










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.41 n.1 Pretoria Jan. 2011
SHORT COMMUNICATION
Expression profiling analyses of porcine MuRF1 gene and its association with muscle production traits
H. ShenI; M.X. LiaoI; S.H. ZhaoI; X.Y. LiI; B. FanI,II,#
IKey Laboratory of Agricultural Animal Genetics, Breeding & Reproduction of Ministry of Education, Key Laboratory of Swine Genetics & Breeding of Ministry of Agriculture, Huazhong Agricultural University, Wuhan 430070, P R China
IIDepartment of Animal Science and Centre for Integrated Animal Genomics, Iowa State University, Ames, 50011, USA
ABSTRACT
Muscle specific RING finger protein-1 (MuRF1) is a member of the muscle specific RING finger protein family, and it is specifically expressed in cardiac and skeletal muscle tissues and is involved in myocyte differentiation, development and morphogenesis. In this study the complete open reading frame (ORF) of the porcine MuRF1 gene consisting of 354 amino acids was obtained and it shared 93% and 90% identity with those of the human and mice, respectively. Using the INRA radiation hybrid panel (IMpRH) technique, the MuRF1 gene was assigned to SSC6q21-26, closely linked to microsatellite markers SW1823 and SW709. The tissue distribution patterns revealed that MuRF1 mRNA was exclusively expressed in cardiac and skeletal muscle tissues. Real-time quantitative reverse transcriptase-polymerase chain reaction (RT-qPCR) results displayed that MuRF1 mRNA was up-regulated in Landrace pigs during the prenatal skeletal muscle development stages. A synonymous T/C single nucleotide polymorphism (SNP) was identified in MuRF1 exon 3 and then a Hin6I PCR -RFLP was developed for SNP genotyping in two pig populations. Association of the genotypes with growth and carcass traits showed that different genotypes of MuRF1 were genetically significantly associated with average daily gain from birth to 90 kg and loin muscle area in one experimental population. The study suggested that the porcine MuRF1 gene might affect muscle growth and development, and could be a potential candidate gene for muscle production traits in the pig.
Keywords: Molecular cloning, expression profile, mapping, association analysis
The muscle specific RING finger protein (MuRF) family comprises mainly of three members, MuRF1, MuRF2 and MuRF3, all of which are specifically expressed in cardiac and skeletal muscle tissues. These MuRF proteins share one highly conserved N-terminal RING domain, which is followed by a zinc-binding B-box motif and two coiled-coil dimerization motif boxes (Centner et al., 2001). MuRF1 (Muscle-specific RING finger-1) was initially described as striated muscle RING zinc finger (SMRZ) (Dai & Liew, 2001). The yeast two-hybrid interaction experiments and glutathione S-transferase (GST) -pulldown assays indicated that MuRF1 binds to the titin repeats, A168/A169, which is adjacent to the titin kinase domain. Centner et al. (2001) suggested that the interaction between titin and MuRF1 was critical for the stability of the sarcomeric M-line region, as well as the thick filament (McElhinny et al., 2002). In addition, MuRF1 has been identified as one of the members of the E3 ubiquitin ligases and it regulates myofibrillar protein metabolism (Bodine et al., 2001). In vitro treatment of myotubes with dexamethasone induced atrophy and showed a specifically increased expression of MAFbx and MuRF1, and mice deficient in either MAFbx or MuRF1 were resistant to atrophy (Sandri et al., 2004; Stitt et al., 2004). Until now the molecular characteristics and biological function of porcine MuRF1 gene have been less reported. This study is the first report showing the molecular characteristics and associations of the MuRF1 gene in the pig.
The whole cDNA sequence of the human MuRF1 gene retrieved from the NCBI database (GenBank accession No: NM_032588) was used for BLAST to search the homologous porcine ESTs (http://www.ncbi.nlm.nih.gov/blast). The consensus sequences of porcine ESTs were obtained using the Align programme of DNASTAR software (DNAStar, Madison, WI, USA). The primer design for the porcine MuRF1 gene cDNA amplification (5'-AGGCTTCGGGTGGGTATT-3'/ 5'-TCCTGTGACTGGTGTGCTT-3') was implemented using Primer 5.0 software (Premier Biosoft International, Palo Alto, CA, USA). The PCR condition was performed in a 20 IL volume, containing 1 × PCR buffer, 50 ng DNA, 0.3 IM of each primer, 80 IM of each dNTP and 1.0 U Taq DNA polymerase (TaKaRa, Dalian, China). The PCR programme was as follows: 94 ºC for 4 min; 35 cycles at 94 ºC for 30 s, 59 ºC for 30 s, and 72 ºC for 45 s; and a final extension for 10 min at 72 ºC. The PCR products were gel-purified and then cloned into the pMD18-T vector (TaKaRa, Dalian, China), and two isolated clones were sequenced commercially by Invitrogen (Invitrogen, Shanghai, China) to confirm the identity. The procedure was in accordance with the protocol described by Cheng et al. (2010). The open reading frame (ORF) of the obtained porcine MuRF1 cDNA sequence was predicted using the online ORFfinder programme (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). Characterization of the amino acid sequence and structure prediction were conducted using ExPaSy tools (http://cn.expasy.org). The alignment of amino acid sequences among different species was performed, using the online CLUSTALW2 programme (http://www.ebi.ac.uk/Tools/clustalw2).
Total RNA was isolated from nine different tissues of an adult Large White female pig, namely heart, liver, spleen, lung, kidney, longissimus dorsi muscle, subcutaneous fat, stomach and duodenum with Trizol reagent (Gibco-BRL, Rockville, USA) following the supplier's protocol. Gene expression patterns were determined by the RT-PCR technique. The PCR mixture consisting of 2 µg total RNA and 5 µL oligo (dT) was incubated at 70 ºC for 5 min. The reactions were chilled on ice for 2 min and the remaining reagents including 5 µL dNTPs (10 mM each dNTP), 10 µL 5 × buffer, 2.5 µL RNAase inhibitor with 300 U M-MLV reverse transcriptase (Promega, Madison, WI, USA) were added to a total volume of 50 µL. Reverse transcription reaction was proceeded for 1 h at 42 ºC. Finally, the reverse transcriptase was inactivated by 5 min incubation at 90 ºC. 2 µL single-stranded cDNA was amplified with MuRF1 express primers (5'-GACTTGATGTGCTGTACGC-3'/5'-CTCAGGGTGTCTGCTATGT-3'). Five µL PCR products were used to detect the expression profile in agarose gel, and the 18S cDNA was performed as a positive control.
To evaluate the expression level of porcine MuRF1 during different muscle development stages (33, e 65 and 90 dpcs), SYBR® Green I-based real-time quantitative PCR RT-qPCR )was employed and 18S ribosome RNA (rRNA) was used as a reference gene. The reverse transcription of 2 Ig total RNA (three Landrace pigs for each stage) was performed as described above. Each RT-qPCR system (20 IL) contained 1 × SYBR Green master buffer (Toyobo, Osaka, Japan), 0.6 IM of each express primer and 1 IL template cDNA. The PCR reaction was completed on the Bio-Rad iQTM5 real-time PCR detection system (Bio-Rad, Hercules, CA, USA) according to the following programme: 95 ºC for 4 min; followed by 40 cycles of 94 ºC for 30 sec, 62 ºC for 40 sec and 72 ºC for 20 sec; then 15 ºC. The gene expression level of porcine MuRF1 gene was quantified relative to the expression of 18S by employing the comparative Ct (or LCt) value method (Livak & Schmittgen, 2001). The ΔCt values were calculated and student's t-test was conducted on these averages to identify the expression difference.
The INRA-University of Minnesota porcine radiation hybrid panel (IMpRH) was employed for chromosomal mapping of porcine MuRF1 gene. The pig-specific primers for MuRF1 gene mapping were designed based on exon 1 sequence (5'-GTGACAAAGGCAAGACCC-3'/5'-ACACGGCAAGATGACCACC-3'). The genotyping results were input into the online IMpRH mapping tool (http://imprh.toulouse.inra.fr) for RH mapping (Milan et al., 2000).
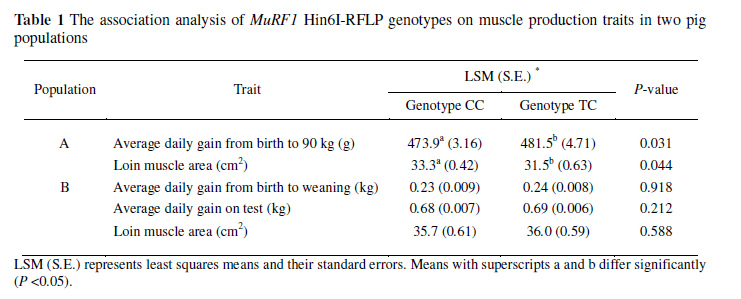
The identification of SNP was accomplished by sequencing the pooled PCR products amplified from six DNA samples and each two were from the Large White, Landrace and Tongcheng pig breeds, respectively. The polymorphism site was detected using the DNASTAR software (DNAStar, Madison, WI, USA) and was validated by the PCR-restriction fragment length polymorphism (PCR-RFLP) method. SNP genotyping and association analysis were implemented in two different pig populations. Population A included 159 pigs that were from the Tongcheng pig (n = 40), the Landrace (n = 25), Large White (n = 22), Large White × (Landrace × Tongcheng) (n = 38) and Landrace × (Large White × Tongcheng) (n = 34). A total of 19 traits consisting of growth, body composition and meat quality were recorded (Tang et al., 2008). The association analysis between genotypes and traits were examined using the mixed procedure of SAS (SAS Institute, release 9.1, Cary, N.C., USA). The model included breed/breed combination, sex, slaughter date and marker genotyping as fixed effects, dam as a random effect and body weight as a covariate. Population B was the ISU Berkshire × Yorkshire pig resource family comprising of 515 F2 animals, and the analyzed traits included birth weight, 16-day weight, average daily gain from birth to weaning, average daily gain on test, carcass weight, carcass length, back fat thickness and loin eye area (Malek et al., 2001). Association analysis was implemented using the mixed model procedure (SAS Institute, release 9.1, Cary, N.C., USA), including sex, slaughter date and marker genotypes as fixed effects, dam (litter) as random effect and body weight as covariate.
A 1168 bp full-length porcine MuRF1 cDNA was identified and contained a 1065 bp predicted ORF (GenBank accession No: FJ905227), which encoded 354 amino acids with a calculated molecular weight of 40.42 kDa and an estimated isoelectric point (pI) of 4.87. This protein contains one N-terminal RING domain, one Zn-binding B-box motif, one MuRF family-specific conserved box (MFC) and two coiled-coil dimerization motif boxes. The result confirmed that the porcine MuRF1 protein is a member of RBCC protein family, which is in agreement with a report on human (Centner et al., 2001). The coding sequence (CDS) of the porcine MuRF1 gene was 91% identical to that of the human (NM_032588) and the amino acids sequences of porcine MuRF1 protein shared 93% and 90% identity with those of humans (GenBank accession No: NP_115977.2) and mice (GenBank accession No: NP_001034137), respectively, which suggested that the MuRF1 gene might be highly evolutionarily conserved in mammal species.
The tissue expression patterns showed that MuRF1 mRNA was specially expressed in longissimus dorsi muscle and heart tissues, and was not detected in other tissues such as the liver, spleen, lung, kidney, subcutaneous fat, stomach and duodenum (Figure 1). RT-qPCR analysis results demonstrated that MuRF1 mRNA expression was up-regulated in Landrace pigs during prenatal skeletal muscle development stages (Figure 2). The porcine MuRF1 mRNA was found to be strictly expressed in striated muscle tissues, similar to humans (Dai & Liew, 2001). The studies showed that Akt1/Foxo/MAFbx/MuRF1 signalling pathway, plays an important role during the progression of skeletal muscle atrophy (Bodine et al., 2001; Nader 2005). Recent studies using MuRF1 KO mice demonstrated that MuRF1 acted as a modulator for energy homeostasis via ubiquitination in skeletal muscle and a supplier of branched chain amino acids (BCAAs) to other tissues by regulating muscle protein turnover (Koyama et al., 2008). The strict expression of MuRF1 in striated muscle tissues implies that it has an important function during porcine striated muscle development. Furthermore, the mRNA expression patterns of this gene showed that the MuRF1 mRNA expression was differentially up-regulated during three development stages of foetal skeletal muscle. Skeletal muscle development during the prenatal stages has important influences on muscle growth and meat quality (Wigmore & Stickland, 1983). Thus, it suggested that the MuRF1 gene might be involved in muscle development and might have a vital role during the stage between 60 and 90 dpc.
A T/C SNP (dbSNP accession No: ss185232798) was identified at position 19 of exon 3 of the MuRF1 gene and it caused a synonymous mutation of glycine. This SNP could not induce any change of restriction enzyme sites, therefore a pair of special primers was designed by creating a restriction site method (Haliassos et al., 1989), and then a Hin6I restriction enzyme site was produced. The PCR-RFLP pattern includes a 142 bp fragment of allele T and a 123bp and19 bp of allele C. Among the 159 pigs in population A, the number of animals carrying genotypes CC, TC and TT were 106, 51 and 2, respectively. Animals with genotype TT were excluded for the association analysis because of the low number of observations. The association analysis showed that the MuRF1 gene was significantly (P <0.05) associated with average daily gain from birth to 90 kg and loin muscle area (Table 1).Interestingly, animals possessing the homozygous, CC, had lower average daily gains from birth to 90 kg than animals with genotype, TC (P = 0.031), but they had more (P = 0.044) loin muscle area than those of pigs with the TC genotype. In population B, the number of individuals carrying CC, TC and TT were 203, 301 and 32, respectively. Similarly, animals with genotype TT were excluded for analysis. Although CC animals had a lower average daily gain from birth to weaning and average daily gain on test than those of TC animals, the difference was not significant (Table 1). It implies the discovery of additional SNPs, and association analysis in other populations of larger sizes and/or populations with genetic backgrounds are needed.
In humans, the homologous MuRF1 gene is located at 1p33-34 (Dai & Liew, 2001). The porcine MuRF1 gene was assigned to SSC6q21-26, being linked to two microsatellite markers SW1823 (LOD = 8.66, 0.42 cR) and SW709 (LOD = 5.39, 0.66 cR), which is in agreement with the comparative mapping results, since human chromosome 1 shares syntenic groups with porcine chromosomes 6. Several QTL related to lean percentage, intramuscular fat and average backfat thickness have been located around this region (http://www.animalgenome.org/cgi-bin/QTLdb). The association analysis indicated that different genotypes of the porcine MuRF1 gene have significant associations with average daily gain from birth to 90 kg and loin muscle area in one population, which were two important traits related to growth rate and meat production. The homozygote favourable to higher loin muscle area was associated with lower growth rate, which is supported by the findings that average daily gain had a low and negative genetic and phenotypic correlation with loin muscle area (Suzuki et al., 2005).
These findings suggested that MuRF1 might be involved in the regulation of muscle growth, and the SNP could be used as a genetic marker in the breeding and selection of market pigs. However, the function analysis, additional SNP discovery and verification of association are needed before this gene can be eventually utilized for marker assisted selection in pigs.
Acknowledgements
This study was financially supported by the National Nat ural Science Foundation of China (30771169),Wuhan Youth Chenguang Project (200750731282), the National High Science and Technology Foundation of China "863" (2010AA10A104), and the creative team project of the Chinese Ministry of Education IRT0831). The authors thank the members of the Max F. Rothschild's research group at Iowa State University for their assistance in this research.
References
Bodine, S.C., Latres, E., Baumhueter, S., Lai, V.K., Nunez, L., Clarke, B.A., Poueymirou, W.T., Panaro, F.J., Na, E., Dharmarajan, K., Pan, Z.Q., Valenzuela, D.M., DeChiara, T.M., Stitt, T.N., Yancopoulos, G.D. & Glass, D.J., 2001. Identification of ubiquitin ligases required for skeletal muscle atrophy. Science 294, 1704-1708. [ Links ]
Centner, T., Yano, J., Kimura, E., McElhinny, A.S., Pelin, K., Witt, C.C., Bang, M.L., Trombitas, K., Granzier, H., Gregorio, C.C., Sorimachi, H. & Labeit, S., 2001. Identification of muscle specific ring fingerproteins as potential regulators of the titin kinase domain. J. Mol. Biol. 306, 717-726. [ Links ]
Cheng, H.J., Xu, X.W., Zhao, S.H., Liu, B., Yu, M. & Fan, B., 2010. Molecular cloning and expression profile analysis of porcine TCAP gene. Mol. Biol. Rep. 37, 1641-1647. [ Links ]
Dai, K.S. & Liew, C.C., 2001. A novel human striated muscle RING zinc finger protein, SMRZ, interacts with SMT3b via its RING domain. J. Biol. Chem. 276, 23992-23999. [ Links ]
Haliassos, A., Chomel, J.C., Tesson, L., Baudis, M., Kruh, J., Kaplan, J.C. & Kitzis, A., 1989. Modification of enzymatically amplified DNA for the detection of point mutation. Nucleic Acids Res. 17, 3606. [ Links ]
Koyama, S., Hata, S., Witt, C.C., Ono, Y., Lerche, S., Ojima, K., Chiba, T., Doi, N., Kitamura, F., Tanaka, K., Abe. K., Witt, S.H., Rybin, V., Gasch, A., Franz, T., Labeit, S. & Sorimachi, H., 2008. Muscle RING-finger protein-1 (MuRF1) as a connector of muscle energy metabolism and protein synthesis. J. Mol. Biol. 376, 1224-1236. [ Links ]
Livak, K.J. & Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25, 402-408. [ Links ]
Malek, M., Dekkers, J.C., Lee, H.K., Baas, T.J., Prusa, K., Huff-Lonergan, E. & Rothschild, M.F., 2001. A molecular genome scab analysis to identify chromosomal regions influencing economic traits in the pig. II. Meat and muscle composition. Mamm. Genome 12, 637-645. [ Links ]
McElhinny, A.S., Kakinuma, K., Sorimachi, H., Labeit, S. & Gregorio, C.C., 2002. Muscle-specific RING finger-1 interacts with titin to regulate sarcomeric M-line and thick filament structure and may have nuclear functions via its interaction with glucocorticoid modulatory element binding protein-1. J. Cell Biol. 157, 125-136. [ Links ]
Milan, D., Hawken, R., Cabau, C., Leroux, S., Genet, C., Lahbib, Y., Tosser, G., Robic, A., Hatey, F., Alexander, L., Beattie, C., Schook, L., Yerle, M. & Gellin, J., 2000. IMpRH server: an RH mapping server available on the Web. Bioinformatics 16, 558-559. [ Links ]
Nader, G.A., 2005. Molecular determinants of skeletal muscle mass: getting the "AKT" together. The Intern. J. Biochem. Cell Biol. 37, 1985-1996. [ Links ]
Sandri, M., Sandri, C., Gilbert, A., Skurk, C., Calabria, E., Picard, A., Walsh, K., Schiaffino, S., Lecker, S.H. & Goldberg, A.L., 2004. Foxo transcription factors induce the atrophy-related ubiquitin ligase atrogin-1 and cause skeletal muscle atrophy. Cell 117, 399-412. [ Links ]
Stitt, T.N., Drujan, D., Clarke, B.A., Panaro, F., Timofeyva, Y., Kline, W.O., Gonzalez, M., Yancopoulos, G.D. & Glass, D.J., 2004. The IGF-1/PI3K/Akt pathway prevents expression of muscle atrophy-induced ubiquitin ligases by inhibiting FOXO transcription factors. Mol. Cell. 14, 395-403. [ Links ]
Suzuki, K., Kadowaki, H., Shibata, T., Uchida, H. & Nishida, A., 2005. Selection for daily gain, loin-eye area, backfat thickness and intramuscular fat based on desired gains over seven generations of Duroc pigs. Livest. Prod. Sci. 97, 193-202. [ Links ]
Tang, Z.L., Peng, Z.Z., Liu, B., Fan, B., Zhao, S.H., Li, X.P., Xu, S.P. & Li, K., 2008. Effect of breed, sex and birth parity on growth, carcass and meat quality in pigs. Front Agric. China 2, 331-337. [ Links ]
Wigmore, P.M. & Stickland, N.C., 1983. Muscle development in large and small pig fetuses. J. Anat. 137, 235-245. [ Links ]
Copyright resides with the authors in terms of the Creative Commons Attribution 2.5 South African Licence.
See: http://creativecommons.org/licenses/by/2.5/za/
Condition of use: The user may copy, distribute, transmit and adapt the work, but must recognise the authors and the South African Journal of Animal Science
# Corresponding author: bfan@iastate.edu


{kind=link}
{kind=link}
{kind=link}