










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.40 n.4 Pretoria Jan. 2010
Influence of growth stage at harvest on fermentative characteristics of Panicum maximum silage
W.A. van Niekerk#; Abubeker Hassen; F.M. Bechaz
Department of Animal and Wildlife Sciences, University of Pretoria, Pretoria 0002, South Africa
ABSTRACT
An experiment was conducted to investigate the influence of stage of growth at harvest on fermentative characteristics of Panicum maximum silage. The treatments were three different growth stages (early vegetative, boot and full bloom stage) that were ensiled directly or wilted prior to ensiling. Directly ensiled and wilted forage material were mixed prior to ensiling with molasses at 12 and 8 kg/ton dry matter, respectively. Thereafter, each treatment was ensiled in 12 one litre glass jars (bottles). From each treatment, samples of the silage were taken from three bottles at 0, 7, 21 and 120 d post-ensiling for analysis of fermentative characteristics. Growth stage in directly cut silage had no effect on silage pH between days 0 and 21. In contrast, within the wilted groups, a lower pH was observed at day 7 when the plants were harvested at the boot stage than at an early vegetative or full bloom stage. In both directly cut as well as prior wilted silage, a higher lactic acid concentration was recorded on day 7 in the boot stage silage compared to the full bloom stage silage, but on day 21 the boot stage had a lower lactic acid concentration than that of the full bloom stage silage. Growth stage had no effect on the acetic acid concentration on day 7. However, on days 21 and 120 the full bloom stage had a higher acetic acid concentration when the material was directly ensiled. In contrast, in prior wilted silage a higher acetic acid concentration was found in the early vegetative stage silage than in the full bloom stage silage at 120 days post-ensiling. Total nitrogen was lower in the full bloom stage silage than in the early vegetative and boot stage silages. A higher level of ammonia nitrogen was recorded at days 7 and 21 for the early vegetative stage silage as compared to the boot stage silage. A similar trend was revealed in the prior wilted groups between days 0 and 21. Harvesting at the boot growth stage consistently resulted in a good fermentation process with desirable fermentation end products (low pH and higher lactic acid concentration) up to a period of 21 days, but when compared at 120 days post-ensiling the differences between boot and bloom stages were not clearly observed in terms of fermentative attributes, probably due to the confounding effect of undesirable fermentation by entrobacteria or yeast.
Keywords: Nutritive value, P. maximum, preservation, tropical grass
Introduction
Panicum maximum (Guinea grass) is indigenous to subtropical areas of southern Africa. It is tolerant of shade and fire and produces high yields of palatable fodder during the warm season, but the nutritive value declines rapidly with increasing stages of maturity (Aganga & Tshwenyane, 2004). The removal of surplus forage (for silage) will ensure a succession of high quality regrowth for grazing and can be used to prepare an area for the production of foggage (De Figueiredo, 2000). The silage could supply quality roughages during late winter and early spring periods when the pasture is low in nutritive value, but high in dry matter (DM) concentration (Rethman, 1983; Tainton, 2000). Silage has several advantages over hay for the conservation of fodder. It does, however, require a good microbial fermentation process to produce a good quality fodder. For tropical grass species, desirable anaerobic fermentation might not take place to ensure a high level of lactic acid production with a low pH as well as low acetic acid and ammonia nitrogen (NH3-N) concentrations (Catchpoole & Henzell, 1971; Sujatha et al., 1986; Imura et al., 2001). This is because tropical grass species generally have a higher buffering capacity (BC) and/or lower water soluble carbohydrate (WSC) levels to support growth and multiplication of lactic acid fermenting bacteria than ideal silage crops. The actual economic value of silage is, however, dependent on the type, extent and level of fermentation and the fermentation end products of the silage. All of these are influenced by the type and quality of the forage crop as well as harvesting and ensiling techniques, which subsequently determine the preservation process, nutritive value and voluntary feed intake by the animal (Wilkins et al., 1971; Demarquilly, 1973). The end products of silage fermentation are often monitored to assess silage quality. This paper reports the influence of growth stages at harvest on the fermentation process of Panicum maximum silage ensiled either directly or wilted prior to the ensiling process.
Materials and Methods
The study was conducted at Potchefstroom Agricultural College, North West Province, South Africa (altitude 1338 m above sea level, 26º42'13.07" S and 27º0.5'52.88" E). The area is an exclusive summer rainfall area with dry autumns and winters. The top soil had a pH (in water) of 6.7 ± 0.26, a pH (in KCl) of 5.7 ± 0.19, and contained P at 60 ± 27.5 mg/kg, K at 53 ± 12.1 mg/kg, Ca at 494 ± 34.1 mg/kg, Mg at 359 ± 45.1 mg/kg and Na at 14 ± 2.8 mg/kg.
Panicum maximum cv. Gatton was established in February on a 0.8 ha area under irrigation. During August at the end of the first winter, the pasture was harvested and materials removed. During the subsequent growing season, the pasture was top dressed with 100 kg nitrogen (N)/ha [350 kg of lime ammonium nitrate (LAN)/ha] in October and with 50 kg N/ha in January. The pasture received 25 mm water weekly (rainfall and/or irrigation) during the growing season.
Before cutting the plants for ensiling purpose, 10 tufts were clipped at a height of 10 cm above the ground. The material from these tufts was pooled and a sample was oven dried at 100 ºC for 24 h to determine the initial DM concentration of the herbage. This was essential to be able to estimate the amount of sugar that had to be added to each treatment during ensiling. Subsequently the third re-growth of the P. maximum pasture was mown at the early vegetative, boot and full bloom stages of growth that corresponded to 28, 37 and 53 days regrowth at harvesting, respectively. The harvested forage material was collected fresh and fed into the maize silage harvester in order to reduce the size to a length of 12 ± 1 mm. There-upon, depending on treatment, about 640 g of the chopped material was either directly ensiled in 1 L jars or wilted for 5 - 6 h to a DM concentration of ± 30% prior before ensiling. The directly ensiled treatments were mixed with 12 kg molasses additive/ton DM while the prior wilted material was mixed with 8 kg molasses/ton DM. Each treatment was ensiled in 12 glass bottles (fruit jars), each with 1 L volume capacity. The bottles were filled and compacted in order to eliminate oxygen and sealed with a tight cover to obtain sustainable anaerobic conditions. The mini-silos were then stored at room temperature (22 ºC) for periods of 0, 7, 21 and 120 days. The Day 0 sample was immediately stored in a deep freezer for later analysis. The DM, WSC concentration as well as the BC of the fresh as well as the wilted grass samples were determined before ensiling of the grass. Thereafter, three mini-silos each from both the directly cut and wilted silage treatments were opened on days 7, 21 and 120 post ensiling. One third of the ensiled forage material from each sample was oven dried at 60 ºC and analysed in duplicate to determine their DM, organic matter (OM) and N concentrations according to the AOAC (2000) methods. The second portion of the silage sample was stored in a freezer at -13 ºC until its pH was measured and the concentration of lactic acid, acetic acid and NH3-N determined. The remaining silage was discarded after sampling. About 40 g of a wet silage sample was placed in a 1000 mL container and 160 mL distilled water and saturated mercuric chloride were added, the latter as a preservative, and stored overnight at 7 ºC. The extract was shaken for 6 h at 180 rpm and filtered through four layers of cheesecloth to remove plant matter. The supernatant was then transferred to 250 mL plastic bottles and stored in a freezer (-15 ºC) for later determination of pH, lactic acid, acetic acid and NH3-N concentrations. The pH was measured after allowing the samples to stand overnight at room temperature. Buffering capacity was determined following the procedure of Payne & McDonald (1966) and WSC concentration using the procedure of McDonald & Henderson (1964). The NH3-N concentration was determined using a Technicon Auto Analyser II following the procedure of Davie (1989). Volatile fatty acid and lactic acid were determined from 3 mL of the silage extract following the procedure of Payne & McDonald (1966) and Pryce (1969), respectively.
The general linear model (GLM) procedure of SAS (2001) was used to test statistical differences between the different growth stages and between sampling periods. The influence of stage of maturity and sampling periods were analysed separately for the directly cut and wilted silage samples. Significance of difference between least squares means was determined by Bonferroni's test (Samuels, 1989).
Results and Discussion
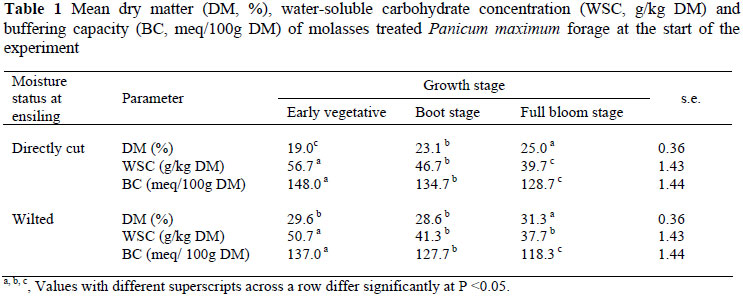
In this study the concentrations of WSC and BC for directly cut as well as wilted silage harvested at the early vegetative stage were higher than at the boot and full bloom stages (Table 1). However, a higher BC for the early vegetative stage harvest means that more acid molecules would be needed to decrease the pH of the silage compared to the boot stage or full bloom stages (Sollenberger et al., 2004). In this study, the range of BC recorded for P. maximum (1183 - 1480 meq/kg DM) was generally higher than values reported by McDonald (1973) for forage maize (200 meq/kg DM), orchard grass (300 meq/kg DM), ryegrass (250 - 400 meq/kg DM), lucerne (400 - 600 meq/kg DM) and clover (500 - 600 meq/kg DM).
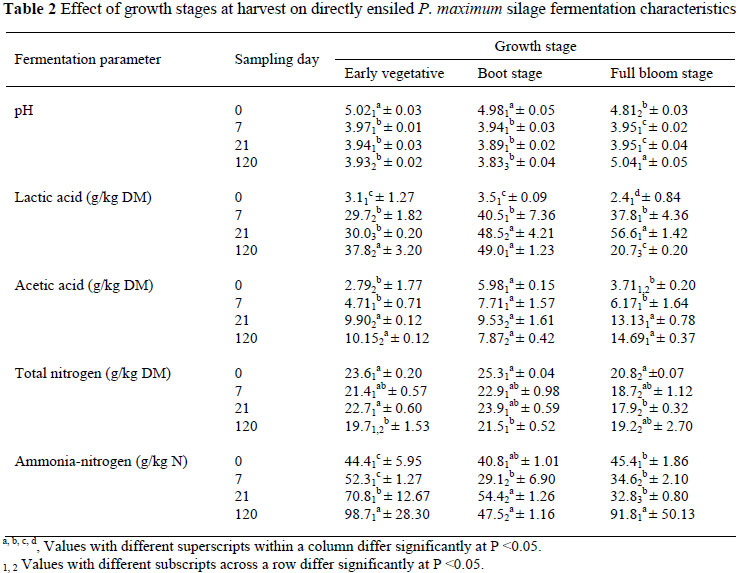
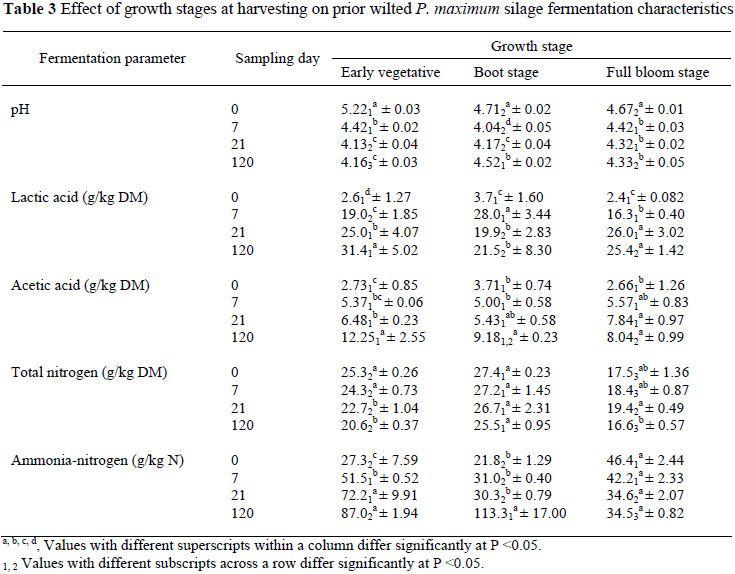
For all directly cut silages, the pH declined (P <0.05) rapidly (below 4) regardless of growth stage (Table 2) while, within the prior wilted treatments the decrease in pH was more gradual for the early vegetative and full bloom stages up to day 21 (Table 3). According to Meeske et al. (2000) and Schroeder (2004) the rate of pH decrease influences the quality of the silage and it appears that the BC and lower WSC concentration recorded at the early vegetative and the full bloom stages, respectively, might have contributed to the gradual decrease in pH recorded for wilted silage harvested at the vegetative and full bloom stages. Kung & Shaver (2001) suggested that good quality silage should have a pH value of less than 4.2, and this should be attained between days 7 and 21 post ensiling. The same authors reported that at a pH of ±4, the silage is well preserved and microbial growth will be completely inhibited. The pH needed for preservation also depends on the DM content. Critical pH values for fermentation are pH 4.2, pH 4.45, pH 4.75 and pH 5 at DM levels of 20%, 30%, 40% and 50%, respectively (Weissbach, 1996). In contrast, the slower rate of decrease in pH will provide more time for the growth of clostridia, which grow well at pH values of 5 and higher (Jonsson, 1991; Meeske et al., 1999; Schroeder, 2004). The rate of decrease in pH is also determined by the epiphytic bacteria present in the crop prior to ensiling and the level of fermentable sugar (water-soluble carbohydrates), which in turn depends on stage of maturity (Meeske et al., 2000; Schroeder, 2004). In this study maturity in particular had a tremendous impact on the moisture level of directly cut silage. At day 0 a higher moisture level was recorded for the early vegetative stage than the boot or full bloom stage forage material/silage (Table 1), and this agrees with previous results reported for directly cut forage crops such as maize silage (Schroeder, 2004). In this study, the gradual increase in pH at 120 days post ensiling for directly cut silage harvested at the full bloom harvest stage is possibly an indication of undesirable fermentation from micro-organisms such as clostridia. According to Schroeder (2004), the Clostridia are more likely to occur in forage ensiled at moisture levels higher than 70% than below, which results in higher pH values (Anonymous, 1995; Meeske et al., 2000). Clostridia are anaerobic bacteria, which break down protein and produce butyric acid, which will lower the palatability and intake of silage (Meeske et al., 2000). Unfortunately the concentration of butyric acid was not recorded in this experiment.
The lactic acid production was significantly affected by the stage of growth at harvesting (Table 2). Except for the directly cut silage on day 120 (Table 2), the boot and full bloom stages showed higher (P <0.05) lactic acid concentrations than the early vegetative stages on days 7 and 21. A higher concentration of lactic acid (in a range of 60 - 100 g/kg DM) is desirable for good quality silage, as this retains more DM and energy and preserves the silage well for a long period of time (Schroeder, 2004; Bethard, 2006). At day 21, the full bloom stage silage resulted in a higher (P <0.05) lactic acid concentration than the boot stage silage, but on day 120 the concentration of lactic acid for the full bloom stage decreased in the directly cut silage. The reduction in lactic acid could be partly the result of the degradation of lactic acid to butyric acid, CO2 and water by the action of Clostridium tyrobutyricum and/or yeasts (Oude Elferink et al., 2000).
In wilted silages, however, lactic acid production increased (P <0.05) gradually up to day 120 for the early vegetative and full bloom stages (Table 3). This increase in lactic acid concentration with fermentation time is in accordance with other studies (Aguilera et al., 1997). For prior wilted treatments, however, the lactic acid concentration declined significantly for the boot stage at days 21 and 120 post ensiling. The decrease in lactic acid concentration was associated with a gradual increase in pH and NH3-N concentration for the same treatments (Table 3). This might be due to clostridial fermentation as the end products have relatively higher pH values, and will result in higher pH values (≈ 5) (Schroeder, 2004). Unfortunately the butyric acid concentration was not measured in our experiment to verify this. However, for directly cut silages, boot stage harvesting resulted in a higher (P <0.05) lactic acid concentration at 120 days compared to harvesting at early vegetative stage or full bloom stages. This means that the directly ensiled silage boot stage is the optimum stage of harvest, since it consistently resulted in a higher lactic acid concentration, a lower NH3-N concentration and lower pH values during the fermentation process. For prior wilted silage, the early vegetative stage seems to be better preserved as compared to the boot or full bloom stages when evaluated in terms of silage pH and lactic acid concentration. However, the recorded higher NH3-N as well as the higher acetic acid concentrations for the early vegetative stage suggests the presence of clostridia activity. At day 120 the full bloom stage has lower pH and NH3-N values compared to the boot stage. By delaying harvest there is a trend of a reduction in WSC and BC as well as an increase in DM concentration. The relative reduction in WSC associated with maturity might have contributed partially for the poor preservation at the boot stage, while at the same time the observed reduction in BC and increase in DM concentration at the full bloom stage have played a positive role towards better preservation. This is because the lower the moisture content of the crop, the higher the pH at which anaerobic stability is reached. Moreover, forages with a high BC are undesirable in silage because more WSC must be used to produce the additional acid required for a reduction in pH which is necessary to inhibit clostridia activity and enhance lactic acid bacteria growth for good preservation. According to McDonald et al. (2002), a DM below 25% favours clostridial activity and DM losses as a results of effluent may also occur. On the other hand if the forage is wilted prior to ensiling, organic acids are lost in the wilting process and this reduces the BC of the plants which improves the ensiling process. In addition, a higher DM concentration (as for the wilted silage) may result in higher DM intake by animals.
With regard to acetic acid concentrations, at day 7 harvesting stage had no significant effect on the level of acetic acid production (Table 2, 3) at these stages. However, the highest acetic acid concentration (14.6 g/kg DM) recorded in this study was lower than the upper limit (20 g acetic acid/kg silage DM) suggested for good quality silage by Ward (2000). Within the directly cut silage, acetic acid concentration was lower (P <0.05) in the early vegetative and boot stages compared to the full bloom stage. Although acetic acid production is somewhat desirable as it can be utilised by ruminants (Schroeder, 2004), it is an indicator of slower fermentation and its higher concentration in silage reduces dry matter intake (Erdman, 1993).
In this study, on days 0 to 21 the N concentration of directly cut full bloom stage silage was the lowest (Table 2), and the same trend was recorded for the wilted silage. This is probably due to a decrease in the N concentration of the forage as the crop matures. Similar to silage pH and lactic acid concentration, NH3-N concentration can be used as an indicator of an undesirable type of fermentation process. Although up to 50% of the total plant protein may be broken down to NH3-N and amines by aerobic bacterial action, the extent of proteolysis depends on the rate of pH decline in the silage (Schroeder, 2004). The acid environment eventually reduces the activity of proteolysis. A more rapid decline in pH would limit degradation and de-amination of forage proteins and reduce NH3-N production (Woolford, 1984; Jonsson, 1991; Meeske et al., 1999; Schroeder, 2004). In contrast, high NH3-N levels (>10% of the total N) may lead to reduced silage intake and this is normally associated with undesirable fermentation by enterobacteria and clostridia (Kung & Shaver, 2001). In this study, the NH3-N level was the lowest (P <0.05) at days 7 and 21 for silages harvested at the boot stage compared to that harvested at the early vegetative or full bloom stages. This pattern corresponded well with a lower silage pH and higher lactic acid concentrations (Tables 2 and 3). However, at day 120 the proportion of NH3-N for the boot stage that was ensiled after wilting was exceptionally high. The exact cause is not clear, but it could be related to clostridia activity. Infiltration of air in the silo might also be an alternative cause that encourages yeast fermentation, which could lead to a higher N breakdown (Meeske et al., 2000) and a higher acetic acid production (McDonald et al., 2002). Insufficient viable lactic acid bacteria on the grass at ensiling could also result in a delay in the drop of pH, higher nutrient losses and silage of a poorer palatability (Woolford, 1984) by favouring the undesirable bacteria species fermentation (McDonald et al., 2002).
Conclusions
In this study, directly cut silage harvesting at the boot stage of growth resulted in a desirable fermentation end product (lower pH and higher lactic acid and total N) up to a period of 120 days. This helped to maintain the nutritive value of the silage during the preservation period. At the same time the breakdown of protein by clostridia under anaerobic conditions was very low for the boot stage growth, which could minimize nutrient loss. Desirable fermentation end products will in turn ensure a higher DM (nutrient) concentration and an additional benefit in terms of improved animal performance.
References
Aganga, A.A. & Tshwenyane, S., 2004. Potentials of Guinea grass (Paniucm maximum) as forage crop in livestock production. Pak. J. Nutr. 3, 1-4. [ Links ]
Aguilera, A., Pérez-Gil, F., Grande, D. de la Cruz, I. & Juárez, J., 1997. Digestibility and fermentative characteristics of mango, lemon and corn stover silages with or without addition of molasses and urea. Small Rumin. Res. 26, 87-91. [ Links ]
Anonymous, 1995. The PIONEER Forage Manual- A Nutritional Guide. Pioneer H-Bred international, Inc. Des Moines, Iowa, USA. pp. 21. [ Links ]
AOAC, 2000. Official Methods of Analysis (15th ed.). Association of Official Analytical Chemists, Inc., Washington D.C., USA. [ Links ]
Bethard, G., 2006. Forage management from a nutritionist perspective. In: The Dairy Professional News letter. Volume 2, issue 6. Monsanto Dairy Business for Industry Professionals. Accessed: http://www.thedairyprofessional.com/midwest/articles/issue6.htm [ Links ]
Catchpoole, V.R. & Henzell, E.F., 1971. Silage and silage making from tropical herbage species. Herbage Abstracts. 41, 213-221. [ Links ]
Davie, S.J., 1989. Laboratory Methods. Animal Nutrition Sub-directorate. Animal and Dairy Science Research Institute, Irene, Tvl, South Africa. [ Links ]
De Figueiredo, M., 2000. Fodder Conservation-Silage. In: Pasture Management in South Africa. Ed. Tainton, N., University of Natal Press, Pietermaritzburg, South Africa. [ Links ]
Demarquilly, C., 1973. Composition chimique, caractéristiques fermentaires, digestibilité et quantité ingérée des ensilages de fourrages: modification par rapport au fourrage vert initial. Ann. Zootech. 22, 1-35. (cited by Meeske et al., 1999). [ Links ]
Erdman, R., 1993. Silage fermentation characteristics affecting feed intake. Proc. National Silage Production Conference. (NRAES, Syracuse, New York. USA). [ Links ]
Imura, Y., Namihira, T. & Kawamoto, T., 2001. Fermentation quality of phasey bean and guinea grass silages. In: Proc. 19th Int. Grasslands Congress, Sao Paulo, Brazil, Piracicaba, SP, Brazil. Eds Jose Alberto Gomide, Wilson Roberto Mattos, Sila Carneiro da Silva. pp. 784-785. [ Links ]
Jonsson, A., 1991. Growth of Clostridium tyrobutyricum during fermentation and aerobic deterioration of grass silage. J. Sci. Food Agric. 54, 557-568. [ Links ]
Kung, L. & Shaver, R., 2001. Interpretation and use of silage fermentation analysis reports. University of Wisconsin, Madison, WI, USA. [ Links ] Focus on Forage, 3 (13), 1-5. [ Links ]
McDonald, P., 1973. The Ensilage Process. In Chemistry and Biochemistry of Herbage. Volume 3. Eds G.W. Butler, G.W. & Bailey, R.W., Academic Press, New York, USA. p. 33. [ Links ]
McDonald, P. & Henderson, A.R., 1964. Determination of water-soluble carbohydrates in grass. J. Sci. Food Agric. 15, 395-398. [ Links ]
McDonald, P., Henderson, A.R. & Heron, S.J.E., 1991. The Biochemistry of Silage. 2nd ed. Chalcombe Publications, Marlow, Bucks, UK. [ Links ]
McDonald, P., Edwards, R.A., Greenhalgh, J.F.D. & Morgan, C.A., 2002. Animal Nutrition. 6th ed. Pearson Education, Harlow, UK. [ Links ]
Meeske, R., Basson, H.M. & Cruywagen, C.W., 1999. The effect of a lactic acid bacterial inoculant with enzymes on the fermentation dynamics, intake and digestibility of Digitaria eriantha silage. Anim. Feed Sci. Technol. 81, 237-248. [ Links ]
Meeske, R., Cruywagen, C.W., Van der Merwe, G.D. & Greyling, J.F., 2000. The effect of adding a lactic acid bacterial inoculant to big round-bale oat silage on intake, milk production and milk composition of Jersey cows. S. Afr. J. Anim. Sci. 30, 80-81. [ Links ]
Oude Elferink, S.J.W.H., Driehuis, F., Gottschal, J.C. & Spoelstra, S.F., 2000. Silage fermentation processes and their manipulation. In: Silage making in the tropics with particular emphasis on smallholders. Ed. t'Mannetje, L., Proc. FAO Electronic Conference on Tropical Silage. 1 Sept. to 15 Dec. 1999. FAO Plant Production and Protection Paper 161, Food and Agricultural Organization of the United Nations, Rome, Italy. [ Links ]
Payne, M.J. & McDonald, P., 1966. The buffering constituents of herbage and silage. J. Sci. Food Agric. 17, 264. [ Links ]
Pryce, J.D., 1969. A modification of the Barker-Summerson method for the determination of lactic acid. Analyst. 94, 1151. [ Links ]
Rethman, N.F.G., 1983. Planted pastures for foggage. Type piece. Nooitgedacht Research Station, Ermelo, Republic of South Africa. [ Links ]
Samuels, M.L., 1989. Statistics for Life Sciences. Collier Macmillan Publishers, London, UK. [ Links ]
SAS/STAT, 2001. Statistical Analysis Systems User's Guide Version 8.2. SAS Institute Inc., Cary, N.C., USA. [ Links ]
Schleifer, K.H. & Ludwig, W., 1995. Phyogenetic relationships of lactic acid bacteria. In: The Genera of Lactic Acid Bacteria. Eds Wood, B.J.B. & Holzapfel, W.H., Blackie Academic & Professional, London, UK. pp. 7-18. [ Links ]
Schroeder, J.W., 2004. Silage fermentation and preservation. NDSU Extension Service. In: http://www.ext.nodak.edu/extpubs/ansci/range/as1254.pdf#search='Silage%20fermentation%20and%20preservation'. North Dakota State University Fargo, North Dakota 58105. [ Links ]
Sollenberger, L.E., Ricardo, A.R., Nussio, L.G., Chamnliss, C.G. & Kunkle, W.E., 2004. Conserved Forage. In: Warm-Season (C4) Grasses. Eds Moser, L.E., Burson, B.L. & Sollenberger, L.E., Agronomy No. 45. American Society of Agronomy Inc. Madison, Wisconsin, USA. pp 355-387. [ Links ]
Sujatha, P., Allen, V.G., Fontenot, J.P. & Jayasuriya, M.C.N., 1986. Ensiling characteristics of tropical grasses as influenced by stage of growth, additives and chopping length. J. Anim. Sci. 63, 197-207. [ Links ]
Tainton, N., 2000. Pasture Management in South Africa. University of Natal Press, Pietermaritzburg, South Africa. [ Links ]
Titterton, M. & Bareeba, F.B., 2000. Grass and legume silages in the tropics. In: Silage making in the tropics with particular emphasis on smallholders. Ed. t'Mannetje, L., Proc. FAO Electronic Conference on Tropical Silage. 1 Sept. to 15 Dec. 1999. FAO Plant Production and Protection Paper 161, Food and Agricultural Organization of the United Nations, Rome, Italy. [ Links ]
Ward, R.T., 2000. Fermentation analysis: use and interpretation. Proc. Tri-State Nutrition Conference, Fort Wayn, IN. (cited by Bethard, 2006). [ Links ]
Weissbach, F., 1996. New developments in crop conservation. Proc. 11th Int. Silage Conf. 8 to 11 September 1996, University of Wales, IGER, Plas Gogerddan, Aberystwyth, Sy 213 3EB, UK, 9 11-25. [ Links ]
Wilkins, R.J., Hutchinson, K.J., Wilson, R.F. & Harris, C.E., 1971. The voluntary intake of silage by sheep. I. Interrelationships between silage composition and intake. J. Agric. Sci. 77, 531-537. [ Links ]
Woolford, M.K., 1984. The Silage Fermentation. Microbiological Series, 14, Marcel Dekker Inc., New York and Basel. 63. [ Links ]
# Corresponding author. E-mail: willem.vanniekerk@up.ac.za


{kind=link}
{kind=link}
{kind=link}