










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 n.3 Pretoria Jan. 2011
SHORT COMMUNICATION
Parentage verification of South African Angora goats, using microsatellite markers
C. Visser; E. van Marle-Köster*; H. Friedrich
Department of Animal and Wildlife Sciences, University of Pretoria, Pretoria 0002, South Africa
ABSTRACT
South African Angora goats are farmed under extensive production systems in relatively large herds. As a result, breeders make use of group and flock-mating systems that limit accurate parentage recording and selection efficiency. In this study the aim was to refine a panel of microsatellite markers suitable for parentage verification in South African Angora goats. The markers were first evaluated based on the number of alleles, allele frequency, PIC, HE, HO and individual exclusion probability, and secondly as part of a panel. Eighteen markers were tested in 192 South African Angora goats representing different family structures with known and unknown parent information. The final set of microsatellite markers, with the strongest exclusion and the least number of microsatellite markers, consisted of 14 microsatellite markers namely BM1258, BM1329, BM1818, BM7160, CSRD247, HSC, INRA63, INRABERN192, MCM527, OarFCB48, SRCRSP5, SRCRSP8, SRCRSP9 and SRCRSP24. This panel had a combined first-parent exclusion probability of 99.7% and it was possible to perform parental identification in a test family.
Keywords: Pedigree allocation, DNA technology, exclusion probability
South African Angora goats are primarily farmed under extensive production systems and herd sizes vary from 1000 to 2500 goats per herd. Most breeders have both a stud and commercial herd. Mating systems used by breeders taking part in the National Small Stock Improvement Scheme primarily include group mating and flock mating, while commercial breeders also make use of over-mating (combining group-mating with a complete flock-mating at the end of the breeding season). These mating practices limit accurate pedigree recording. It has been estimated that of the Angora kids born between 2000 and 2005, 23% had incomplete or inaccurate pedigree data, with unknown sires posing the main limitation (Snyman, 2010, Grootfontein Agricultural Development Institute, Jansenville, Private Bag X529, Middelburg, Eastern Cape, 5900, South Africa.). Other factors that contribute to potential errors in identification of the parents include the use of large paddocks in extensive production systems, which have also been reported in similar studies (Bolormaa et al., 2008). Angora ewes are known to abandon kids if they feel threatened during parturition (Hafez & Hafez, 2000), leading to incorrect maternal allocation. Inaccurate parentage recording over time results in lower selection efficiency due to mating based on incorrect pedigree data (Pollak, 2005).
The South African Angora goat industry is currently the world leader in mohair production with exports of 3.5 million kg of mohair annually (Van der Westhuysen, 2005; Retief, 2008). In order to maintain a high quality clip it is essential for breeders to be able to select the best parents for mating from accurate pedigree data and breeding value information. DNA-based parentage testing can therefore play an important role in improving the efficiency of selection.
A large number of microsatellite markers are available for most farm animal species and parentage panels have been commercialized and used in the routine testing of dairy and beef cattle (Van Eenennaam et al., 2007; Van de Goor et al., 2009). The goat genome has not been studied as widely when compared to other farm animal species (Fadiel et al., 2005; Maddox & Cockett, 2007) with approximately 420 microsatellite markers available in the INRA Goatmap database (http://dga.jouy.inra.fr/cgibin/lgbc/summary.operl?BASE=goat). The International Society of Animal Genetics (ISAG) recommended 18 microsatellite markers for parentage verification in goats during 2001/2002 and 2005, respectively (http://www.isag.org.uk/Docs/2005_PanelsMarkersSheepGoats.pdf). These markers were tested in several laboratories and results generally compared well with regard to individual marker performance. However, the test results of these marker panels indicated a wide variation in polymorphicity and heterozygosity levels between different goat breeds. These parameters have a direct impact on the combined exclusion probabilities of the panels and should therefore be tested in each specific population. Studies were also conducted by Luikart et al. (1999), Ganai & Yadav (2005), Glowatzki-Mullis et al. (2007), Bolormaa et al. (2008) and De Araujo et al. (2010) who added additional microsatellite markers to the ISAG panels for parentage verification in specific goat breeds. In this study the aim was to refine a panel of microsatellite markers suitable for parentage verification in South African Angora goats.
Blood samples (5 mL) were annually collected in EDTA tubes from a number of Angora stud herds participating in the Small-stock Bio-bank at Grootfontein Agricultural Development Institute (GADI) in Middelburg, Eastern Cape, South Africa. Blood was stored at -40 ºC until DNA extraction with a Roche DNA Isolation kit for Cells and Tissues (Roche Applied Sciences) at GADI. DNA samples of 192 South African Angora goats originating from six different herds were provided by the Bio-bank and quantified using a Nanodrop ND-1000 UV-vis Spectrophotometer (http://www.nanodrop.com) at the Department of Genetics, University of Pretoria. Most animals were unrelated, with no more than 10 half-sibs included per herd. This population was used to estimate the individual and panel parameters for inclusion of the markers into a verification panel.
Microsatellite markers were selected from the panels recommended by ISAG, as well as markers used in previous parentage verification studies on goats. Thirteen markers were selected from the ISAG panels. The remaining ISAG-recommended markers performed poorly in previous studies on SA Angora goats, with low polymorphicity values and/or low amplification success. Five alternative markers were selected based on the usage in other studies and performance in the SA Angora goat population (Visser & Van Marle-Köster, 2009; Visser et al., 2010). Parameters for inclusion in the current test panel included previously reported levels of polymorphism, heterozygosity, null allele frequencies and fragment sizes. The 18 markers were grouped into two genotyping sets according to range and fluorescent labelling for cost-efficient genotyping.
Individual PCRs were performed for each microsatellite. The PCR was carried out in a 15 µL reaction, containing 5 µL of DNA template with a concentration of 100 ng/µL, 0.3 µL of 10 pmol/µL each of the reverse and forward primer, 0.75 µL 0.25 mM MgCl2, 0.3 µL 10 nM dNTP's, 0.4 µL 1.5 U Taq, 1.5 µL 5 x Colorless GoTaq Flexi Buffer (Promega - Whitehead Scientific Inc. South Africa) and 6.45 µL deionized water. The amplification was performed using a GeneAmp® PCR System 9700 thermocycler (Applied Biosystems, Foster City, USA) using the following PCR programme: 10 min at 94 ºC, followed by 33 cycles of 45 sec at 94 ºC, 80 sec at the annealing temperature and 60 sec at 72 ºC and an extension step of 5 min at 72 ºC. PCR products were diluted with distilled water to 1:10 and prepared for genotying on an ABI PRISM ® 3100 DNA Genetic Analyzer (Applied Biosystems, Foster City, USA). Fragment sizes were determined using GenemapperTM software (Applied Biosystems, Foster City, USA).
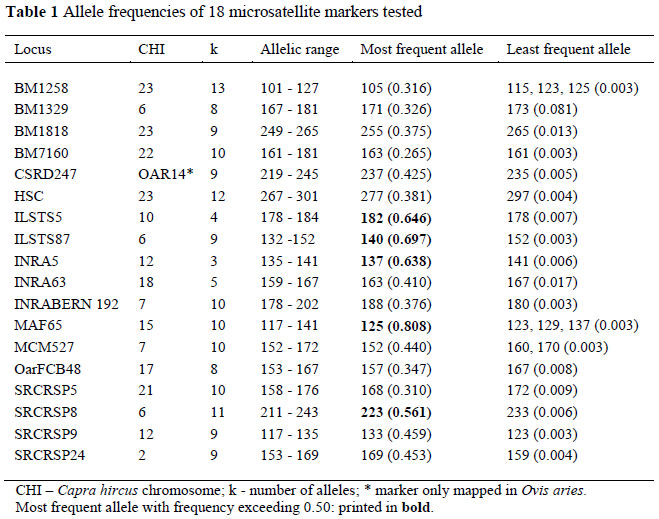
Allele frequencies, Polymorhic Information Content (PIC) and Observed and Expected Heterozygosity (HO and HE) values were calculated using a Microsatellite toolkit (Park, 2001). Parameters for parentage verification included Hardy-Weinberg equilibrium (HWE) using Genepop version 4.0.10 (Raymond & Rousset, 1995), Null allele (FNull) frequency and exclusion probabilities (PE1, PE2) that were calculated using Cervus 3.0 (Marshall et al., 1998) for each locus. In order to compile a panel with the highest potential exclusion power, based on the combined performance of the markers, the Combined Probability of Exclusion (CPE) for a given set of loci was calculated several times, excluding markers with poorer performance sequentially. The panel was verified in a test family consisting of 11 kids, 11 does and 12 possible sires. The criteria for interpretation of the LOD scores were based on Slate et al. (2000). A total of 18 markers, tested on 192 Angora goats, were analysed for their suitability for parentage verification. The markers were found to be polymorphic with an average of 8.8 alleles per microsatellite marker, ranging between three alleles (INRA5) and 13 alleles (BM1258). The number of alleles detected for the microsatellite markers were similar to values reported by Luikart et al. (1999), Ganai & Yadav (2005), Jimenez-Gamero et al. (2006), Glowatzki-Mullis et al. (2007) and Bolormaa et al. (2008). Four of the 18 microsatellite markers (MAF65, ILSTS87, ILSTS5 and INRA5) analyzed each had an allele with a relatively high frequency (0.64 to 0.82) that affected their effective polymorphism, as shown in Table 1. The high frequency (exceeding 0.60) of one allele for these markers was undesirable. As indicated by Marshall et al. (1998), markers with specific alleles with frequencies exceeding 0.5 are generally not suited for inclusion in a parentage verification panel. These high frequency alleles negatively affect the markers' PIC, HE, and HO values. It should be noted that the markers might perform at differing levels in other populations where the allele frequencies might be different.
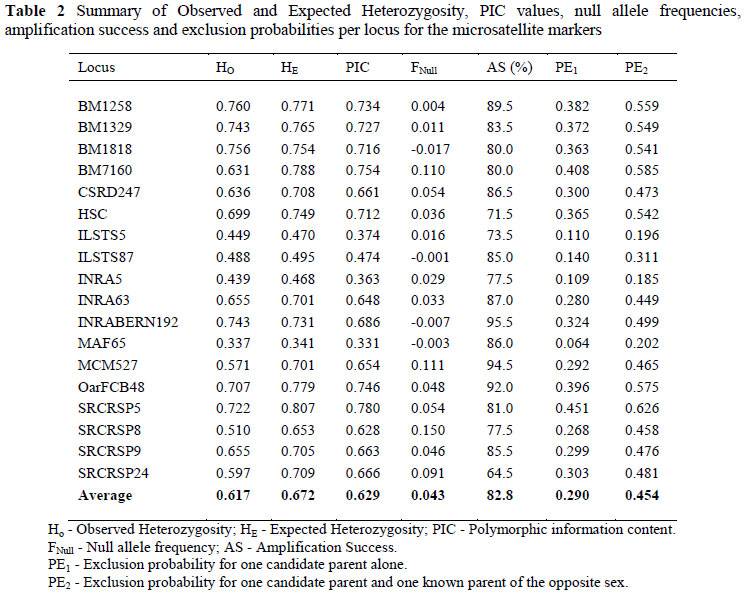
In Table 2 a summary is provided of the performance of the 18 markers for heterozygosity and polymorphicity values, as well as the occurrence of null alleles and the amplification success of individual markers. All the markers were polymorphic and 14 markers showed heterozygosity (HE and HO) and PIC values above 50%. The HE values ranged from 0.341 (MAF65) to 0.807 (SRCRSP5). HO values varied between 0.337 (MAF65) and 0.76 (BM1258). PIC values ranged between 0.331 for MAF65 and 0.780 for SRCRSP5 with 13 markers having PIC values above 0.650. The PIC estimates for SRCRSP9 reported by both Luikart et al. (1999) (0.812) and Jimenez-Gamero et al. (2006) (0.781) were higher than found in the current study (0.663). The PIC value estimated for MAF65 (0.339) was much lower than that found in the study by Luikart et al. (1999) at 0.671.
The Null allele frequency ranged from -0.017 (BM 1818) to 0.150 (SRCRSP 8) with an average of 0.043 (Table 2). Two of the markers' values were marginally over 0.05, while three markers (BM7160, MCM527 and SRCRSP8) had values exceeding 0.1. Markers with null allele frequencies exceeding 0.05 are generally not suitable for inclusion in parentage verification panels (Marshall et al., 1998) as they tend to have heterozygote deficiencies.
All loci were tested to conclude if the markers were in HWE in the population. Three of the 18 microsatellite markers (BM1818, SRCRSP5 andBM1329) showed significant (P <0.002) departure from HWE following Bonferroni correction.
Three of the 18 microsatellite markers had an amplification success of less than 75%. INRABERN 192 amplified the best with an amplification rate of over 95% whereas SRCRSP 24 had only a 64.5% success rate (Table 2).
The exclusion probabilities, as shown in Table 2, are values based on the allele frequencies of each of the markers alone, and can thus be computed in any family structure. PE1 is the exclusion probability of each microsatellite marker when the genotypes of the alleged parent (most often the sire) and the offspring are known. PE2 differs from PE1 in that the alleged parent (most often the sire), offspring, and the known parent's (most often the dam) genotypes are known. Markers ILSTS5, ILSTS87, INRA5, INRA63, MAF65 and SRCRSP8 performed below average for the individual parameters estimated.
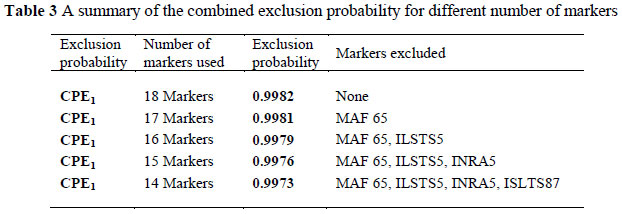
In order to construct an economically-feasible panel of microsatellite markers with the highest exclusion probability and no parental allocation errors, markers that did not perform optimally were excluded from the panel. MAF65, INRA5, ILSTS5 and ILSTS87 were excluded based on their allele frequencies, number of alleles, heterozygosity values and PIC values. To estimate the impact that the inclusion or exclusion of these markers would have on a parentage verification panel, the CPE was calculated several times, excluding markers with poorer performance sequentially. Combined Probability of Exclusion1 (CPE1) decreased from 0.998 to 0.997 when the markers included in the panel were decreased from 18 to 14, as shown in Table 3.
The CPE values are influenced not only by the number of markers included, but also by the genotypes available from both known and candidate parents (Gerber et al., 2000). In cases where genotypes of both parents are available, the exclusion power will increase the statistical probability to exclude certain non-parents. Breeders, however, (often due to financial reasons) usually only submit samples of the sire and offspring. Parentage panels should therefore have sufficient power for providing accurate results with less information available. In this study the CPE1 decreased by 0.1% when the four markers with the poorest performance were excluded. The final set of microsatellite markers with the strongest exclusion power and the least number of microsatellite markers consisted of 14 microsatellite markers, namely BM1258, BM1329, BM1818, BM7160, CSRD247, HSC, INRA63, INRABERN192, MCM527, OarFCB48, SRCRSP5, SRCRSP8, SRCRSP9 and SRCRSP24. The CPE1 of this panel was 99.7%. This value corresponded closely to those reported in previous parentage verification studies on goats, which were above or very close to 99% (Luikart et al., 1999; Ganai & Yadav, 2005; Jimenez-Gamero et al., 2006; Glowatzki-Mullis et al., 2007; Bolormaa et al., 2008).
These markers were combined into two sets for cost-effective genotyping and were compiled according to size range and fluorescent labels. The panel was verified in a small family of 11 offspring with maternal and paternal genotypes. Parental identification could be conclusively performed for 10 of the progeny. For these animals no mismatches were recorded with LOD scores above +3.0. It was possible to assign eight of the 11 offspring to the candidate sires at a 95% level of confidence, two at 80% confidence and 1 offspring could not be allocated conclusively.
In this study microsatellite markers were evaluated for a number of criteria to test suitability for parentage analyses. A panel of 14 markers was shown to be effective for parentage assignment in South African Angora goats. DNA-based parentage therefore has the potential to assist SA Angora breeders to improve pedigree recording and selection accuracy, resulting in an increase in the rate of genetic improvement.
References
Bolormaa, S., Ruvinsky, A., Walkden-Brown, S. & Van der Werf, J., 2008. DNA-based parentage verification in two Australian goat herds. Small Rumin. Res. 80, 95-100. [ Links ]
De Araujo, A.M., Guimaraes, S.E.F., Pereira, C.S., Lopes, P.S., Rodrigues, M.T. & Machado, T.M.M., 2010. Paternity in Brazilian goats through the use of DNA microsatellites. Revista Brasileira de Zootecnia. 39, 1011-1014. [ Links ]
Dodds, K.G., McEwan, J.C. & Davis, G.H., 2007. Integration of molecular and quantitative information in sheep and goat industry breeding programmes. Small Rumin. Res. 70, 32-41. [ Links ]
Fadiel, A, Anidi, I. & Eichenbaum, K.D., 2005. Farm animal genomics and informatics: an update. Nucleic Acids Res. 33, 19. [ Links ]
Ganai, N.A. & Yadav, B.R., 2005. Parentage determination in three breeds of Indian goat using heterologous microsatellite markers. In: Applications of Gene-based Technologies for Improving Animal Production and Health in Developing Countries, 613 - 620. IAEA. ISBN: 978-1-4020-3311-7. [ Links ]
Gerber, S., Mariette, S., Streiff, R., Bodenes, C. & Kremer, A., 2000. Comparison of microsatellites and amplified fragment length polymorphism markers for parentage analysis. Mol. Ecol. 9, 1037-1048. [ Links ]
Glowatzki-Mullis, M.L., Muntwyler, J. & Gaillard, C., 2007. Cost-effective parentage verification with 17-plex PCR for goats and 19-plex PCR for sheep. Anim. Genet. 38, 86-88. [ Links ]
Hafez, B. & Hafez, E.S.E., 2000. Reproduction in Farm Animals. Blackwell Publishing, The Atrium, Southern Gate, Chichester, West Sussex, PO 19 8 SQ, UK. 7th edition. ISBN 0683305778, 9780683305777. [ Links ]
Jimenez-Gamero, I., Dorado, G., Munoz-Serrano, A., Analla, M. & Alonso-Moraga, A., 2006. DNA microsatellites to ascertain pedigree-recorded information in a selecting nucleus of Murciano-Granadina dairy goats. Small Rumin. Res. 65, 266-273. [ Links ]
Luikart, G., Biju-Duval, M-P., Ertugrul, O., Zagdsuren, Y., Maudet, C. & Taberlet, P., 1999. Power of 22 microsatellite markers in fluorescent multiplexes for parentage testing in goats (Capra hircus). Anim. Genet. 30, 431-438. [ Links ]
Maddox, J.F. & Cockett, N.E., 2007. An update on sheep and goat linkage maps and other genomic resources. Small Rumin. Res. 70, 4-20. [ Links ]
Marshall, T.C., Slate, J., Kruuk, L.E.B. & Pemberton, J.M., 1998. Statistical confidence for likelihood- based paternity inference in natural populations. Mol. Ecol. 7, 639-655. [ Links ]
Park, S.D.E., 2001. Trypanotolerance in West African cattle and the population genetic effects of selection. Ph.D. thesis. University of Dublin, Ireland. [ Links ]
Pollak, E.J., 2005. Application and impact of new genetic technologies on beef cattle breeding: a "real world" perspective. Aust. J. Exp. Agr. 45, 739-748. [ Links ]
Retief, L., 2008. Sybokhaarkwekersvereniging: Jaarverslag. The Angora Goat and Mohair Journal - Spring 2008. [ Links ]
Raymond, M. & Rousset, F., 1995. GENEPOP (version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 86, 248-249. [ Links ]
Slate, J., Marshall, T. & Pemberton, J., 2000. A retrospective assessment of the accuracy of the paternity inference program CERVUS. Mol. Ecol. 9, 801-808. [ Links ]
Van de Goor, L.H.P., Panneman, H. & Van Haaeringen, W.A., 2009. A proposal for standardization in forensic bovine DNA typing: allele nomenclature of 16 cattle-specific short tandem repeat loci. Anim. Genet. 40, 630-636. [ Links ]
Van der Westhuysen, J.M., 2005. Marketing goat fibres. Small Rumin. Res. 60, 215-218. [ Links ]
Van Eenennaam, A.L., Weaber, R.L., Drake, D.J., Penedo, M.C.T., Quaas, R.L., Garrick, D.J. & Pollak, E.J., 2007. DNA-based paternity analysis and genetic evaluation in a large, commercial cattle ranch setting. J. Anim. Sci. 85, 3159-3169. [ Links ]
Visser, C. & Van Marle-Köster, E., 2009. Genetic variation of the reference population for quantitative loci research in South African Angora goats. AGRI 45, 113-119. [ Links ]
Visser, C., Crooijmans, R.P.M.A. & Van Marle-Köster, E., 2010. A genetic linkage map for the South African Angora goat. Small Rumin. Res. 93, 171-179. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence.
See: http://creativecommons.org/licenses/by/2.5/za
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science
* Corresponding author: carina.visser@up.ac.za


{kind=link}
{kind=link}
{kind=link}