










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 n.3 Pretoria Jan. 2011
Effect of dietary protein on the allometric relationships between some carcass portions and body protein in three broiler strains
R. Danisman; R.M. Gous*
Animal and Poultry Science, University of KwaZulu-Natal, Pietermaritzburg 3209, South Africa
ABSTRACT
This paper reports the allometric relationships between some of the physical parts and body protein weight of three commercial broiler strains reared, sexes separate, on four dietary protein levels to six weeks of age. Birds were sampled at day old and then weekly from each of the treatments to determine the weights of the physical parts and the chemical composition of each of 582 birds. Allometric regressions were compared between strains, sexes and dietary protein levels using linear regression with groups. Whereas these regressions were similar over strains and sexes, some interactions were evident between factors, and differences occurred when broilers were fed differing dietary protein levels. These differences may be explained on the basis that lipid is deposited to different extents in each of the parts in response to dietary protein, although this was not tested. The allometric regressions presented are an attempt to provide information that would enable the prediction of the weights of breast meat, thigh, drum and wing at different stages of growth of broilers whose genotype and feed composition are adequately described.
Keywords: Breast meat, thigh, drum, wing, carcass yield, simulation modelling
Introduction
The increasing consumer demand for breast meat has resulted in a search for ways of producing chickens that yield greater amounts of this desirable body component, the two most important approaches being through genetic selection and nutrition. Broilers have been subjected to selection for many decades, and geneticists have in that time considerably improved traits such as body weight gain, feed conversion efficiency and carcass characteristics (Havenstein et al., 2003; McKay, 2008). Many reports claim that improvements have been made in the yield of parts (breast meat in particular) of broilers, with resultant 'high yield' strains having been developed by some breeding companies (Cobb, 2007; Aviagen, 2009). Similarly, there are numerous claims that breast meat yield can be improved by altering dietary nutrient specifications: examples are the comprehensive dataset published by Hakansson et al. (1978) and the effects of dietary protein content on breast meat yield (Kemp et al., 2005). These improvements raise a general problem in growth analysis, which is to be able to predict the effects of genotype, feed and environment on the growth of protein and lipid in breast meat and other body components.
Such predictions may be accomplished directly, using a growth function specific to each component, or indirectly, by predicting the growth of the component as a proportion of body protein growth. Potential protein growth rates have been described for various genotypes (Hancock et al., 1995; Gous et al., 1996) using a Gompertz growth function (Gous et al., 1999) and simulation models already exist for predicting the actual growth of body protein as influenced by genotype, feed and environment (Emmans, 1981; EFG Software, 2007). Such models may therefore be used to predict the growth of any scaled component of the body, as any components that share the same rate of maturing can be scaled allometrically (Emmans, 1987; 1988). What is needed for each genotype is a description of the potential rates of growth of body protein and of the different components of the body so that the allometric relationships between the important components of the body and body protein can be determined. Differences in the allometric relationships between genotypes, or those brought about through nutritional or environmental manipulation would enable these effects to be modelled.
The proportions of the body change in a regular fashion throughout growth, and such non-isometric scaling is called allometry. Many morphological and physiological variables are scaled, relative to body size, according to allometric equations of the general form y = axb. In a simple proportionality the slope (b) will be 1.0, as with the weight of blood in relation to body mass, or the amount of ash relative to the weight of body protein (Schmidt-Nielsen, 1984; Emmans, 1987). If the dependent variable increases at a slower rate than would be indicated by simple proportionality the regression line would have a slope less than 1.0, such as with metabolic rate (0.75 of body weight) and body water (0.9 of body protein weight). Body lipid content would generally increase at a faster rate than body protein content, and hence the slope would be greater than 1.0. Where the quantities do not change with body size, such as the haemoglobin concentration in the blood, the slope is zero. Some functions even decrease with body size: the slope for heart rate vs. body size, for example, is -0.25. Knowledge of these relationships would be invaluable for predicting the weights of the commercially important physical components of a broiler, as it means that only the weight of body protein needs to be predicted, after which the weights of all the remaining chemical and physical components of the body can be calculated using the appropriate allometric coefficients. However, no such relationships for broilers have been published in the scientific literature nor is it possible to determine such relationships accurately using the data that have been published.
Where mean weights have been published of the physical components of broilers resulting from changes in genotype, feed quality or the environment, a rough estimate of the allometric relationships with body weight can be obtained, but this is inaccurate for two reasons: firstly, because of variation between individuals on the same treatment the regressions would be more accurate if the individual results were used instead of treatment means; and secondly, because the treatments applied often influence the amount of lipid in the body, it is more accurate to regress component weights against body protein weight rather than on body weight thereby eliminating the confounding effect of body lipid content. This implies that the body protein content should be measured of each bird sampled for the measurement of body component weights.
In the trial reported here, the weights of various parts of the body were measured at different stages of growth after which all components of each bird were minced together in order to determine the chemical composition of each bird. With the information about the weights of the components and of the protein content of each bird it was possible to determine the allometric relationships between the components and body protein, and to compare these between genotypes and feed protein levels. Apart from similar exercises having been conducted previously in our laboratory (Lindsay, 1992: Gous et al., 1996) no other such data exist for comparison.
This study was designed to examine the relationships between some important components of the body and body protein among three broiler genotypes and two sexes fed four protein levels over a period of six weeks to ascertain whether differences in these allometric relationships exist.
Materials and Methods
Day-old sexed broiler chickens of three strains available in South Africa, namely, Ross 788, Ross 308 and Cobb 500, were used in the trial. A total of 48 pens was used with 70 chickens being placed initially per pen and with males and females being reared separately, i.e. eight pens of males and eight of females of each of three strains were used in the trial. The trial was terminated when the birds were 42 d of age.
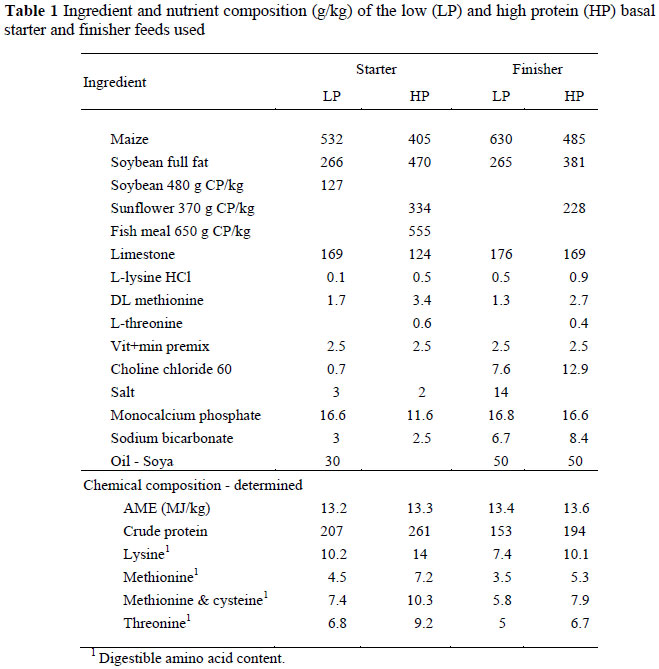
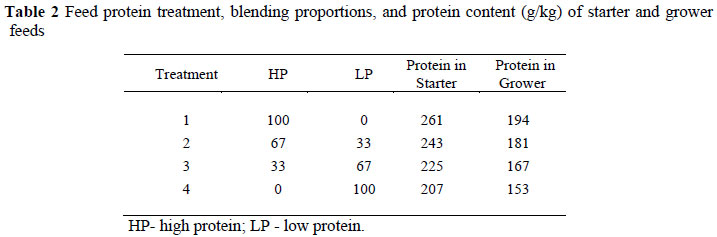
Two basal starter and finisher feeds, one high and the other low in protein were formulated (Table 1) to contain equal contents of apparent metabolisable energy (AME) and major minerals, using a well balanced amino acid mixture derived from the EFG Broiler Growth Model (EFG Software, 2007). These two feeds were blended to produce two intermediate levels of protein (Table 2). The basal feeds were sampled after mixing and these were analysed for AME using the method of Fisher & McNab (1987) and digestible amino acid contents using a Waters amino acid analyser (AOAC, 2003). The starter feeds were fed to three weeks of age and the finisher feeds were used thereafter. Feed and water were offered ad libitum throughout the trial.
A representative sample (0.5) of birds in each pen was weighed at weekly intervals up to 42 d. Feed allocated to each pen was weighed and transferred to the feeding troughs when necessary. At the end of each week feed remaining in the troughs and in the bag was weighed, to determine the amount of feed consumed during the week.
At day old, six birds were sampled from each strain x sex, the weights of the parts were dissected and measured then replaced before each chick from each strain and sex was minced and subjected to chemical analysis. Two birds were removed from each pen for carcass analysis each week up to the 6th week, resulting in a total of 6 (initial) + 96 x 6 = 582 birds being used for carcass analysis. The weight of each bird was recorded after killing (without loss of blood), plucked weight was recorded (feather weight calculated by difference), and then the breast, without skin or bone, breast skin, breast bone, thighs, drums, wings, intestine, liver, and remainder were dissected and weighed. All the body parts, except for feathers, from each bird were placed together in a separate plastic bag, clearly marked with the identity of the bird, and stored in a freezer until mincing took place. Each chicken was minced separately and sampled for chemical analysis which included water, protein (as N using the Dumas method on a LECO N analyser, AOAC, 2003), ash (AOAC, 2003), and gross energy (GE) using a bomb calorimeter. Lipid (L) content was calculated from GE using the equation L = -0.8756 + 0.04754*GE (University of KwaZulu-Natal, unpublished).
The weight and natural log (ln) of each chemical component of the body was calculated, from which the allometric relationships between the components (Y) and body protein weight (X) were determined using the equation ln Y = ln a + b ln X. The exponent b represents the slope of the linear regression obtained in a logarithmic plot.
A factorial design was used, with three protein levels, four strains and two sexes. Two replications of each treatment combination were used. Data were subjected to statistical analysis using analysis of variance for calculation of treatment means only, and simple linear regression of the form lnY = a +b lnX using natural logarithms (ln) of both the component (Y) and body protein (X) weights to determine the allometric coefficients. The groups procedure of Genstat (2002) was applied to determine differences between constant terms (a) and slopes (b) of the regressions. Main effects and two- and three-way interactions between factors were compared in this way.
Results
The objective of the trial was not to determine the extent to which broiler genotypes differed, or the extent to which dietary protein content would alter growth rate and feed conversion efficiency, but rather to produce a wide range of body weights and carcass compositions throughout the growing period such that the allometric relationships between the physical parts of broilers and their body protein contents could be determined. Therefore, although the body weights and food intakes of broilers on each of the treatments were recorded each week, only the mean final body weight and feed conversion efficiency (FCE) for each strain, sex and balanced protein level are presented here (Table 3) to indicate the extent to which these varied between birds on the various treatments. In all cases, males were heavier than females, and dietary protein content influenced growth rate, with the highest body weight being recorded on the second-highest feed protein level. Food intake was maximised on the second-highest feed protein level, declining at higher and lower levels, whereas FCE decreased linearly with dietary protein content, the highest (600 g gain/kg food) being on the highest protein feed.
Also presented in Table 3 are the mean body lipid contents of the broilers sampled for carcass analysis at 42 d of age. Females had higher (P <0.001) lipid contents than males (158 vs. 107 g/kg) and there was a linear increase in lipid content as dietary protein content declined. The mean values presented here demonstrate that the factors used, namely, strain, sex and feed protein content, produced differences in the body weights and feed efficiencies of the broilers, and because feed efficiencies differed, so too did the body composition of birds on the different protein treatments.
Of the components of the body that were measured, only the weights of breast meat, thigh, drum and wing are given in the tables below. The mean weights of the remaining five components measured, namely, breast skin and bone, liver, gut and the remainder of the carcass are not presented in this paper, nor are the results of the chemical analyses of the carcasses other than the lipid content at six weeks. The entire dataset is available from the corresponding author on request.
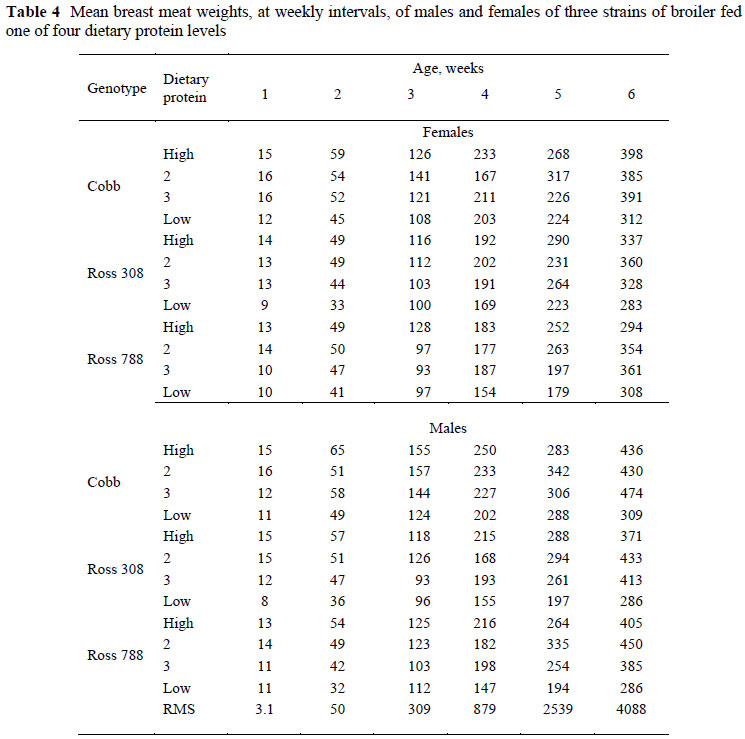
The mean breast meat weights, at weekly intervals, of males and females of three strains of broiler fed four dietary protein levels are given in Table 4. The weights refer to the entire breast meat and exclude skin and bone. Over all ages the weights were heavier in males than in females (390 vs. 343 g at 42 d), heavier in the Cobb strain than in Ross 308 or 788 (392, 351 and 355 g, respectively at 42 d), and dietary protein content resulted in a curvilinear response, with the highest weight of breast meat on protein level 2 (374, 402, 392 and 297 g, respectively at 42 d).
Data for breast meat and body protein weight from each bird sampled were used to calculate allometric equations for each strain, sex and feed protein level, which were compared using simple linear regression with groups in Genstat, the respective coefficients for the main effects of the three factors being presented in Table 5. Where no differences were apparent in either the constant term or regression coefficient the common value is presented, as with those between strains and sexes, where only one constant term (-2.379) and regression coefficient (1.409) is necessary to describe breast meat weight in terms of body protein content in all three strains and both sexes. The only differences in allometric coefficients between factors for breast meat were those brought about by feeding different levels of dietary protein, which resulted in linear trends in both constant terms (decreasing from -1.214 on the highest protein level to -1.536 on the lowest) and regression coefficients, which increased from 1.191 to 1.237.
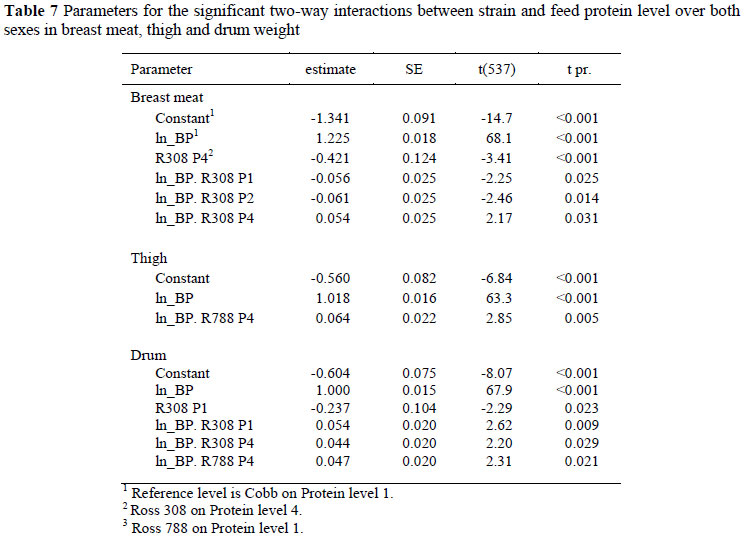
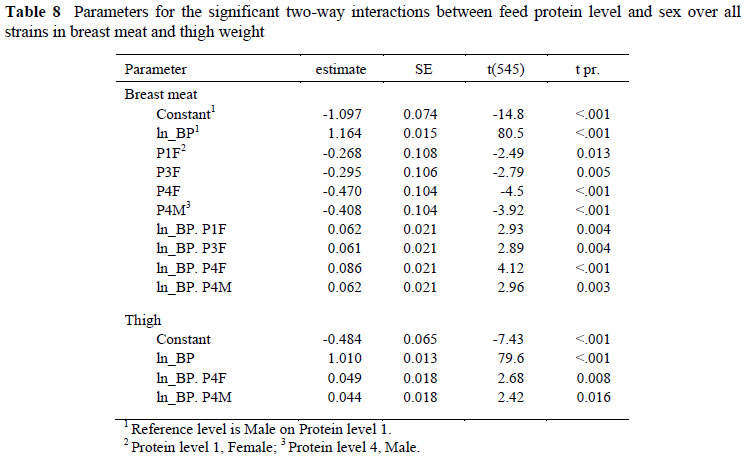
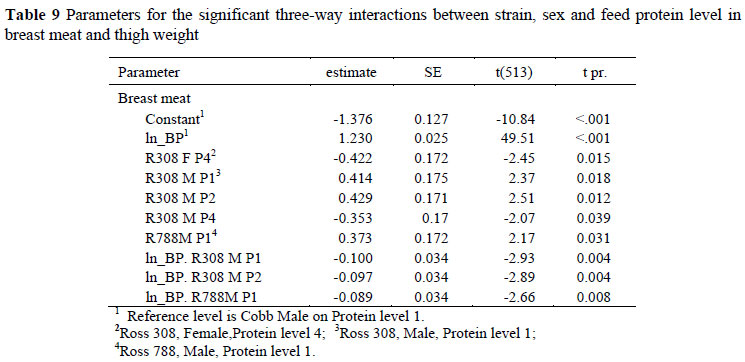
Significant two-way interactions in the allometric coefficients for breast meat yield occurred between strain and sex (Table 6) with Ross 308 and 788 males differing from Cobb males and all females. The constant term for Ross males was between 0.182 ± 0.090 (Ross 788) and 0.196 ± 0.090 (Ross 308) higher, and the regression coefficient 0.054 and 0.058 ± 0.018 lower than for the Cobb males, indicating that the Ross strain had higher breast meat weights in the early stages of growth but with the Cobb strain overtaking this advantage at some stage before the end of the growing period. Interactions also occurred between strain and feed (Table 7) where the Ross 308 strain on protein level 4, for example, had a lower constant term (-0.421 ± 0.124) and a higher regression coefficient (0.054 ± 0.025) than the Cobb strain on the highest protein level, and between feed protein level and sex (Table 8). An example of the latter interaction is that between males and females on the highest protein level, where females had a lower constant term (-0.268 ± 0.108) and a higher regression coefficient (0.062 ± 0.021). The three way interactions between strain, sex and dietary protein levels are given in Table 9. No significant trends were evident but Ross 308 appeared to differ more than the Ross 788 from the Cobb strain.
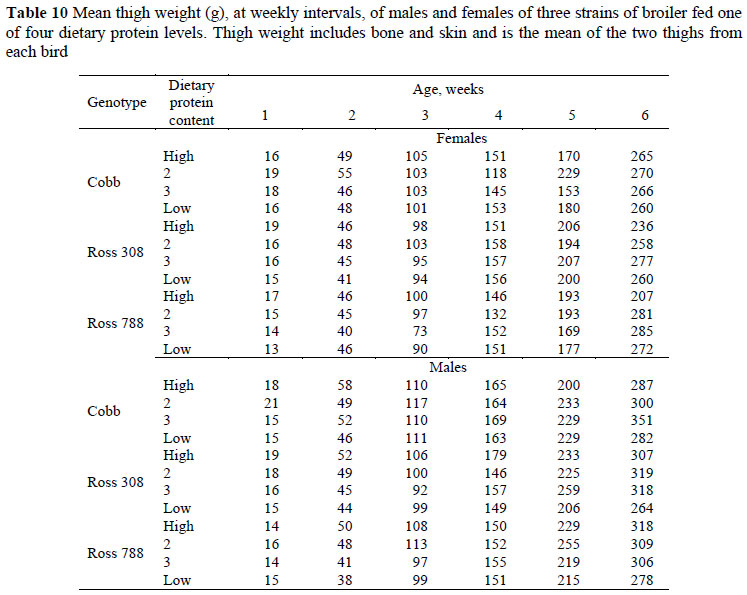
The mean thigh weights of broilers on the various treatments, at weekly intervals, are given in Table 10. The weight referred to here includes bone and skin and is the mean of the two thighs from each bird. At six weeks the mean thigh weights of the three strains over all protein levels were similar (285, 282 and 280 g for Cobb, Ross 308 and 788, respectively), whereas the mean weight of male thighs (303 g) was 42 g heavier than those of females. A curvilinear response in thigh weight to dietary protein level was evident, with mean weights at six weeks of 270, 290, 300 and 270 g, respectively. The main effect of strain on the allometric relationship between thigh weight and body protein weight (Table 5) resulted in significant differences in constant terms between the Cobb strain vs. the two Ross strains but all having the same regression coefficient, i.e. the rate at which thigh weight increased with body protein content was the same in all strains, but the initial weight of the Cobb was higher, and this difference therefore was maintained throughout the growing period. Both the constant term and regression coefficient were the same for both sexes, but the constant term, whilst the same for the three highest protein levels, was significantly lower for the lowest protein level whilst the regression coefficient for this treatment was steeper than for the other protein levels.
There were no two-way interactions between strain and sex in the case of thigh weight, but feed protein level interacted with both strain (Table 7) and sex (Table 8). The interaction with strain occurred in one case only, with the regression coefficient for the Ross 788 fed the lowest feed protein level being 0.064 ± 0.022 higher than all other combinations of strain and feed protein content. The interaction with sex was brought about also at the lowest feed protein content, with a higher regression coefficient being exhibited by females (0.049 ± 0.018), and males (0.044 ± 0.018) on protein level 4. No three-way interactions were evident for thigh weight.
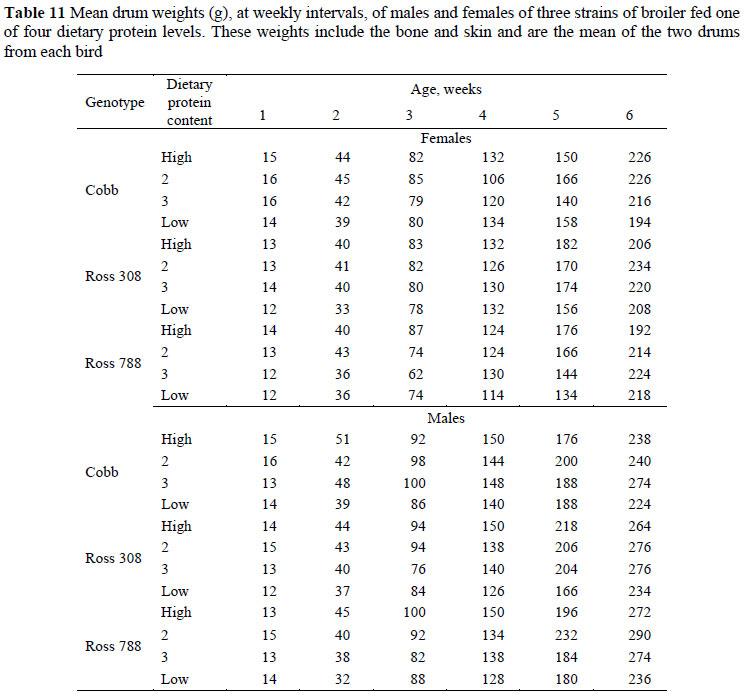
The mean weights of the drum of broilers on the various treatments, at weekly intervals, are given in Table 11. These weights include bone and skin and represent the mean weights of the two drums from each bird. At six weeks the mean drum weight of the Cobb strain, over all protein levels, was lighter (230 g) than that of the Ross strains, which were both 240 g. Mean weight of male drums (129 g) was 22 g heavier than that of females. A curvilinear response in drum weight to dietary protein level was evident, with mean weights at six weeks of 233, 247, 247 and 220 g, respectively.
Two-way interactions were evident between strain and sex (Table 6) and between strain and feed protein level (Table 7), but not between sex and feed protein level. Females of the Ross 308 strain differed from all other strains and sexes by having a lower constant term (-0.174 ± 0.058) and a higher regression coefficient (0.040 ± 0.012) than the other combinations. This strain also responded differently to the highest and lowest feed protein levels, in the first case showing the same trend as the Ross 308 females (lower constant term and higher regression coefficient), and, in the latter case, together with the Ross 788 strain, showing a higher regression coefficient on the lowest dietary protein level. There were no three-way interactions in drum weight between factors.
The mean weights of the wing of broilers on the various treatments, at weekly intervals, are given in Table 12. These weights include bone and skin and represent the mean weights of the two wings from each bird. Male wings were heavier than those of females at six weeks (201 vs. 174 g) and those of all three strains were similar (185, 185 and 193 g for Cobb, Ross 308 and 788, respectively). Weights were curvilinearly related to dietary protein level (186, 194, 195 and 175 g, respectively). There were no two- or three-way interactions between factors in the case of mean wing weight.
Discussion
The three strains, two sexes and four dietary protein levels used in the trial had the desired effect of producing a wide range of rates of growth of the body and of the weights of the physical and chemical components of the birds sampled. The body weights achieved exceeded the breed standards for the three strains used. In addition, because a large number of birds (n = 582) was sampled throughout the growing period, the resultant allometric equations derived for each treatment combination are likely to be accurate representations of the effects of the various treatments imposed. The objective was to derive and then compare these allometric equations between the weights of the various physical parts and body protein, and this was accomplished using simple linear regression with groups; the groups in this case being the factors strain, sex and feed protein level. Because the constant terms and slopes of the regressions were being compared, and not the mean values at different stages of growth, it is not appropriate to use an analysis of variance or t- tests to compare these relationships (Morris, 1983; Gous, 2010).
A number of interesting effects were noted in the relationship between breast meat and body protein content. In spite of differences in growth rates and carcass composition between the strains and sexes used in the trial, breast meat weight was unaffected by these factors when expressed as a proportion of the body protein content of the broiler. However, differences in the constant term and regression coefficient describing this relationship were brought about by rearing the birds on different levels of dietary protein, with the constant terms decreasing and the slopes increasing as feed protein content decreased. One possible explanation for this effect would be that increasing amounts of lipid are deposited in the breast muscle as feed protein content is reduced resulting in increasing breast weights at a given body protein weight. It is known that body lipid deposition increases as feed protein content declines (Gous et al., 1990) and this is evident also from the results of this trial (Table 3), where maximum rates of body growth were achieved on feeds intermediate in protein content. Such differences in lipid deposition would help to explain the two- and three-way interactions evident between factors, as the different strains and sexes deposited body lipid at different rates on the various dietary protein levels. If the skin and bone had been added to the portion of breast measured, the amount of lipid incorporated into the portion, and hence the effect of dietary protein content, would have been greater. But as the lipid content of the breast meat was not determined in this trial it is not possible to state to what extent differences in lipid deposition occurred in this muscle between treatments, and this information would clearly be useful in explaining the change in this allometric relationship with feed protein content.
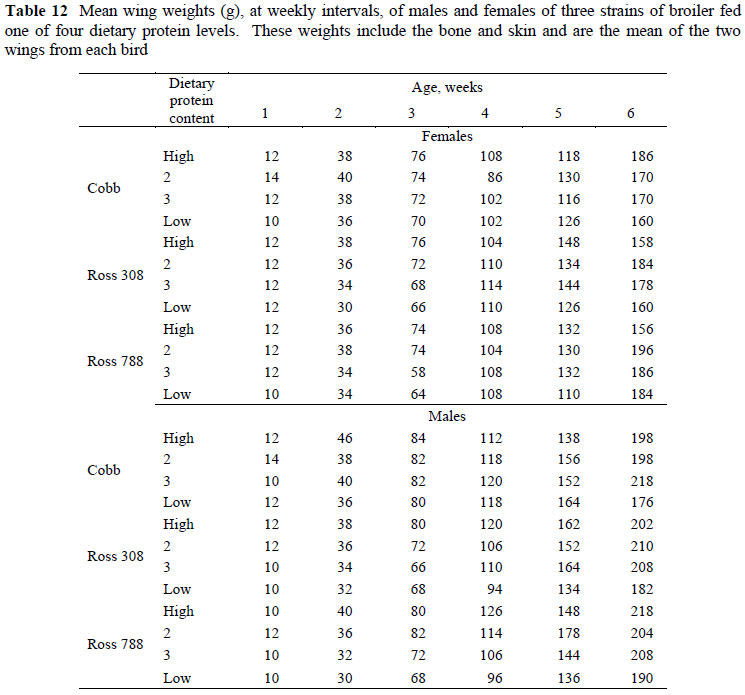
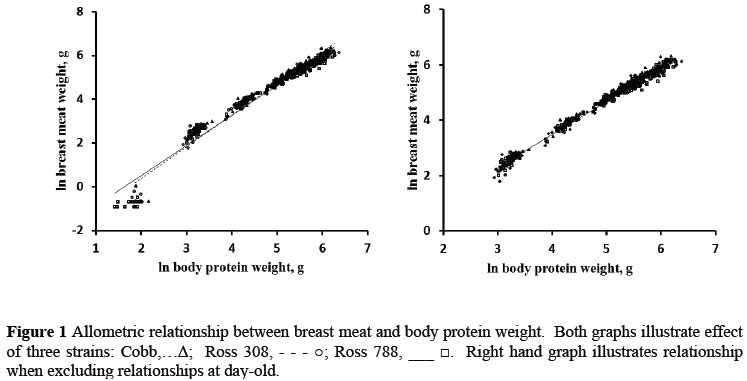
The relationships between ln breast meat and ln body protein weight and between ln thigh weight and ln body protein weight are illustrated in Figures 1 and 2 for the three strains and the four feed protein levels. It is evident in Figure 1 that the weights of breast meat at day old did not conform to the allometric relationship developed during the remaining six week growing period: all day old weights fell below the regression line, suggesting that nutrients had been depleted from this tissue prior to, or immediately after hatching. Only the breast meat was affected in this way as no other tissues measured in this trial fell below the fitted allometric regression at day old. There is evidence in the literature that breast tissue is used to supply nutrients to the body when these are needed and when they are not supplied by the feed (Halevy et al., 2006; Noy & Sklan, 1998). Although early and in ovo feeding have been shown to increase breast muscle weight (Uni & Ferket, 2004; Noy & Sklan, 1997; 1998), the weights reported on those held and early-fed treatments have not been accompanied by body protein weights, so it is not possible to determine whether such treatments increased the breast meat weight specifically at an equivalent body protein weight, or whether the weights of all components were similarly increased.
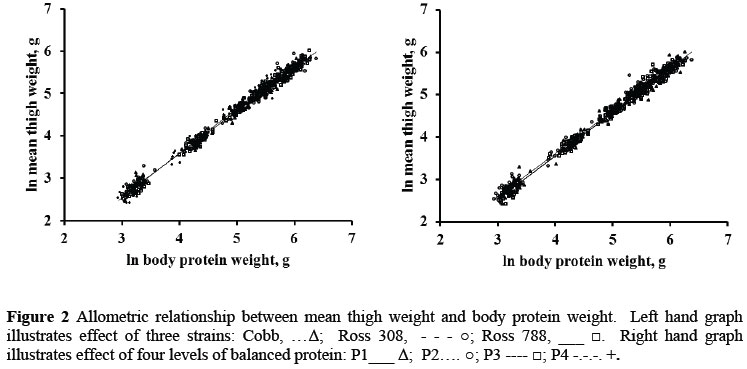
The relationships between the thigh and drum and body protein weight were marginally affected by strain and sex but more so by the feed protein content, again suggesting that these differences may be the result of increasing amounts of lipid being deposited in these components as feed protein content is reduced. Ideally, the weight of protein in the various parts should be measured, to be regressed against body protein weight, rather than including the lipid content of these parts in deriving allometric equations, for the same reason as was given for using body protein content as the independent variable rather than body weight. But it would then be necessary to determine the amount of lipid that is deposited in each of the body parts depending on the feed protein content, and this would have to be added to the weight of protein (and bone where appropriate) to predict the weight of each component. If the amount of lipid deposited in each part, on feeds differing in protein content, could be predicted, the weights of the parts could be adjusted by this amount to improve the accuracy of prediction, without having to define a different allometric equation for each level of dietary protein fed to broilers.
The allometric regressions presented here are the first such regressions to be published in the scientific literature. No previous relationships have regressed weights of different body components against body protein weight, thereby ignoring the variable amounts of body lipid that are deposited in broilers dependent on the genotype and feed protein content. The regressions presented here suffer also from the confounding effects of body lipid, which in this case are at the level of the body components and not the entire body. Although the equations presented here are more useful in predicting the weights of the physical parts of a broiler than any previous attempts, such predictions would be improved if the amount of lipid that is deposited in each component in relation to the feed protein content were known, or could be predicted. Apart from addressing a general problem of using body protein as a predictor of component weights whilst avoiding the problem of fatness, such information would be valuable to simulation modellers attempting to predict the weights of these important components of the body.
Conclusions
The research reported here suggests that, aside from the variable amounts of lipid that are deposited in the various body components resulting from differences in dietary protein content, the weights of these components may be accurately estimated from the body protein content of the broiler. It appears that day-old breast meat weight is influenced by the nutritional status of the embryo but that no other components measured in this trial are thus affected. To improve the accuracy of prediction of component weights as influenced by strain, sex and dietary protein content the minimum and maximum amounts of lipid that can be deposited in these components by different broiler strains should be measured.
References
AOAC, 2003. Official Methods of Analysis. (17th ed.). Association of Official Analytical Chemists, Inc., Arlington, Washington, USA. [ Links ]
Aviagen 2009. http://www.aviagen.com/ss/ross-708/ (accessed July 2010). [ Links ]
Cobb 2007. http://www.cobbvantress.com/Products/ProductProfile/Cobb700_Sales_Brochure_2007.pdf (accessed July 2010). [ Links ]
EFG Software 2007. http://www.efgsoftware.net/ (accessed July 2007). [ Links ]
Emmans, G.C., 1981. A model of the growth and feed intake of ad libitum fed animals, particularly poultry. In: Computers in Animal Production. Br. Soc. Anim. Prod. 5, 103-110. [ Links ]
Emmans, G.C., 1987. Growth, body composition and feed intake. Wrld Poult. Sci. J. 43, 208-227. [ Links ]
Emmans, G.C., 1988. Genetic components of potential and actual growth. pp. 153-181. In: Animal Breeding Opportunities. Occ. Pub. No 12. Br. Soc. Anim. Prod., Penicuik, Midlothian, U.K. [ Links ]
Fisher, C. & McNab, J.M., 1987. Techniques for determining the ME content of poultry feeds. In: Recent Advances in Animal Nutrition. Eds Haresign, W. & Cole, D.J.A., Butterworths, London. pp. 3-17. [ Links ]
GenStat, 2002. GenStat statistical software, Release 6.1 Lawes Agricultural Trust. [ Links ]
Gous, R.M., 2010. Scientific Research: The Planning Process. S. Afr. J. Anim. Sci. 40 (Suppl.1), 400-406. [ Links ]
Gous, R.M., Emmans, G.C., Broadbent, L.A. & Fisher, C., 1990. Nutritional effects on the growth and fatness of broilers. Br. Poult. Sci. 31, 495-505. [ Links ]
Gous, R.M., Pym, R.A.E., Mannion, P. & Wu, J.X., 1996. An evaluation of the parameters of the Gompertz growth equation that describe the growth of eight strains of broiler. Aust. Poult. Sci. Symp. 8, 174-177, University of Sydney, Sydney, NSW, Australia. [ Links ]
Gous, R.M., Moran Jr., E.T., Stilborn, H.R., Bradford, G.D. & Emmans, G.C., 1999. Evaluation of the parameters needed to describe the overall growth, the chemical growth and the growth of feathers and breast muscles of broilers. Poult. Sci. 78, 812-821. [ Links ]
Hancock, C.E., Bradford, G.D., Emmans, G.C. & Gous, R.M., 1995. The evaluation of the growth parameters of six strains of commercial broiler chickens. Br. Poult. Sci. 36, 247-264. [ Links ]
Hakansson, J., Eriksson, S. & Svensson, S.A., 1978. Influence of feed energy level on chemical composition of tissues and on the energy and protein utilization by broiler chicks. Swedish University of Agricultural Sciences, Department of Animal Husbandry, Report 59, Uppsala, Sweden. [ Links ]
Halevy, O., Yahav, S. & Rozenboim, I., 2006. Enhancement of meat production by environmental manipulations in embryo and young broilers. Wrld Poult. Sci. J. 62, 485-497. [ Links ]
Havenstein, G.B., Ferket, P.R. & Qureshi, M.A., 2003. Carcass composition and yield of 1957 versus 2001 broilers when fed representative 1957 and 2001 broiler diets. Poult. Sci. 82, 1509-1518. [ Links ]
Kemp, C., Fisher, C. & Kenny, M., 2005. Genotype - nutrition interactions in broilers; response to balanced protein in two commercial strains. Proc. 15th Eur. Symp. Poult. Nutr., Balatonfured, Hungary. pp. 54-56. [ Links ]
Lindsay, H.A., 1992. Allometric relationships between the physical parts of broiler chickens. MSc Agric. thesis, University of Natal, South Africa. [ Links ]
McKay, J.C., 2008. The genetics of modern commercial poultry. In: Proc. XXIII Wrld Poult. Congr., Brisbane, Australia. [ Links ]
Morris, T.R., 1983. The interpretation of response data from animal feeding trials. In: Recent Advances in Animal Nutrition. Ed. Haresign, W., Butterworths, London. pp. 12-23. [ Links ]
Noy, Y. & Sklan, D., 1997. Posthatch development in poultry. J. Appl. Poult. Res. 6, 344-354. [ Links ]
Noy, Y. & Sklan, D., 1998. Yolk utilisation in the newly hatched poultry. Br. Poult. Sci. 39, 446-451. [ Links ]
Schmidt Nielsen, K., 1984. Scaling, Why is animal size so important? Camb. Univ. Press, New York. [ Links ]
Uni, Z. & Ferket, R.P., 2004. Methods for early nutrition and their potential. Wrld Poult. Sci. J. 60, 101-111. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence.
See: http://creativecommons.org/licenses/by/2.5/za/
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science
* Corresponding author: gous@ukzn.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}