miller, G., Blum, R., Glennon, W. & Burton, A., 1960. Measurement of CMC-ase activity. Analyt. Biochem. 2, 127-132. [ Links ]
Miller, G., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31, 426-428. [ Links ]
Ngwa, A.T., Pone, D.K. & Mafeni, J.M., 2000. Feed selection and dietary preferences of forage by small ruminants grazing natural pastures in the Sahelian zone of Cameroon. Anim. Feed Sci. Technol. 88, 253-266. [ Links ]
Ngwa, A.T., Nsahlai, I.V. & Bonsi, M.L.K., 2002. The rumen digestion of dry matter, nitrogen and cell wall constituents of the pods of Leucaena leucocephala and some Acacia species. J. Sci. Food Agric. 82, 98-106. [ Links ]
Ngwa, A.T., Nsahlai, I.V. & Iji, P.A., 2003. Effect of feeding pods or alfalfa in combination with poor quality grass straw on microbial enzyme activity and production of VFA in the rumen of South African Merino sheep. Small Rumin. Res. 48, 83-94. [ Links ]
Nsahlai, I.V. & Umunna, N.N., 1996. Comparison between reconstituted sheep faeces and rumen fluid inocula and between in vitro and in sacco digestibility methods as predictors of intake and in vivo digestibility. J. Agric. Sci., Camb. 126, 235-248. [ Links ]
Nsahlai, I.V., Siaw, D.E.K.A. & Umunna, N.N., 1995. Inter-relationships between chemical constituents, dry matter and nitrogen degradability of fresh leaves of multi-purpose trees. J. Sci. Food Agric. 69, 235-246. [ Links ]
Nsahlai, I.V., Umunna, N.N. & Osuji, P.O., 1998. Complementarity of bird-resistant and non-bird-resistant varieties of sorghum stover with cottonseed cake and noug (Guizotia abyssinica) cake when fed to sheep. J. Agric. Sci., Camb. 130, 229-239. [ Links ]
Nsahlai, I.V., Umunna, N.N. & Osuji, P.O., 1999. Influence of feeding sheep on oilseed cake following the consumption of tanniferous feeds. Livest. Prod. Sci. 60, 59-69. [ Links ]
Pell, A.N. & Schofield, P., 1993. Computerized monitoring of gas production to measure forage digestion in vitro. J. Dairy Sci. 76, 1063-1073. [ Links ]
SAS, 2000. Statistical Analysis System user's guide (Version 8). SAS Institute Inc., SAS Campus Drive, Cary, N.C., USA. [ Links ]
Scalbert, A., 1991. Antimicrobial properties of tannins. Phytochemistry. Oxford, Pergamon Press, UK. 30 (12) pp. 3875-3883. [ Links ]
Seyis, I. & Aksoz, N., 2005. Investigation of factors affecting xylanase activity from Trichoderma harzianum 1073 D3. Braz. Arch. Bio. Technol. 48, 187-193. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. See: http://creativecommons.org/licenses/by/2.5/za
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science
# Corresponding address: nsahlaii@ukzn.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}