










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.40 no.4 Pretoria Jan. 2010
Response of broiler chickens to diets containing artificially dried high-moisture maize supplemented with microbial enzymes
M.M. Bhuiyan; A.F. Islam; P.A. Iji#
Department of Animal Science, School of Environmental and Rural Science, University of New England, Armidale, NSW 2351, Australia
ABSTRACT
The effect of feeding high-moisture maize grains dried in the sun or artificially in a forced draught oven at 80, 90 or 100 ºC for 24 hours and supplemented with microbial enzymes (Avizyme 1502 and Phyzyme XP) on growth performance, visceral organs, tissue protein, enzyme activity and gut development was investigated in a broiler growth trial. Feed intake (FI) up to 21 days decreased as a results of oven drying of grains whereas supplementation with microbial enzymes increased FI compared to the non-enzyme diets (881.1 vs. 817.2 g/bird). The highest FI (900 g/bird) was found only in sun-dried grain diets. There was no effect of grain drying temperature or enzyme supplementation on FI when assessed at 7 days of age. Up to day 21 there was a reduction in live weight (LW) with increase in grain drying temperature while supplementation with enzymes significantly improved LW only on the diets containing sun-dried grains (731 g/bird) and grains dried at 90 ºC (634 g/bird). Live weight was significantly higher in chickens on the enzyme supplemented diets than on diets without enzymes (638 vs. 547 g/bird). The feed conversion ratio (FCR) at this age was poorer with an increase in grain drying temperature but improved when the diets were supplemented with enzymes (1.48 vs. 1.62 g/g). There was an increase in the relative weight of the small intestine and liver with an increase in grain drying temperature at day 21 but there was no difference in the relative weight of these organs when the diets were supplemented with enzymes. Only the activities of the alkaline phosphatase at day 7 and maltase and sucrase at days 7 and 21 increased as a result of grain drying treatment but not by microbial enzyme supplementation. The ileal digestibility of gross energy, protein and starch was not significantly changed with an increase in grain drying temperature or by enzyme supplementation. The concentrations of ileal formic and acetic acids and caecal propionic and valeric acids were significantly increased by an increase in grain drying temperature but not affected by the microbial enzyme supplementation. The populations of lactic acid and lactobacilli bacteria in the ileal content were reduced on diets containing enzymes but were not affected by an increase in grain drying temperature. In the caecal content, the total anaerobic bacterial count was higher in birds on diets supplemented with microbial enzymes (8.1 vs. 7.8 log10cfu x/g digesta). The resident lactic acid bacteria population also increased as a result of an increase in grain drying temperature. From results of the current study, diets based on sun-dried maize or maize dried at 90 ºC provided comparatively better gross response. It may be inferred that there was a positive response to addition of microbial enzymes. Overall, it may be assumed that, for broiler chickens, there is little or no difference in the nutritive value of sun-dried grain and grains artificially dried at 90 ºC.
Keywords: High moisture maize, drying temperature, bird performance, enzyme supplement, gut microflora
Introduction
In many parts of the world, especially in humid areas, maize is harvested at a relatively high moisture content, with a view to minimizing damage in the field when left to dry naturally. The grain is then subjected to artificial drying, which may result in loss of quality such as an increase in retrograde starch (Brown, 1996). The retrograde starch is caused by high temperature heating of grains followed by storage at a lower temperature. The digestibility of cereal grains is influenced by the starch component, especially the ratio between amylose and amylopectin (McDonald et al., 1995). Noy & Sklan (1994) stated that about 15% of maize starch is known to remain undigested up to the terminal ileum and is assumed to be resistant to digestion. This presents an opportunity for the use of exogenous enzymes as is done with wheat and other temperate cereals. Starch in high moisture grain also anneals due to heat processing. It is not well known how much of these processes occur during routine production and processing of maize.
Due to the amorphous nature of its structure, amylopectin is more readily digested than the amylose. Moreover, this readily available amylopectin content may increase due to artificial drying of high moisture grain and conversely amylose content may decrease. This was confirmed by Bhuiyan et al. (2010), but it is not known if further improvements could be achieved through supplementation of microbial enzymes.
The normal structure (spherical, 10 - 16 microns across) of starch granules with protein bodies and matrix may be altered easily and can create a favourable environment for enzymatic digestion (Taylor & Belton, 2002). However, the normal structure of starch granules may also change, becoming more shrunken if dried at a high temperature, and maize quality can be compromised (Bhuiyan et al., 2010). The research of Panigrahi et al. (1996), who evaluated the effects of artificial drying of grains that had stack-burn and reduction in in vitro digestibility, showed that the quality of diets with stack burned/overheated maize grain tends to negatively affect broiler performance. There are many reasons advanced for these reductions in quality such as denaturation of heat-labile vitamins and damage to proteins via an interaction with reducing sugars (Maillard reaction) and retrograde starch formation after heat processing (Panigrahi et al., 1996; Iji et al., 2003; Cowieson, 2005).
Exogenous enzymes have been used for many years to break down cell walls in feed ingredients, to reduce the viscosity of digesta and to improve the digestibility of carbohydrates and proteins (Bedford & Moran, 1996). Generally they are used to improve the nutritive value of wheat- and barley-based diets, but some enzyme preparations are also used to increase the nutritional value of maize and its by-products (Boros et al., 2004; Cowieson, 2005). However, in some investigations the positive effect of enzymes was not registered (Peric et al., 2002; Iji et al., 2003). There is little in the literature about the addition of enzyme mixture to diets based on artificially dried maize, especially if the grains were harvested with high-moisture.
The aim of this trial was to evaluate the response of high-moisture maize grain to artificial drying and supplementation with microbial enzymes. These investigations were carried out considering feed intake (FI), body weight (BW) and feed conversion ratio (FCR), as well as the physiological mechanisms behind these responses.
Materials and Methods
A 2 x 4 factorial experiment was performed to study the effect of drying high-moisture maize at different temperatures (sun-dried and artificially dried at 80, 90 or 100 ºC) and the addition of microbial enzymes to the diet on the nutritive value of the grain and physiological responses in broiler chickens. Three hundred and eighty-four (384) day-old male Cobb broiler chicks (Baiada Poultry Pty. Ltd, Tamworth, NSW, Australia), weighing 40.1 ± 0.08 g, were randomly assigned to 48 cages (600 × 420 × 23 cm each) in four-tier battery brooders, housed in an environmentally controlled house. Each of the eight treatments was randomly assigned to six cages with eight birds per cage. The birds were initially brooded at a temperature of 33 ºC, but this was gradually reduced to 24 ± 1 ºC at 21 days of age when the feeding trial ended. Sixteen hours of lighting per day were provided throughout the trial period.
Maize grain obtained from Inverell in northern New South Wales, Australia (2009 planting year; at the end of April) was used in this experiment. The maize contained a moisture content of 23% and was split into four batches. The first batch was dried in the sun for three days until the moisture content dropped to about 13%. The other three batches were dried artificially for 24 hours using a forced draught oven at 80, 90 or 100 ºC. After drying, the dry matter (DM) content was 95.0, 96.3 and 98.0%, respectively.
Four basal diets were formulated to meet minimum National Research Council (1994) nutrient recommendations for broiler chickens (Table 1). The experimental diets were semi-purified, comprising of more than 70% maize grain: sun-dried, or dried at 80, 90 or 100 ºC. Each of these diets was fed as such or supplemented with enzymes at a rate of 0.5 g/kg of Avizyme 1502 (containing amylase 800, xylanase 1200, protease 8000 U/g) and 0.1 g/kg of Phyzyme XP (1000 FTU). Both these enzymes were supplied by Danisco Animal Nutrition, UK. An indigestible marker, titanium dioxide (TiO2), was incorporated in the diets to assess nutrient digestibility of the feed. The diets were formulated to be iso-energetic and iso-nitrogenous, and fed for 21 days. Feeds and water were available to the birds ad libitum.
On days 7 and 21, two birds and three birds, respectively from each cage, randomly selected, were weighed and humanely killed by cervical dislocation. The abdominal cavity was opened and the small intestine was ligated and removed. The content of the ileum and caecum was collected in labelled plastic containers for the determination of nutrient digestibility and short-chain fatty acids (SCFA) content. Approximately 1 g of ileal and caecal samples was collected separately into pre-prepared McCartney bottles containing anaerobic broth for the enumeration of microbial populations. The McCartney bottles and plastic containers containing digesta samples were kept at -20 ºC pending bacterial enumeration and analysis of SCFA. For the determination of the TiO2 as well as nutrient digestibility, the digesta from the ileum were pooled on a cage basis, homogenized and stored at -20 ºC. Later, the samples were freeze-dried, ground through a small coffee grinding and stored in airtight containers at -4 ºC for the chemical analysis of TiO2, gross energy, starch and protein.
The experiment was approved by the Animal Ethics Committee of the University of New England (Approval No.: AEC09/004). Health and animal husbandry practices complied with the "Code of Practice for the Use of Animals for Scientific Purposes" issued by the Australian Bureau of Animal Health (National Health and Medical Research Council, 1990).
Live weights, live weight gain (LWG) and FI were recorded on a cage basis at weekly intervals. Mortality was recorded and FCR was corrected for mortality. The body weight and the weight of the proventriculus, gizzard and small intestine with content, pancreas, bursa of fabricius, yolk sac, spleen and liver were recorded at days 7 and 21. The relative organ weight was subsequently calculated as an indication of mass per unit of body weight (g/100 g of body weight).
To examine the digestive enzyme activity and protein concentration, the jejunal tissue was processed as described by Shirazi-Beechey et al. (1991). Sub-samples of the homogenate were taken, put into Eppendorf tubes (Eppendorf South Pacific, North Ryde, Australia) and stored in a freezer (-20 ºC) pending enzyme analysis. The pancreas was processed in a similar procedure to the jejunum except that Milli-Q water (Millipore Australia, North Ryde, Australia) was used instead of a buffer, and the entire tissue was homogenized (Nitsan et al., 1974). The specific activities of jejunal and pancreatic enzymes were assessed by incubation with fixed substrate concentrations as standardized for poultry by Iji et al. (2001b). On the jejunal homogenates, the assays were conducted for mucosal protein content and activities of alkaline phosphatase (EC 3.1.3.1), maltase (EC 3.2.1.20) and sucrase (EC 3.2.1.10). For the pancreas, assays were conducted for protein content and chymotrypsin amidase (EC 3.4.21.1). Specific activities of enzymes were measured according to the methods previously described for other species (Holdsworth, 1970; Serviere-Zaragoza et al., 1997) after standardization for poultry. The concentration of protein in both the jejunal mucosa and pancreatic tissue was measured using the Coomassie dye-binding procedure described by Bradford (1976). The raw data outputs were processed using the recommended Lowry Software (Mcpherson, 1985) before running the statistical analysis.
The analytical method described by Jensen et al. (1995) was adopted with modifications for the analysis of organic acid (SCFA, lactic acid and succinic acid) concentrations. Before running on a Varian CP3400 CX gas chromatograph (Varian Analytical Instruments, Palo Alto, CA, USA), sample vials were kept in a heating block at 80 ºC for 20 min and left at room temperature for 48 h. Total organic acid concentration was derived as the sum of all the organic acids observed in a sample, expressed as mg/g digesta after log10 +1 transformation.
The TiO2 content of the digesta and diet samples was measured according to the method of Short et al. (1996). Aliquots of the solutions obtained and of similarly prepared standard solutions were analyzed using a Hitachi 150-20 UV spectrophotometer (Hitachi Science Systems Ltd., Ibaraki, Japan), measuring the absorbance at 410 nm. The TiO2 content, measured in mg/mL, was determined from the standard curve and converted to mg/g of the sample. The TiO2 marker was used to calculate the digestibility coefficient. Diets and ileal digesta were analyzed for protein and gross energy using standard AOAC (2002) methods and starch was determined with a Megazyme assay kit (Megazyme International Ireland, Bray Business Park, Bray, Ireland) as described by McCleary et al. (1994). The digestibility coefficient of nutrients was calculated using the following equation:
Digestibility coefficient = 1 - [{digesta nutrient (g/kg DM)/digesta TiO2 (g/kg DM)}/{diet nutrient (g/kg DM)/diet TiO2 (g/kg DM)}]
Fresh intestinal content weighing ca. 1 g from the ileum and caeca was transferred into 15 mL MacCartney bottles containing 10 mL of an anaerobic broth. The suspension was homogenized for 2 min in CO2-flushed plastic bags using a bag mixer (Interscience, St. Norm, France) and then serially diluted in 10-fold increments in anaerobic broth according to the procedure described by Miller & Wolin (1974) and Engberg et al. (2004). One millilitre of the homogenized suspension was then transferred into 9 mL of anaerobic broth and serially diluted from 10-1 to 10-5 for ileum samples and 10-1 to 10-6 for caecal samples. From the last three diluted samples, 0.1 mL each was plated on the appropriate medium (10 mL) for enumeration of microbial populations.
Lactic acid bacteria were enumerated on MRS agar (Oxoid, CM0361) incubated under anaerobic conditions at 39 ºC for 48 h. Coliform (red colonies) and lactose-negative Enterobacteria (colourless colonies) were counted on MacConkey agar (Oxoid, CM 0007) incubated aerobically at 39 ºC for 24 h as red and collarless colonies, respectively. Lactobacilli were enumerated on Rogossa agar (Oxoid, CM 0627) after anaerobic incubation at 39 ºC for 48 h. Total anaerobic bacteria were counted using anaerobic roll tubes containing 3 mL of Wilkins-Chalgren anaerobic agar (Oxoid, CM 0619) incubated at 39 ºC for seven days. Number of C. perfringens (C.p) was counted on a Tryptose-Sulfite-Cycloserine and Shahidi-Ferguson Perfringens agar base (TSC & SFP) (Oxoid, CM 0587 OPSP) mixed with egg yolk emulsion (Oxoid, SR0047) and a Perfringens (TSC) selective supplement (Oxoid, SR0088E) according to the pour-plate technique, where plates were overlaid with the same agar after spreading the inoculum and incubated anaerobically at 39 ºC for 24 h. An anaerobic AnaeroGen' sachet (AN0025A, Oxoid Ltd, Hampshire, UK) was used to generate the anaerobic environment (<1% O2 and 9 - 13% CO2) for all anaerobically incubated agar plates. After incubation, colonies formed on the respective media were carefully counted, converted into logarithmic equivalents (log10) and expressed as number of colony forming units (CFU) per gram of wet intestinal content.
The data were analysed using the multiple regression options of SPSS, Version 17.0.0 (SPSS Inc, 2009) for the main effects of increasing drying temperature and enzyme supplementation (Morris, 1998). The data were also subjected to GLM analysis and reported where there was an interaction between drying temperature and microbial enzyme supplementation. Data for organic acid concentrations were log-transformed (log10 +1) prior to analyses. Separation of means within a significant effect was conducted using Duncan's Multiple Range Test (DMRT) through the post hoc procedure of SPSS. Differences between mean values were considered significant at P <0.05, unless otherwise specified.
Results
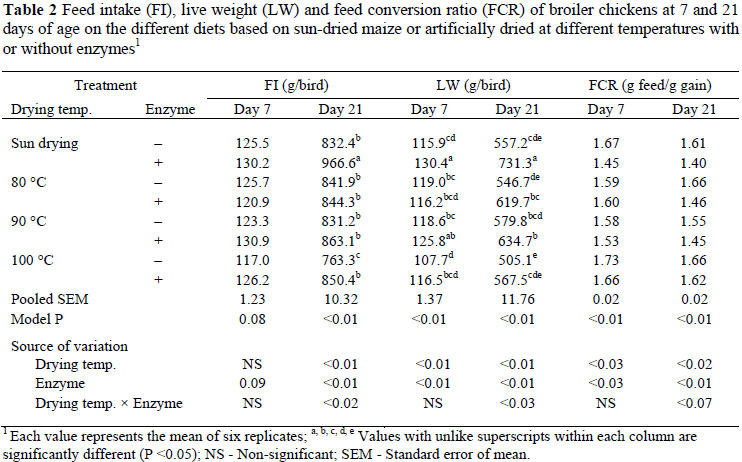
The gross response of chickens fed with the experimental diets is shown in Table 2. Up to seven days of age, there was no effect of grain drying temperature and enzyme supplementation on FI. However, the FI was marginally higher (P <0.08) in diets with enzymes than in unsupplemented diets (127.0 vs. 122.9 g/bird). Live weight at this age was affected (P <0.01, R2 = 0.25) by drying temperature but the trend was dependent on microbial enzyme supplementation. The microbial enzyme also improved (P <0.01, R2 = 0.25) LW only on the diets containing sun-dried grains. The FCR was significantly improved (P <0.03, R2 = 0.18) by increasing grain drying temperature up to 90 ºC. In general, FCR up to 7 days of age was better (P <0.03) on enzyme-supplemented diets than on diets without the microbial enzyme (1.56 vs. 1.64 g feed/g gain).
Feed intake up to day 21 declined (P <0.01, R2 = 0.39) with an increase in drying temperature of grains, while supplementation with microbial enzymes increased (P <0.01, R2 = 0.39) FI, but this was significant only on the diets based on sun-dried maize and the maize dried at 100 ºC (Table 2). The interaction in FI between drying temperature and enzyme was also significant (P <0.02). In general, enzyme supplementation resulted in a higher FI compared to non-enzyme diets (881.1 vs. 817.2 g/bird). At 21 d of age there was a reduction (P <0.01, R2 = 0.49) in LW with an increase in grain drying temperature while supplementation with microbial enzymes (P <0.01, R2 = 0.49) improved LW only on the diets containing sun-dried grains and grains dried at 90 ºC. Generally, LW was higher (P <0.01) in chickens on the enzyme-supplemented diets than on diets without enzymes (638.3 vs. 547.2 g/bird). Furthermore, the interaction between drying temperature and enzyme on LW was significant (P <0.03). The FCR at this age was reduced (P <0.02, R2 = 0.37) with an increase in grain drying temperature while the FCR improved (P <0.01, R2 = 0.37) on diets when supplemented with enzyme. Feed conversion ratio was also better (P <0.01) with diets containing enzymes than without enzymes (1.48 vs. 1.62 g feed/g gain).
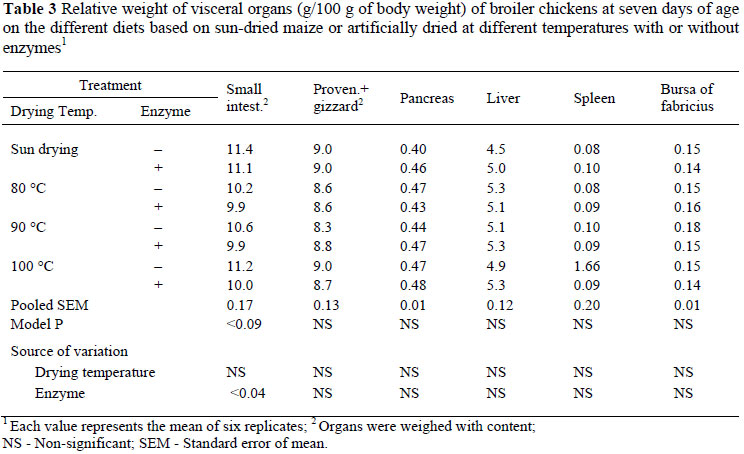
At day 7, the relative weight of the small intestine was not significantly affected by increase in grain drying temperature but was decreased (P <0.04, R2 = 0.11) in chickens on diets that were supplemented with microbial enzymes (Table 3). The relative weights of the proventriculus plus gizzard, pancreas, liver, spleen and bursa of fabricius were not significantly affected by an increase in grain drying temperature with or without enzyme supplementation.
At day 21, there was a significant increase (P <0.04, R2 = 0.10) in the relative weight of the small intestine with an increase in grain drying temperature while there was no change in the relative weight of this tissue due to supplementation with microbial enzymes (Table 4). The relative weight of the pancreas was not affected by grain drying temperature but was increased (P <0.01, R2 = 0.24) in chickens on diets supplemented with enzymes. At this age the relative weight of the liver was significantly increased (P <0.01, R2 = 0.14) with an increase in grain drying temperature but this effect was absent in chickens on diets containing microbial enzymes. There was no significant effect of grain treatments with or without enzyme on the relative weight of the spleen and bursa of fabricius at the same day of age.
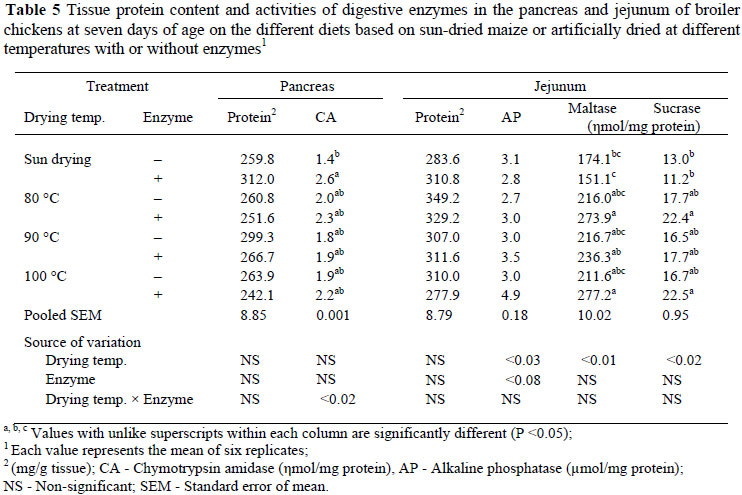
At 7 days of age pancreatic tissue protein content and chymotrypsin amidase activity were not affected by grain drying temperature or enzyme supplementation (Table 5). However, there was a significant interaction between the effects (P <0.02) of drying temperature and enzyme on the activity of chymotrypsin amidase.
In the jejunum there was no significant variation on tissue protein content at day 7 due to an increase in grain drying temperature, enzyme supplementation or any interactions between these factors (Table 5). The activity of alkaline phosphatase was significantly increased (P <0.03, R2 = 0.16) with an increase in grain drying temperature while supplementation with microbial enzymes also marginally increased (P <0.08, R2 = 0.16) the activity of this enzyme. The activity of maltase increased significantly (P <0.01, R2 = 0.18) with an increase in grain drying temperature but this effect was absent on diets supplemented with microbial enzymes. The activity of sucrase also increased (P <0.02, R2 = 0.15) with an increase in grain drying temperature but this activity was not affected by microbial enzyme supplementation.
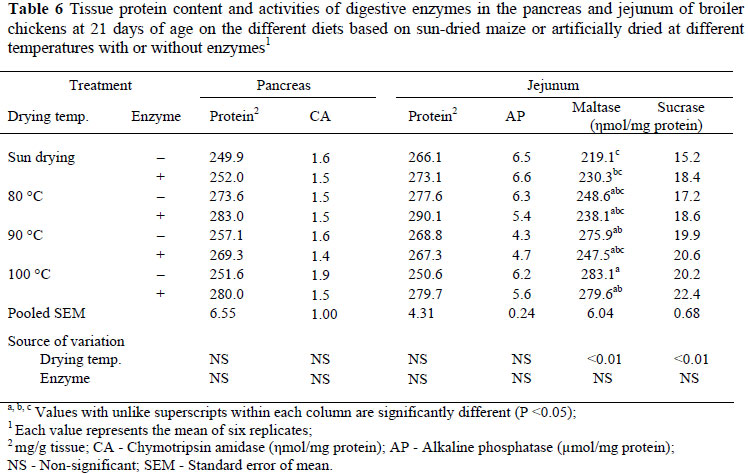
At day 21 the pancreatic protein content, chymotrypsin amidase activity, jejunal protein content and alkaline phosphatase activity were not significantly affected by increases in grain drying temperature, enzyme supplementation or interactions between the main factors (Table 6). The activity of maltase rose (P <0.01, R2 = 0.27) in line with an increase in grain drying temperature but was not affected by the microbial enzyme supplements. There was an increase (P <0.01, R2 = 0.18) in the activity of sucrase with an increase in drying temperature of grains but this activity was not affected in birds on diets supplemented with enzymes.
At 21 days of age the ileal digestibility of protein, gross energy and starch was not significantly changed by an increase in grain drying temperature or enzyme supplementation of diets (Table 7). However, the digestibility of protein was affected by the interaction (P <0.04) between drying temperature and enzyme, in which the digestibility of protein was higher on the diet containing grain dried at 100 ºC with enzymes, than on diet containing grain dried at 90 ºC without enzymes. In general, energy digestibility tended to increase in the diets containing the microbial enzyme supplements.
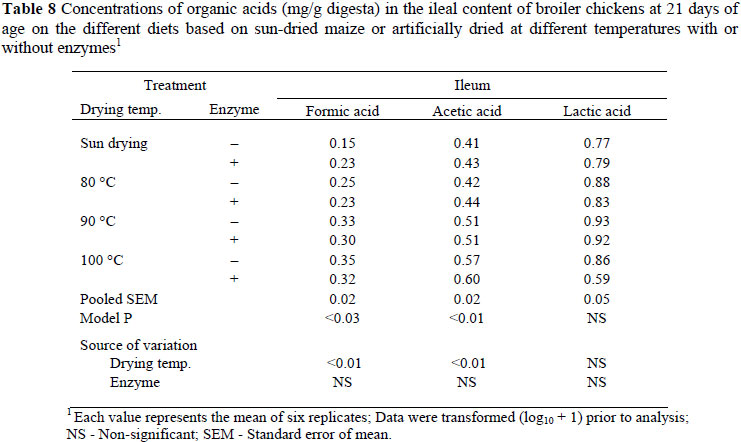
The concentrations of organic acids in the ileum of broiler chickens at day 21 are shown in Table 8. Formic acid content increased (P <0.01, R2 = 0.25) with an increase in drying temperature of grains used in the diets, but the concentration of formic acid was not changed when diets were supplemented with microbial enzymes. There was a significant increase (P <0.01, R2 = 0.34) in the concentration of acetic acid with increase in grain drying temperature but this was not affected by inclusion of microbial enzymes in the diets.
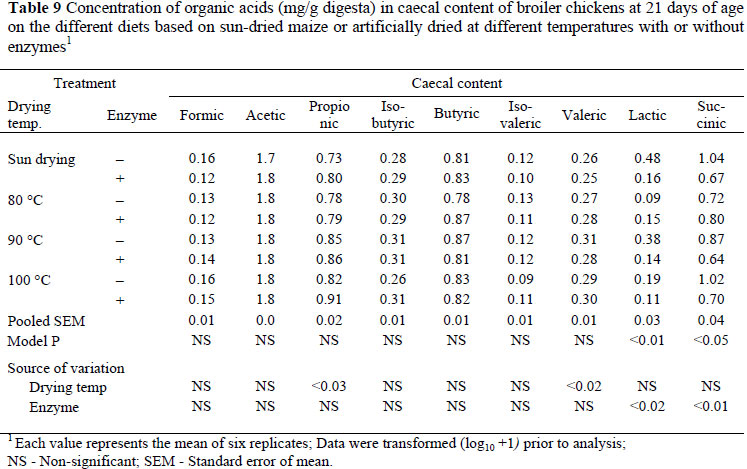
In the caecal content the concentrations of formic and acetic acids were not changed in response to an increase in drying temperature of grains used in the diets with or without enzyme supplementation (Table 9). The concentration of propionic acid increased (P <0.03, R 2 = 0.12) with increase in grain drying temperature but enzyme supplementation had no effect. The concentrations of isobutyric acid, butyric acid, and isovaleric acids were not affected by increase in drying temperature of grains, enzyme supplementation or interaction between the main factors. The concentration of valeric acid was increased (P <0.02, R2 = 0.12) by an increase in grain drying temperature but the concentration of this acid was not changed on diets supplemented with microbial enzymes. There was a significant reduction in the concentration of lactic acid (P <0.02, R2 = 0.14) and succinic acid (P <0.01, R2 = 0.15) on diets with the enzyme supplements but the concentrations of these acids were not affected by grain drying temperature.
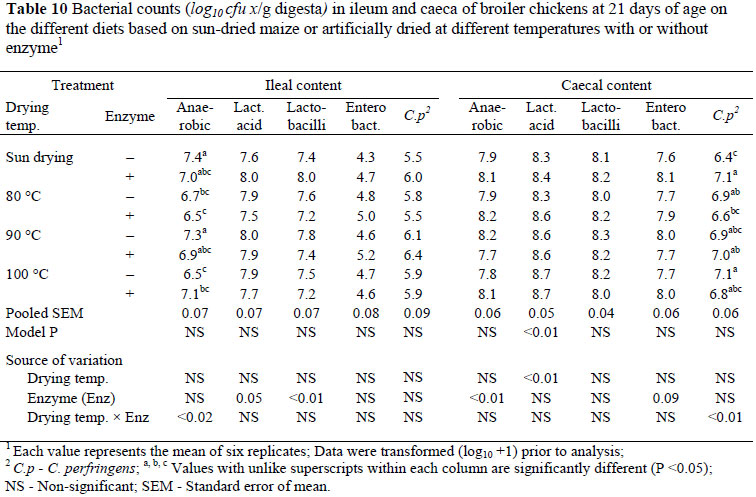
In the ileal content there was no significant effect of grain drying temperature or enzyme supplementation on total anaerobic bacterial count (Table 10). However, the interaction between drying temperature and enzyme supplementation on this bacterial count was significant (P <0.02). The populations of lactic acid bacteria (P <0.05, R2 = 0.10) and lactobacilli bacteria (P <0.01, R2 = 0.28) were decreased on diets supplemented with enzymes, but were not affected by increasing grain drying temperature. There were no significant changes in the populations of Enterobacteria and C. perfringens in the ileal content at 21 days of age as a result of variation in grain drying temperature, enzymes or their interactions.
In the caecal content the total anaerobic bacterial count was not significantly changed by increases in drying temperature of grains but the population of these bacteria was increased (P <0.01, R2 = 0.17) on diets when supplemented with microbial enzymes (Table 10). The population of lactic acid bacteria was increased (P <0.01, R2 = 0.18) as a result of an increase in grain drying temperature but this effect was absent on diets containing microbial enzymes. There were no significant changes in the populations of lactobacilli, Enterobacteria and C. perfringens in the caecal content at 21 days of age as a result of variation in grain drying temperature, enzyme supplementation or their interaction.
Discussion
The present study demonstrated that the maize dried either naturally in the sun or forced-dried at 90 ºC adequately supported chicken performance including FI, LW and FCR, up to 21 days of age. Enzyme supplementation also improved digestibility and bird performance on diets based on sun-dried and 90 ºC-dried grains. Therefore, sun drying, as practised in many areas is a suitable method for processing of maize which will be used as broiler feed. However, where sun drying is not possible due to climatic or other factors, artificial drying at 90 ºC is recommended, although enzyme supplementation may be needed to get the best results. This improvement is, perhaps, due to better chemical composition (Bhuiyan et al., 2010) and ultra-structural properties found in grain dried at 90 ºC and in sun-dried grain compared to grain dried at other high temperatures. The adverse effect of high temperature drying of maize have been reported previously (Kaczmarek et al., 2009) where authors also reported that exogenous enzymes were effective in partially ameliorating this inconsistency in energetic value if the grain is dried at more than 100 ºC. In the current study, the worst performance results (LW and FCR) were found in the batch dried at 100 ºC. This may be due to the decrease of its nutritional quality through high temperature drying, as reported previously (Brown, 1996; Iji et al., 2003).
The improvement in body weight through microbial enzyme supplementation of the diet based on 90 ºC-dried maize is consistent with our previous findings that maize dried at this temperature has a higher amylose concentration. In addition, Zanella et al. (1999) showed that enzyme supplementation improved BW and FCR ratio by 1.9 and 2.2%, respectively in broiler chickens.
In the present study, the relative weight of the small intestine varied according to the drying temperature of the grain in the diet and whether the diet was supplemented with enzymes or not. The relative weight of the small intestine of chickens fed diets containing sun-dried grain and without enzyme supplementation was found to be higher than that of birds on diets containing grain artificially dried at other temperatures with enzyme supplementation. These increases in the mass of the small intestine are approximately proportional to the increase in body weight, and might, therefore, be a general consequence of faster growth.
In this study, significant differences were observed in pancreatic protease (chymotrypsin amidase) activity due to interactions between drying temperature and enzymes, and higher pancreatic protease activity was found in birds on diets containing sun-dried grains with enzyme supplementation than in groups on unsupplemented diets. A dissimilar finding was reported by Mahagna et al. (1995), who observed a reduction in the activity of pancreatic enzymes in situ and in intestinal digesta as a result of supplementation with a microbial enzyme with proteolytic and amylolytic activities. The reasons for the discrepancy in these findings are unclear at this stage.
The activities of jejunal maltase and sucrase were significantly affected by drying temperature of grains but not by supplemental enzyme, and higher maltase and sucrase activities were found in birds on diets with grain dried at 80, 90 and 100 ºC compared to sun-dried grain. The increase in intestinal enzymes is stimulated mechanically by chime passing through the digestive tract (Duke, 1986). This may be due to higher DM content of diets at 80, 90 and 100 ºC than the sun-dried grain diet. The high activity of maltase and sucrase in the birds receiving these diets is probably related to substrate (maltose and sucrose) availability in the jejunum. The relationship between high-moisture grain dried at different temperatures and the activity of digestive enzymes has apparently not been examined previously. A reduction in jejunal tissue protein content was observed in this study at day 7, due to microbial enzyme application. Similar results have been reported by other workers (Danicke et al., 2000a; Iji et al., 2004), though grain with a lower moisture content was used.
There was a significant effect of drying temperature on starch digestibility, with higher starch digestibility found in sun-dried and 100 ºC-dried grain than in grain dried at other temperatures. However, the enzyme supplementation also enhanced starch digestibility in each case. The starch component of maize is considered to be highly digestible, but Brown (1996) reported that such starch may be resistant to digestion, and such resistance increases when grains are dried artificially. Starch digestion was completed in the hindgut, suggesting that some of the starch was indeed resistant. The enzyme mixture may have improved digestion of this fraction. The above phenomenon could warrant the use of exogenous enzymes (xylanase, glucanase, pectinase, cellulase, mannanase, galactanase) to increase the digestibility of nutrients found in maize (Meng & Slominski, 2005).
In this study it was found that protein and starch digestibility were enhanced in diets containing grains dried at 90 and 100 ºC. This may be due to the role that the addition of microbial phytase in poultry diets plays in increasing calcium and phosphorus availability, a finding that is well established in the literature. Phytic acid is a critical factor present in feedstuffs and it binds minerals, proteins, lipids and starch (Thompson & Yoon, 1984), reducing the digestibility of these nutrients for poultry (Sebastiana et al., 1997). Recent studies have also shown a beneficial effect of phytase addition to broiler diet on metabolizable energy and total amino acid digestibility (Namkung & Leeson, 1999; Ravindran et al., 1999). This also may be due to the activity of exogenous enzyme which improves the nutritive value of maize by hydrolyzing polysaccharides that are involved in the encapsulation of starch or protein, thereby rendering compounds available for digestion that were previously not accessible to endogenous enzyme (Bedford, 1996).
The changes in microbial populations may be due to a decrease in the quality of the grain. Although the ileal viscosity of birds was not measured in the present study, it is possible that there were major negative effects due to oven drying. However, in vitro viscosity has been measured using oven-dried grain in another study and no significant variation was found (Bhuiyan et al., 2010). There was a significant effect on the number of lactobacilli with a higher lactobacilli population found in the diet without enzyme supplementation but only to a lesser extent. Carbohydrates that remain undigested in diets without microbial enzymes are fermented in the hind gut, leading to an increase in microbial populations. The populations of other species such as lactic acid bacteria, enterobacteria and C. perfringens were not affected by grain quality or enzyme supplementation, although the count of C. perfringens, the causative agent of necrotic enteritis, was slightly increased.
In caecal content there was a significant variation on total anaerobic count with higher total anaerobic bacteria found in chickens on enzyme-supplemented diets than those on the unsupplemented diets. This is an unusual occurrence as microbial enzymes tend to increase ileal digestion and reduce caecal fermentation (Apajalahti & Bedford, 1999; Fernandez et al., 2000; Apajalahti et al., 2004).
In the current study there was a significant effect of drying temperature on the concentration of formic acid and acetic acid in the ileum of broiler chickens, and the higher concentration of formic acid was associated with grain dried at 100 ºC rather than sun-dried grain. This result suggests that the grains dried at 80 and 100 ºC were fermented more by the ileal microflora. Therefore, the growth and activity of formic acid- and acetic acid-producing bacteria in the ileum are likely promoted by the grain dried at 80 and 100 ºC. The composition of microbiota or increase in their digestive activity can have positive as well as negative effects on the host (Gabriel et al., 2006). The positive effects are competitive exclusion, SCFAs production and release of nutrients that can be absorbed in the intestine. Similar significant changes in the concentration of lactic acid were found due to enzyme supplementation, with a higher lactic acid content found in birds on diets without enzyme supplementation than on supplemented diets. Lactate was the most predominant organic acid in the ileal content, while acetic acid was predominant in the caecal digesta. Lactic acid is an electron sink, further oxidized to other SCFAs such as acetic, propionic and butyric acids in the caeca due to longer retention time of the caecal digesta (Vidanarachchi, 2006). An increased SCFA production through bacterial fermentation of undigested carbohydrates such as resistant starch also results in lower pH, which is an accepted mechanism for the inhibition of acidophilic bacteria such as enterobacteria and C. perfringens in broiler chickens (Terada et al., 1994; Orban et al., 1997). In the current study, however, no reductions in the populations of enterobacteria or C. perfringens were found due to grain drying or enzyme supplementation.
Conclusions
It may be concluded from the present study that diets based on sun-dried maize or maize dried at 90 ºC gave comparatively better performance in terms of feed intake, live weight and FCR of broilers than diets based on maize dried at other temperatures. Moreover, it may be inferred that there was a significant positive response to supplementation with microbial enzymes. The relative weight of the small intestine and liver, the bacterial populations and starch digestibility were also higher in birds on diets based on sun-dried maize compared to those based on artificially dried grain. Considering all of the above points in this experiment, it may be assumed that, for broiler chickens, there is little or no difference in the nutritive value of sun-dried grain compared to grain artificially dried at 90 ºC. Supplementation with a suitable enzyme may also be of some benefit in terms of visceral organ development, performance, protein digestibility and gut microflora.
Acknowledgement
This project was financially supported by the University of New England, Australia and Danisco Animal Nutrition, UK.
References
AOAC, 2002. Official Methods of Analysis (17th ed.). Association of Official Analytical Chemists, Inc., Maryland, USA. [ Links ]
Apajalahti, J. & Bedford, M.R., 1999. Improve bird performance by feedings its microflora. Wrld Poult. Sci. J. 15, 20-23. [ Links ]
Apajalahti, J., Kettunen, A. & Graham, H., 2004. Characteristics of gastrointestinal microbial communities with special reference to the chicken. Wrld Poult. Sci. J. 60, 223-232. [ Links ]
Bedford, M.R., 1996. The effects of enzymes on digestion. J. App. Poult. Res. 5, 370-378. [ Links ]
Bedford, M.R. & Moran, A.J., 1996. The use of enzymes in poultry diets. Wrld Poult. Sci. J. 52, 419-427. [ Links ]
Bhuiyan, M.M., Iji, P.A., Islam, A.F. & Mikklesen, L.L., 2010. Variation in nutrient composition and structure of high-moisture maize dried at different temperatures. Paper presented at the Aust. Poult. Sci. Symp., Sydney, Australia 21, 99-102. [ Links ]
Boros, D., Slominski, B.A., Guenter, W., Campbell, L.D. & Jones, O., 2004. Wheat by-products in poultry nutrition. Part II. Nutritive value of wheat screenings, bakery by-products and wheat mill run and their improved utilization by enzyme supplementation. Can. J. Anim. Sci. 84, 429-435. [ Links ]
Bradford, M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-254. [ Links ]
Brown, I., 1996. Complex carbohydrates and resistance starch. Nutr. Rev. 54, S115-S119. [ Links ]
Cowieson, A.J., 2005. Factors that affect the nutritional value of maize for broilers. Anim. Feed Sci. Technol. 119, 293-305. [ Links ]
Danicke, S., Bottcher, W., Jeroch, H., Thielebein, J. & Simon, O., 2000. Replacement of soybean oil with tallow in rye-based diets without xylanase increases protein synthesis in small intestine of broiler. J. Nutr. 130, 827-834. [ Links ]
Duke, G.E., 1986. Alimentary canal: secretion and digestion, special digestive functions and absorption. In: Avian Physiology. Ed. Sturkie, P.D., Springer-Verlag, New York. pp. 289-302. [ Links ]
Engberg, R.M., Hedemann, M.S., Steenfeldt, S. & Jensen, B.B., 2004. Influence of whole wheat and xylanase on broiler performance and microbial composition and activity in the digestive tract. Poult. Sci. 83, 925-938. [ Links ]
Fernandez, F., Sharma, R., Hinton, M. & Bedford, M.R., 2000. Diet influences the colonisation of Campylobacter jejuni and distribution of mucin carbohydrates in the chick intestinal tract. Cell Mol. Life Sci. 57, 1793-1801. [ Links ]
Gabriel, I., Lessire, M., Mallet, S. & Guillot, J.F., 2006. Microflora of the digestive tract: Critical factors and consequences for poultry. Wrld Poult. Sci. J. 62, 499-511. [ Links ]
Holdsworth, E.S., 1970. The effect of vitamin D on enzyme activities in the mucosal cells of the chick small intestine. J. Membr. Bio. 3, 43-53. [ Links ]
Iji, P.A., Saki, A. & Tivey, D.R., 2001. Body and intestinal growth of broiler chicks on a commercial starter diet. 2. Development and characteristics of intestinal enzymes. Br. Poult. Sci. 42, 514-522. [ Links ]
Iji, P.A., Khumalo, K., Slippers, S. & Gous, R.M., 2003. Intestinal function and body growth of broiler chickens on diets based on maize dried at different temperatures and supplemented with a microbial enzyme. Reprod. Nutr. Dev. 43, 77-90. [ Links ]
Iji, P.A., Khumalo, K., Slippers, S. & Gous, R.M., 2004. Intestinal function and body growth of broiler chickens on maize-based diets supplemented with mimosa tannins and a microbial enzyme. J. Sci. Food Agric. 84, 1451-1458. [ Links ]
Jensen, M.T., Cox, R.P. & Jensen, B.B., 1995. Microbial production of skatole in the hind gut of pigs fed different diets and its relation to skatole deposition in backfat. Anim. Sci. 61, 293-304. [ Links ]
Kaczmarek, S., Bochenek, M., Jozefiak, D. & Rutkowski, A., 2009. Effect of enzyme supplementation of diets based on maize or hominy feed on performance and nutrient digestibility in broilers. J. Anim. Feed Sci. 18, 113-123. [ Links ]
Mahagna, M., Nir, I., Larbier, M. & Nitsan, Z., 1995. Effect of age and exogenous amylase and protease on development of the digestive tract, pancreatic enzyme activities and digestibility of nutrients in young meat-type chicks. Reprod. Nutr. Dev. 35, 201-212. [ Links ]
McCleary, B.V., Solah, V. & Gibson, T.S., 1994. Quantitative measurement of total starch in cereal flours and products. J. Cereal Sci. 20, 51-58. [ Links ]
McDonald, P., Edwards, R.A., Greenhalgh, J.F.D. & Morgan, C.A., 1995. Animal Nutrition, Longman Scientific and Technical. Harlow, UK. [ Links ]
Mcpherson, G.A., 1985. LOWRY PROGRAM: Elsevier-BIOSOFT, 68 Hills Rd, Cambridge, CB2, 1LA, UK. [ Links ]
Meng, X. & Slominski, B.A., 2005. Nutritive values of corn, soybean meal, canola meal and peas for broiler chickens as affetced by a multicarbohydrase preparation of cell wall degrading enzymes. Poult. Sci. 84, 1242-1251. [ Links ]
Miller, T.L. & Wolin, M.J., 1974. A serum bottle modification of the Hungate technique for cultivating obligate anaerobes. Appl. Envir. Microbiol. 27, 985-987. [ Links ]
Morris, T.R., 1998. Experimental Design and Analysis in Animal Sciences. University of Reading, Department of Agriculture, Reading, UK. [ Links ]
Namkung, H. & Leeson, S., 1999. Effect of phytase enzyme on dietary nitrogen-corrected apparent metabolizable energy and the ileal digestibility of nitrogen and amino acids in broiler chicks. Poult. Sci. 78, 1317-1319. [ Links ]
National Health and Medical Research Council, 1990. Australian Code of Practice for the care and use of animals for scientific purposes, Commonweath Scientific and Industrial Research Organisation, Australian Agricultural Council (Austalian Govt. Pub. Service). Canberra, National Health and Medical Research Council. [ Links ]
National Research Council, 1994. Nutrient Requirements of Poultry, 9th Revised Ed., National Research Council. National Academy Press, Washington, D.C., USA. [ Links ]
Nitsan, Z., Dror, Y., Nir , I. & Shapira, I., 1974. The effects of force-feeding on enzymes of the liver, kidney, pancreas and digestive tract of chicks. Br. J. Nutr. 32, 241-247. [ Links ]
Noy, Y. & Sklan, D., 1994. Digestion and absorption in the young chick. Poult. Sci. 74, 366-373. [ Links ]
Orban, J.I., Patterson, J.A., Sutton, A.L. & Richards, G.N., 1997. Effect of sucrose thermal oligosaccharide caramel, dietary vitamin-mineral level, and brooding temperature on growth and intestinal bacterial populations of broiler chickens. Poult. Sci. 76, 482-490. [ Links ]
Panigrahi, S., Bestwick, L.A., Davis, R.H. & Wood, C.D., 1996. The nutritive value of stackburned yellow maize for livestock: tests in vitro and in broiler chicks. Br. J. Nutr. 76, 97-108. [ Links ]
Peric, L., Kovcin, S., Stanacev, V. & Milosevic, N., 2002. Effect of enzymes on broiler chick performance. Buletinul USAMV, Cluj-Napoca, Romania. 57, 245-249. [ Links ]
Ravindran, V., Cabahug, S., Ravindran, G. & Bryden, W.L., 1999. Influence of microbial phytase on apparent ileal amino acid digestibility of feedstuffs for broilers. Poult. Sci. 78, 699-706. [ Links ]
Sebastiana, S., Touchburn, S.P., Chavez, E.R. & Lague, P.C., 1997. Apparent digestibility of protein and amino acids in broiler chickens fed a corn-soybean diet supplemented with microbial phytase. Poult. Sci. 76, 1760-1769. [ Links ]
Serviere-Zaragoza, E., Navarrete Del Toro, M.A. & Garcia-Carreno, F.L., 1997. Protein hydrolyzing enzymes in the digestive system of the adult Mexican blue abalone, Haliotis fulgens (Gastropoda). Aquaculture 157, 325-336. [ Links ]
Shirazi-Beechey, S.P., Smith, M.W., Wang, Y. & James, P.S., 1991. Postnatal development of lamb intestinal digestive enzymes is not regulated by diet. J. Physiol. 437, 691-698. [ Links ]
Short, F.J., Gorton, P., Wiseman, J. & Boorman, K.N., 1996. Determination of titanium dioxide added as an inert marker in chicken digestibility studies. Anim. Feed Sci. Technol. 59, 215-221. [ Links ]
SPSS Inc. 2009. Version 17.0.0. SPSS Inc.,Chicago, USA. [ Links ]
Taylor, J.R.N. & Belton, P.S., 2002. In: Pseudocereals and Less Common Cereals. Eds Belton, P.S. & Taylor J.R.N., Springer, Berlin, Germany. pp. 25-91. [ Links ]
Terada, A., Hara, H., Sakamoto, J., Sato, N., Takagi, S., Mitsuoka, T., Mino, R., Hara, K., Fujimori, I. & Yamada, T., 1994. Effects of dietary supplementation with lactosucrose (4-b-D-galacosylsucrose) on caecal flora, caecal metabolites and, performance in broiler chickens. Poult. Sci. 73, 1663-1672. [ Links ]
Thompson, L.U. & Yoon, J.H., 1984. Starch digestibility as affected by polyphenol and phytic acid. J. Food Sci. 49, 1228-1229. [ Links ]
Vidanarachchi, J.K., 2006. Regulation of intestinal microflora and productivity of broiler chickens by prebiotic and bioactive plant extracts. PhD thesis, The University of New England, Armidale, NSW, Australia. [ Links ]
Zanella, I., Sakomura, N.K., Silversides, F.G., Fiqueirdo, A. & Pack, M., 1999. Effect of enzyme supplementation of broiler diets based on corn and soyabeans. Poult. Sci. 78, 561-568. [ Links ]
# Corresponding author. E-mail: piji@une.edu.au


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}