










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 n.2 Pretoria Jan. 2011
SHORT COMMUNICATION
The effect of dosing Megasphaera elsdenii NCIMB 41125 (Me) on lactation performance of multiparous Holstein cows
P.H. HenningI,#,*; L.J. ErasmusII; H.H. MeissnerI; C.H. HornI
IAgricultural Research Council, Private Bag x2, Irene 0062, South Africa
IIDepartment of Animal and Wildlife Sciences, University of Pretoria, 0002 Pretoria, South Africa
ABSTRACT
The objective of the study was to determine whether early post-partum dosing of Megasphaera elsdenii NCIMB 41125 (Me) will be beneficial to performance of high producing TMR-fed cows. Sixty multiparous Holstein cows were randomly allocated to four treatments (60% or 70% concentrate diet and placebo or Me [single oral dose of 1011 cfu in 250 mL suspension on day of calving and Days 10 and 20 post-partum, respectively]). Observations were recorded between calving and 80 days post-partum. Performance data were analysed for all 60 cows combined and for the 40 highest producing cows only, since they were considered more susceptible to ruminal acidosis. For all 60 cows, body weight, condition score and milk yield tended to increase with Me, but data for the 40 highest producing cows suggested that this response could be ascribed primarily to higher producing cows on the higher concentrate diet. Dry matter intake and milk protein were not affected by Me, whereas milk fat percentage increased with Me but only in cows on the 60% concentrate diet. Results support the hypothesis that dosing with Megasphaera elsdenii is most likely to benefit higher producing cows with greater risk of acidosis.
Keywords: Dairy cow, SARA, concentrate, feed intake, body weight, milk yield
Ruminal acidosis, especially sub-acute ruminal acidosis (SARA), is a growing concern in high producing dairy cows (Oetzel, 2003; Enemark, 2008) because of associated ill-health, poor production, lameness due to laminitis, and early culling (Hall & Averhoff, 2000; Oetzel, 2003). The condition normally occurs during the transition and early post-partum phases, when the cow is not yet adapted to increasing levels of non-structural carbohydrates (NSC), and often results in irregular feed intake (Beauchemin & Penner, 2009). Volatile fatty acids (VFA's) accumulate in the rumen because fermentation exceeds removal, and variability in ruminal pH is aggravated by fluctuating intake. If ruminal pH remains below 5.6 for more than 3h/24h (Plaizier et al., 2008), or below 5.8 for more than 5.2h/24h (Zebeli et al., 2008), the condition is defined as SARA, which is associated with reduced fibre digestion, rumen wall damage, endotoxin release into the bloodstream and inflammation (Gozho et al., 2006; Plaizier et al., 2008; Ametaj et al., 2009). The situation may be aggravated by production of much stronger lactic acid during rapid rumen fermentation of NSC. Maintaining ruminal pH above SARA levels through rapid removal of lactate and limiting accumulation of weaker acids, is central to preventing the condition or mitigating its effect.
Megasphaera elsdenii (Me) is the most important lactate-utilising bacterium in the rumen, but its numbers may often be low during the transition and early post-partum phases. Megasphaera elsdenii NCIMB 41125 (Me) was isolated in South Africa from cattle adapted to high-concentrate diets, and developed as a direct-fed microbial (DFM) for high-producing ruminants, including dairy cows. Administering of Me as an oral dose during transition from mainly roughage to high levels of concentrate diets has proved effective in removing lactic acid and maintaining ruminal pH above SARA thresholds (Henning et al., 2010a; b; Meissner et al., 2010). The objective of this research was to determine whether Me, when dosed to dairy cows during adaptation post-partum, will be beneficial to milk production and associated productivity measures.
Sixty multiparous Holstein cows with milk yields in excess of 9000 kg (305 days) in the previous lactation were blocked for milk production and body weight, resulting in 15 blocks of four cows each. Within block, cows were randomly allocated to one of the following treatments:
- 60% concentrate diet, cows not dosed with Me (Control) (C60);
- 60% concentrate diet, cows dosed with Me (Me60);
- 70% concentrate diet, cows not dosed with Me (Control) (C70);
- 70% concentrate diet, cows dosed with Me (Me70).
The diets were presented as total mixed rations (TMR).
Cows were fed ad libitum and milked twice per day. Me-treated cows were dosed with 1011 cfu of Me (in 250 mL suspension) on day of calving and again on days 10 and 20 post-partum. Control cows were similarly dosed with 250 mL distilled water. Trial observations were recorded between day of calving and Day 80 post-partum. Feed intake and milk yield were measured daily, milk fat and protein weekly, and body weight and condition score monthly. Feed samples were collected weekly, composited within treatment and stored at -20 ºC for analyses. The samples were analysed for dry matter (DM, AOAC, 2000, method 934.01), organic matter (OM) (Engels & Van der Merwe, 1967), nitrogen (N, AOAC, 2000, method 920.40), ether extract (EE, AOAC, 2000, method 920.39), nitrogen free extract (NDF) (Van Soest et al., 1991), ash (AOAC, 2000, method 942.05), soluble crude protein (CP, AOAC, 2000, method 988.05), Ca (Castellano Giron, 1973) and P (AOAC, 2000, method 965.17). Non-structural carbohydrates (NSC) were calculated from the equation: NSC% = 100 - (CP% + NDF% + EE% + Ash%) and metabolizable energy (ME), rumen degradable protein (RDP) and undegradable protein (UDP) calculated according to Van der Merwe & Smith (1991). Composite milk samples were analysed for fat and protein using the System 4000 Infrared Analyser (Foss Electric, Hillerod, Denmark).
Data were analysed as a completely randomized block design, using the GenStat® (2005) programme (Payne et al., 2005). Previous lactation milk production was used as covariate and milk yields are reported as covariate adjusted values. Since there was no significant dietary × dosing treatment interaction, contrast analysis was used to determine significant differences between treatments, respectively: Me vs. Control, Me60 vs. C60, and Me70 vs. C70. Differences were declared significant at P <0.05 and considered as trends at P <0.10.
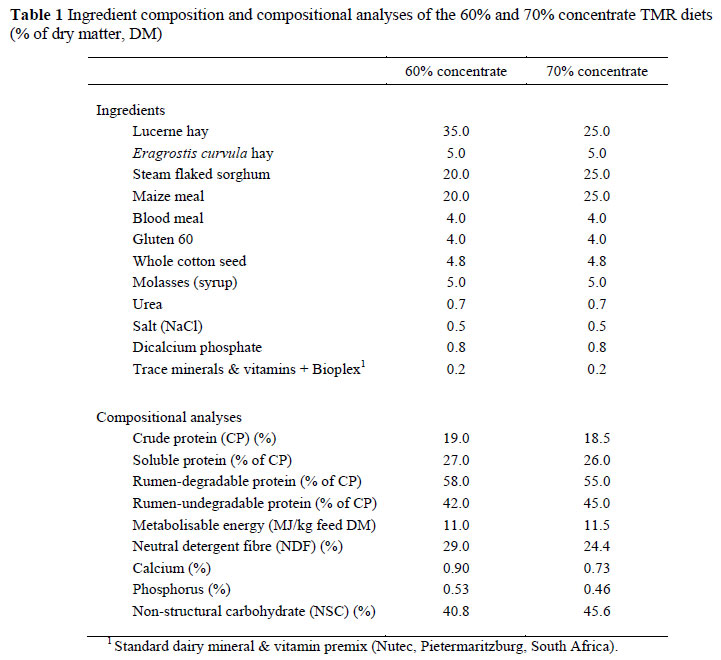
The composition of the experimental diets is shown in Table 1. Diets were formulated to fulfil the energy and nutrient requirements of a 680 kg cow, producing 40 kg milk per day, containing 3.6% fat (NRC, 2001).
From an energy supply point of view the primary differences between the two diets are the NDF and NSC levels. The latter reflects starch content. Cows on the 70% concentrate diet with more NSC, therefore, were expected to be more susceptible to SARA.
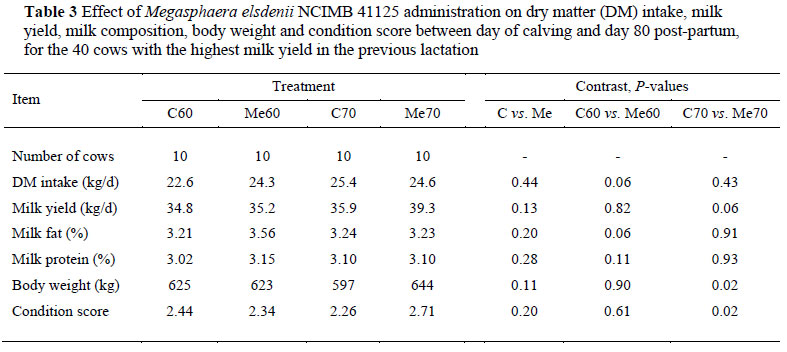
Table 2 shows the production data for all 60 cows whereas Table 3 shows only the data for the 40 cows from the 10 blocks which had the highest milk yields in the previous lactation. The rationale for the split in presentation of the data is (1) that higher producing cows, compared to lower producing cows, are expected to have higher feed intakes with more NSC's entering the rumen, thus rendering them more susceptible to SARA, and (2) if Me administration is successful, it is hypothesised that the effect should be more explicit in higher-producing cows with their greater energy (NSC) intakes and acidosis risk.
The 30 cows dosed with Me tended to produce more milk (average 35.1 kg vs. 33.1 kg) and had higher body weights (average 640 kg vs. 610 kg) and condition scores (average 2.63 vs. 2.38) than Control cows (Table 2). These differences were accentuated in the milk production, body weight and condition score responses of the higher-producing cows on the higher energy diet (Me70) as compared to the corresponding cows on the Control diet (C70) (Table 3). The corresponding differences for higher-producing cows on the lower energy diet (Me60 vs. C60) were not significant. This supports the hypothesis that cows on higher energy diets are more likely to benefit from Me administration. In fact, if the lower body weight and poorer condition score of cows on C70 compared to cows on Me70 are considered (Table 3), C70 cows probably suffered from SARA whereas the condition was mitigated in Me70 cows. This notion is supported by the results of Aikman et al. (2009).
Milk fat percentage was increased in cows dosed with Me on the 60% concentrate diet but not in cows on the 70% concentrate diet, whereas milk protein was unaffected. This is in contrast to the results of Aikman et al. (2009) who found that milk fat and protein were reduced by Me in high-producing cows on both their standard and high-energy diets. Hagg et al. (2010) on the other hand found no significant effect of Me on milk composition. These variable responses in milk composition are probably associated with differences in dietary composition and feed processing: the primary energy source in the present trial was steam-flaked sorghum and maize meal, whereas it was wheat and barley in the trial of Aikman et al. (2009) and maize meal only in the trial of Hagg et al. (2010). It is well-established that these sources differ in ruminal fermentation rate, and consequently in ruminal passage rate (NRC, 2001).
It is concluded that dosing cows with Megasphaera elsdenii NCIMB 41125 post-partum (when intake of NSC usually increases rapidly) can be beneficial, but that the production response may be greater in higher-producing animals, and may be affected by the source of dietary energy and feed processing.
References
Aikman, P.C., Henning, P.H., Horn, C.H. & Jones, A.K., 2009. Effect of using Megasphaera elsdenii NCIMB 41125 as a probiotic on feed intake and milk production in early lactation dairy cows. In: Ruminant Physiology - Digestion, metabolism and effects of nutrition on reproduction and welfare. Eds Chilliard, Y.. Glasser, F., Faulconnier, Y., Bocquier, F., Veissier, I. & Doreau, M., Wageningen Academic Publishers. pp. 110-111. [ Links ]
Ametaj, B.N., Koenig, K.M., Dunn, S.M., Yang, W.Z., Zebeli, Q. & Beauchemin, K.A., 2009. Backgrounding and finishing diets are associated with inflammatory responses in feedlot cattle. J. Anim. Sci. 87, 1314-1320. [ Links ]
A.O.A.C., 2000. Official methods of analysis (17th ed.). Association of Official Analytical Chemists, Inc., Arlington, V.A., USA. [ Links ]
Beauchemin, K.A. & Penner, G.B., 2009. New developments in understanding ruminal acidosis in dairy cows. Tri-State Dairy Nutrition Conf., 21-22 April 2009. pp. 1-12. [ Links ]
Castellano Giron, H., 1973. Comparison between dry ashing and wet digestion in the preparation of plant material for atomic absorption analysis. Atomic Absorption Newslet. 12, 28. [ Links ]
Enemark, J.M.D., 2008. The monitoring, prevention and treatment of sub-acute ruminal acidosis (SARA): A review. The Vet. J. 176, 32-43. [ Links ]
Engels, E.A.N. & Van der Merwe, F.J., 1967. Application of an in vitro technique to South African forages with special reference to the effect of certain factors on the results. S. Afr. J. Agric. Sci. 10, 983-992. [ Links ]
Gozho, G.N., Krause, D.O. & Plaizier, J.C., 2006. Rumen lipopolysaccharide and inflammation during grain adaptation and subacute ruminal acidosis in steers. J. Dairy Sci. 89, 4404-4413. [ Links ]
Hagg, F.M., Erasmus, L.J., Henning, P.H. & Coertze, R.J., 2010. The effect of a probiotic (Megasphaera elsdenii NCIMB 41125) on the productivity and health of Holstein cows. S. Afr. J. Anim. Sci. 40, 101-112. [ Links ]
Hall, M.B. & Averhoff, K.S., 2000. The real cost of digestive upset. Proc. 37th Florida Dairy Prod. Conf. Gainesville, 2-3 May, 2000. pp. 99-104. [ Links ]
Henning, P.H., Horn, C.H., Leeuw, K-J., Meissner, H.H. & Hagg, F.M., 2010a. Effect of ruminal administration of the lactate-utilizing strain Megasphaera elsdenii (Me) NCIMB 41125 on abrupt or gradual transition from forage to concentrate diets. Anim. Feed Sci. Technol. 157, 20-29. [ Links ]
Henning, P.H., Horn, C.H., Steyn, D.G., Meissner, H.H. & Hagg, F.M., 2010b. The potential of Megasphaera elsdenii isolates to control ruminal acidosis. Anim. Feed Sci. Technol. 157, 13-19. [ Links ]
Meissner, H.H., Henning, P.H., Horn, C.H., Leeuw, K-J, Hagg, F.M. & Fouché, G., 2010. Ruminal acidosis: A review with detailed reference to the controlling agent Megasphaera elsdenii NCIMB 41125. S. Afr. J. Anim. Sci. 40, 79-100. [ Links ]
NRC, 2001. National Research Council, 2001. Nutrient Requirements of Dairy Cattle, 7th Rev. Ed. National Academy Press, Washington D.C., USA. [ Links ]
Oetzel, G.R., 2003. Subacute ruminal acidosis in dairy cattle. Proc. Western Canadian Dairy Seminar: Advances in Dairy Technology 15, 307-317. [ Links ]
Payne, R.W., Murray, D.A., Harding, S.A., Baird, D.B. & Soutar, D.M., 2005. GenStat® for WindowsTM 10th Edition Introduction, VSN International, UK. [ Links ]
Plaizier, J.C., Krause, D.O., Gozho, G.N. & McBride, B.W., 2008. Subacute ruminal acidosis in dairy cows: The physiological causes, incidence and consequences. The Vet. J. 176, 21-31. [ Links ]
Van der Merwe, F.J. & Smith, W.A., 1991. Dierevoeding. Printomatic, Long Street, Cape Town, South Africa. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lewis, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and non-starch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583-3597. [ Links ]
Zebeli, Q, Dijkstra, J., Tafaj, M., Steingass, H., Ametaj, B.N. & Drochner, W., 2008. Modeling the adequacy of dietary fiber in dairy cows, based on the responses of ruminal pH and milk fat production to composition of the diet. J. Dairy Sci. 91, 2046-2066. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence. See: http://creativecommons.org/licenses/by/2.5/za/
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science
# Corresponding author: pieterh@msbiotech.biz
* Current address: Megastarter Biotech Pty Ltd t/a MS Biotech, PO Box 10520, Centurion 0046, South Africa


{kind=link}
{kind=link}
{kind=link}