










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.40 n.4 Pretoria Jan. 2010
Influence of supplemental aminooligosaccharides on in vitro disappearance of diets for dairy cattle and its effects on milk yield
J.A. SánchezI; J.M. Pinos-RodríguezII,#; S.S. GonzálezI; J.R. BarcenaI; J.C. GarcíaII
IGanadería, Colegio de Posgraduados, Montecillo, 56230, México
IIInstituto de Investigación de Zonas Desérticas, Universidad Autónoma de San Luis Potosí, San Luis Potosí, S.L.P. 78377, México
ABSTRACT
Two experiments were conducted to evaluate the influence of supplemental aminooligosaccharides (AOS) on the disappearance and fermentation of diets for lactating cows, and its effects on milk yield and quality. In the first study, diets containing 0 or 0.2% AOS (dry matter basis) were incubated for 24, 48 and 72 h with ruminal fluid to evaluate the in vitro dry matter disappearance (IVDMD) and fermentation characteristics. The addition of AOS improved IVDMD of diets, induced higher total bacterial and cellulolytic bacterial counts and decreased the number of bacteria that produce lactate compared to the control. At 24 h incubation, AOS increased volatile fatty acid concentrations and the molar proportion of propionate, though at 48 and 72 h incubation AOS did not affect the fermentation characteristics of the diets. In a further experiment, milk yield and milk quality were evaluated using 50 Holstein dairy cows (193 days in milk). Cows were randomly assigned to the diets tested in the in vitro study. Feed intake was not modified by AOS. Milk yield of cows fed AOS was significantly higher than that of cows not supplemented. Because AOS tended to decrease the percentage of fat in milk, fat and solid-corrected milk content were similar for the two treatments. Percentage and yield of non-fat solids, protein and lactose, and the somatic cell count in milk were not affected by AOS. The milk urea-N concentration of the cows fed AOS was lower than of those not supplemented. Supplementing AOS enhanced the disappearance and fermentation characteristics of the diet. Further studies with cows earlier in lactation are necessary to evaluate the effects of AOS on milk yield and milk components.
Keywords: Milk production, milk components, ruminal bacteria, volatile fatty acids
Introduction
Antibiotics and hormones have been used in dairy cows in order to improve productivity and to protect them against pathogens. However, new alternatives and better solutions must be found since consumers and several government and non-government organizations demand non-chemical and consumer friendly products. A wide variety of prebiotics has received considerable attention in animal nutrition. Gibson & Roberfroid (1995) defined a prebiotic as a nondigestible food ingredient that beneficially affects the host by selectively stimulating the growth and activity of one or a limited number of bacteria in the gut, and thus improving host health. In animals, the most common prebiotics studied, are fructooligosaccharides (FOS), mannanoligosaccharides (MOS), and galactooligosaccharides (GOS). The beneficial effects of these prebiotics on feed efficiency and immune stimulation of these additives have been demonstrated mainly in dogs (Swanson et al., 2002), pigs (Burkey et al., 2004), poultry (Bozkurt et al., 2009a; b) and young calves (Heinrichs et al., 2003). It is possible that, due to the fact that some prebiotics might not be degraded in the gastrointestinal tract, these functional carbohydrates could reach the large intestine intact where they could have beneficial effects.
In ruminants there is evidence that prebiotics increase the population of ciliated protozoa in the rumen (Cendrowska et al., 2006), decrease ammonia nitrogen (NH3-N) concentration (Shibata, 1985; Biggs & Hancock, 1998), increase volatile fatty acid (VFA) and microbial protein concentrations, and stabilize the rumen pH (Li et al., 2007). However, other studies do not support these results, probably due to the fact that prebiotics are rapidly fermented by ruminal microorganisms. In fact, FOS induced lameness in dairy heifers since heifers developed classic signs of acute ruminal and systemic acidosis after an oligofructose overload (Danscher et al., 2009; Niss et al., 2009). Also, FOS increased nitrogen (N) excretion and decreased protein digestibility, and the milk fat and protein content in dairy cows (Gressley & Armentano, 2007). A new natural aminooligosaccharide (AOS) has been developed for animal feeding, but has not been evaluated in dairy cows. In this study the addition of AOS to dairy cow diets was investigated by measuring its influence on the in vitro dry matter disappearance (IVDMD) and in vitro fermentation of the diet, and this was followed by a study on its effect on milk production and of lactating dairy cows and the composition of the milk.
Materials and Methods
The in vitro study was performed in the Laboratory for Microbiology of Colegio de Postgraduados, Campus Montecillo in México. The trial with lactating dairy cows was carried out on a dairy farm in the State of Guanajuato (Ojo de Agua Farm), located at 1819 m above sea level, and an average annual temperature and rainfall of 19.5 ºC and 360 mm, respectively. According to the manufacturer, the aminooligosaccharide product (aminoglucolite, Química Foliar, S.A. de C.V., Naucalpan, Edo. de México) contains 75% aminooligosaccharides and amino acids at concentrations of (/100 g DM): 2.11 g histidine, 0.6 g lysine, 0.34 g glutamine, 0.29 g isoleucine, 0.25 g leucine, 0.25 g methionine, 0.25 g asparagine, 0.21 g serine, 0.15 g alanine, 0.13 g glycine, 0.13 g arginine, 0.12 g phenylalanine, 0.12 g proline, 0.11 g tyrosine and 0.07 g threonine.
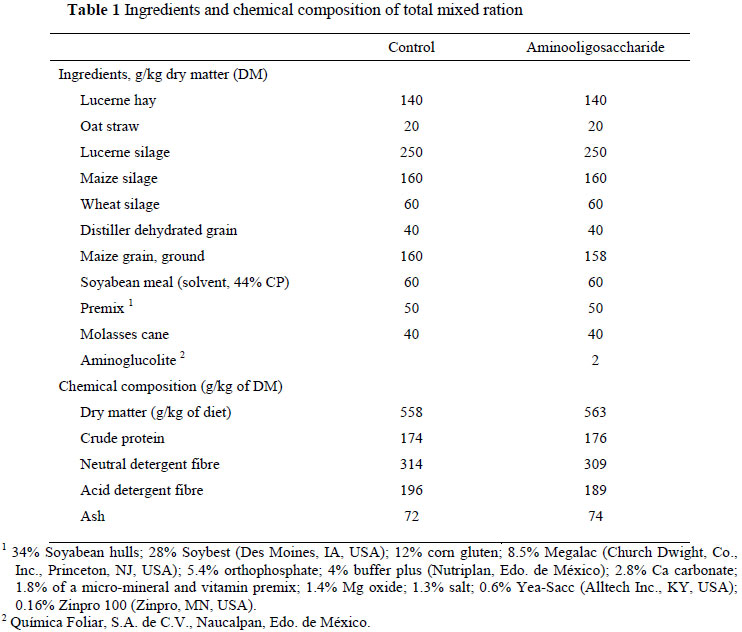
Ruminal fluid was collected from two Holstein cows (650 kg body weight) fitted with ruminal cannulae. The cows had free access to water and received a complete diet (50% forage and 50% feed grain). Samples (200 g) of the experimental diets with 0 (control) or 0.2% AOS (Table 1) were ground through a 1 mm screen (Tomas Willey), oven-dried at 105 ºC for 24 h, and sterilized (121 ºC for 20 min). The concentrations of crude protein (CP; micro-Kjeldahl method), ash (AOAC, 1995), and neutral detergent fibre (NDF) and acid detergent fibre (ADF) (Van Soest et al., 1991) in the diets were determined (Table 1). The experimental design constituted two treatments: A control (without AOS) and a diet containing 0.02% AOS (dry matter basis).
Two hundred milliliters of ruminal content from each cow were obtained 4 h after morning feeding and squeezed through two layers of cheesecloth into a 500 mL Erlenmeyer flask with an O2-free CO2 headspace. The flask was then placed in a 39 ºC waterbath and left undisturbed for 15 min, which allowed the feed particles to rise to the top of the flask. Particle-free fluid (1 mL/tube) from the flask was anaerobically transferred to glass tubes (18 x 150 mm) with butyl rubber stoppers. The tubes contained 9 mL of an anaerobic buffer (9 mL/tube; Cobos et al., 2002) and 100 mg of the diet (as DM). Twenty four roll tubes with the diet (100 mg) and ruminal fluid plus buffer (9 mL) were incubated for 24, 48 and 72 h at 39 ºC. After the incubation, a 1 mL sample was collected, and serial 10-fold dilutions in an anaerobic mineral solution (Cobos et al., 2002) were prepared. The Hungate (1969) anaerobic technique was used to prepare the media and to cultivate cellulolytic bacteria in the roll-tubes (triplicate) with the complete medium described by Cobos et al. (2002). Dilutions of 10VI to 109 were used to cultivate total cellulolytic bacteria. One milliliter of diluted sample from each tube was used to inoculate 5 mL of culture media in the roll tubes. The existence of cellulolytic bacteria was assumed by the degradation of a filter paper strip (Whatman 541). For bacteria that utilize lactate, a Man-Rogosa-Sharp (MRS) culture media was used. Counts of ruminal bacteria were carried out microscopically (Olympus BX51 100x) in a counting chamber (Petroff-Hausser chamber, Partnership SQMM, Horsham, PA, USA). Thus, 1 mL of formalin (9.2%) and 1 mL of incubated fluid were mixed in roll tubes (13 x 100 mm). Using a Pasteur pipette, one drop was dropped into a chamber. Counts of 10 squares of 0.005 x 0.005 cm each were averaged. Some ruminal fluid dilutions were developed using distilled water. The equation used to quantify the total bacteria was as follow:
Bacteria/mL =  (FD) (2 x 107), where = medium and, FD = dilution factor.
(FD) (2 x 107), where = medium and, FD = dilution factor.
Roll tubes with a butyl rubber stopper were uncapped, and pH was determined immediately in the culture fluid with a portable pH meter. For analysis of volatile fatty acids (VFA), 0.2 mL of 25% metaphosphoric acid (wt/vol) was added to 1 mL of fermentation fluid, and stored at -30 ºC until analyzed. One milliliter of thawed fermentation fluid was centrifuged (10 000 g for 10 min at 4 ºC), and VFA's were analyzed by gas chromatography (Erwin et al., 1961). One milliliter of thawed sample was centrifuged (27 000 g for 20 min at 4 ºC) and NH3-N (McCullough, 1967) concentrations were determined using a UV-VIS Spectrophotometer.
The IVDMD of the diets at 24, 48 and 72 h was estimated from the dry matter (DM) remaining in the roll tube after incubation. The non-degradable residue was collected on a pre-weighed oven-dried Whatman 541 filter by vacuum filtration. The filter and undigested residues were oven-dried at 105 ºC for 24 h to remove the moisture and then weighed. The IVDMD was calculated as original dry sample weight minus dry residue weight divided by the original sample weight.
For the fermentation study, i.e. microorganism count and IVDMD, 24 roll tubes incubated for 24, 48 and 72 h were assigned to the two treatments (control and AOS). Thus, four tubes were used for each treatment at each incubation time. An All ANOVA analysis using PROC GLM of SAS V8 (The SAS Institute, Cary, N.C., USA) was used in the statistical analysis.
In the study on lactating dairy cows, 50 Holstein cows in late lactation were randomly assigned to the two experimental diets: 0 AOS (control) or 0.2% AOS (Table 1). At the onset of the study the cows in second lactation were on average 193 ±18 days in milk (DIM), produced 25.3 ± 2.1 kg milk/d and weighed 642 ± 39.9 kg. Cows were allocated in one pen per treatment and were fed once a day. At the beginning of the trial the average milk yield was 22.8 ± 3.8 kg/d for the control and 23.1 ± 4.1 for the AOS treatment. To ensure a correct AOS level (40 g/kg), the AOS product was previously added to a mineral premix (Table 1). The trial lasted 69 days. The cows were milked twice daily (8:00 and 16:00) and yields were recorded, electronically. Feed offered and orts were measured daily. Orts were sampled from each pen once a week and analyzed for DM (105 ºC for 24 h) to calculate dry matter intake (DMI). Since the cows were group-fed DMI between treatments was not analyzed statistically. The diets were analysed for CP, ash, NDF and ADF, using the techniques as reported above. Milk (a.m. and p.m.) was sampled every week from each cow and analyzed for concentration of milk fat, protein, lactose, non-fat solids, milk urea nitrogen (MUN) (Infrared Analyzer, MilkoScan 133B, FOSS Analytical A/S, Hillerød, Denmark) and somatic cell count (SCC; Fossomatic 5000, FOSS Analytical A/S, Hillerød, Denmark) by Alpura S.A. de C.V., Querétaro, México.
Milk production by cow was averaged per week. Fat-corrected milk (FCM) and non-fat solids (SCM) were calculated as follows:
3.5% FCM = (0.4324 * lb milk) + (16216 * lbs of milk fat);
SCM = milk lb * ((12.24 * fat %) + (7.10 * protein %) + (6.35 * lactose %) - 0.0345)).
The FCM and SCM (lbs basis) were converted to a kg basis. To approximate the normal distribution, a natural logarithmic transformation of SCC was performed. For presentation purposes, logarithmic transformation was converted to SCC after analysis. Overall milk production from a week prior to the onset of the study was included as a covariate (P >0.05) in the model. Weekly means for milk production and milk components were analyzed using the Mixed Procedure of SAS V8 (The SAS Institute, Cary, N.C., USA). The model included the effects of treatments (1 df) and time (weekly, 8 df). A cow was considered as a random component in the model. The repeated measure was analyzed within cow. The covariate structure that resulted in the lowest Akaike's information criterion was the first-order autoregression (The SAS Institute, Cary, N.C., USA).
Results and Discussion
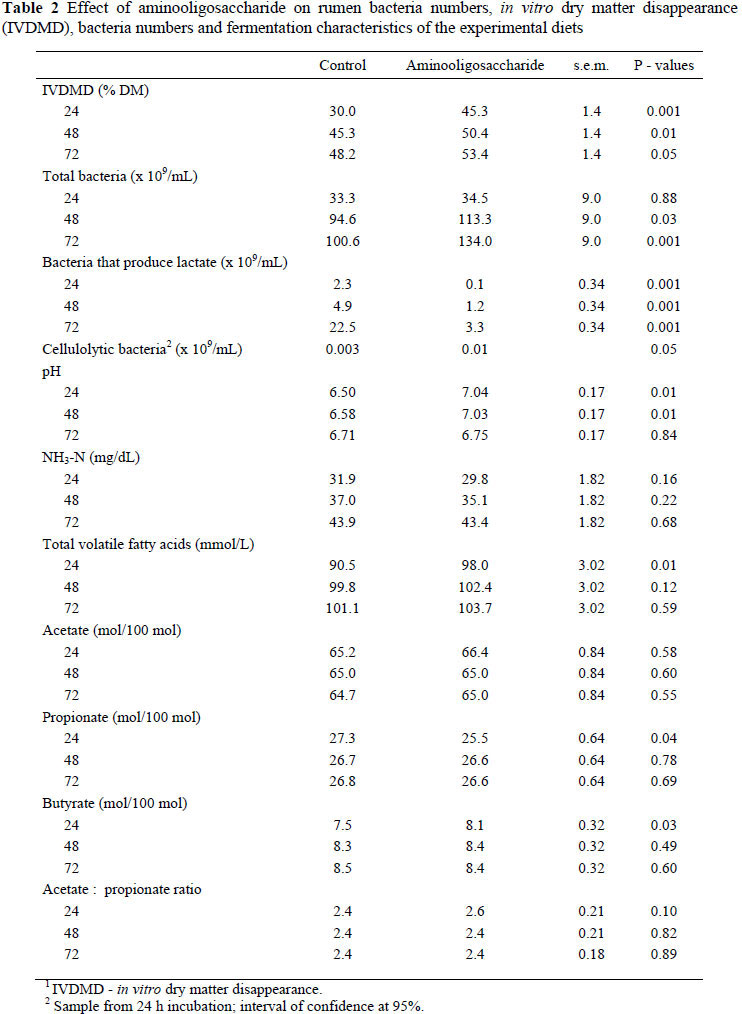
The chemical composition of experimental diets was similar (Table 1). During all three incubation periods the IVDMD of the diet containing AOS was higher (P <0.05) than that of the control diet. The AOS induced a lower number of bacteria that produce lactate, but higher numbers of cellulolytic bacteria than the control. The total number of bacteria was also higher in the AOS treatment than in the control (Table 2). The improved IVDMD due to OAS could be attributed to the increase in desirable bacteria such as cellulolytic bacteria, and the inhibitory effects on undesirable bacteria such as those that produce lactate. Although the mechanism(s) by which AOS stimulates or inhibits bacterial growth is uncertain, scientific evidence suggests that these prebiotics reduce the growth of pathogenic bacteria such as Enterobacterium and Clostridium, and increase the growth of the beneficial Bifidobacterium in the gut of monogastric animals (Hussein et al., 1999; Buddington et al., 2002).
After 24 and 48 h of incubation, pH values were higher (P <0.01) in the OAS treatment than in the control. At 24 h incubation, AOS also increased total VFA concentration and the molar proportion of butyrate, and reduced the molar proportion of propionate as compared to the control. Similar results were found under in vitro conditions by Ling et al. (2007) and Öztürk (2009) who indicated that functional oligosaccharides increase total VFA under in vitro conditions. The concentrations of NH3-N, the molar proportion of acetate and the acetate : propionate ratio were not modified by OAS at any time of incubation. Also, at 24 and 48 h incubation, OAS did not affect total VFA concentration or molar proportions of propionate and butyrate. The increased pH induced by OAS could be a result of the inhibition of the number of lactate producing bacteria. We expected that OAS would decrease to NH3-N concentrations, as observed for MOS and FOS (Ling et al., 2007; Öztürk, 2009), but our results did not confirm that.
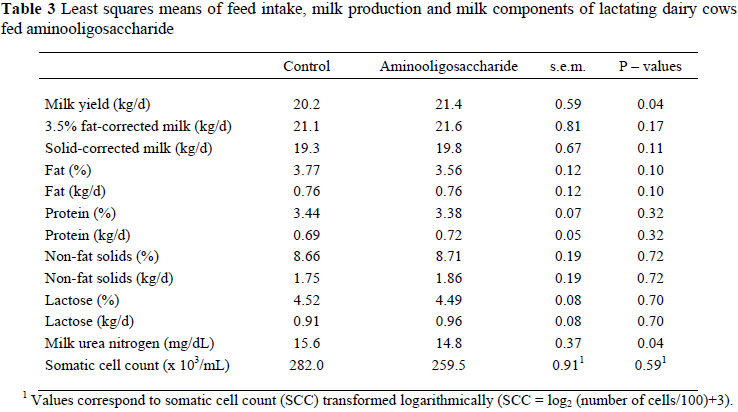
In the study with the lactating cows no interaction was recorded between time and treatments. Therefore, only overall production responses and milk composition are presented in Table 3. Daily DMI for the group receiving AOS (20.7 kg DM) was very similar to that of the control (21.3 kg DM). Cows receiving OAS produced more (P <0.05) milk than cows in the control. Since OAS tended (P = 0.10) to decrease the percentage of fat in milk, the FCM and SCM levels in the milk were similar for the two treatments. The decreased milk fat percentage caused by AOS could be due to a decrease in ruminal pH, since Mwenya et al. (2005) reported that supplementation of functional oligosaccharides to Holstein cows may reduce the ruminal pH. Furthermore, Gressley & Armentano (2007) observed that abomasal infusions of FOS as inulin seemed to elicit a decrease in milk fat apparently due to a decrease in the secretion of 18-carbon fatty acids. Milk components, expressed as % or kg/d were not affected by AOS. The MUN concentration of cows fed AOS was lower than that of cows not supplemented. Reductions in MUN attributable to functional oligosaccharides were confirmed by Gressley & Armentano (2007). The reduction of MUN by AOS could be due to a better efficiency of N utilization (Nousiainen et al., 2004) or to a reduced digestibility of the diet (Gressley & Armentano, 2007). The SCC in the milk of cows in both treatments was similar.
Conclusions
Aminooligosaccharides could be of great value in increasing microbial growth and creating and maintaining a desired, stable microflora in the rumen. Because DMI in late lactation cows versus early lactations differs substantially, additional work is needed to clarify the differences in response to aminooligosaccharide addition in the diets of early lactation cows.
References
AOAC, 1995. Official Methods of Analysis. 16th ed. Association of Official Analytical Chemists, Arlington, V.A., USA. [ Links ]
Biggs, D.R. & Hancock, K.R., 1998. In vitro digestion of bacterial and plant fructans and effects on ammonia accumulation in cow and sheep rumen fluids. J. Gen. Appl. Microbiol. 44, 167-171. [ Links ]
Bozkurt, M., Küçükyılmaz, K., Çatl, A.U. & Çınar, M., 2009a. Effect of dietary mannan oligosaccharide with or without oregano essential oil and hop extract supplementation on the performance and slaughter characteristics of male broilers. S. Afr. J. Anim. Sci. 39, 223-232. [ Links ]
Bozkurt, M., Küçükyılmaz, K., Çatl, A.U. & Çınar, M., 2009b. The effect of single or combined dietary supplementation of prebiotics, organic acid and probiotics on performance and slaughter characteristics of broilers. S. Afr. J. Anim. Sci. 39, 197-205. [ Links ]
Buddington, K.K., Donahoo, J.B. & Buddington, R.K., 2002. Dietary oligofructose and inulin protect mice from enteric and systemic pathogens and tumor inducers. J. Nutr. 132, 472-477. [ Links ]
Burkey, T.E., Dritz, S.S., Nietfeld, J.C., Johnson, B.J. & Minton, J.E., 2004. Effect of dietary mannanoligosaccharide and sodium chlorate on the growth performance, acute-phase response, and bacterial shedding of weaned pigs challenged with Salmonella enterica serotype Typhimurium. J. Anim. Sci. 82, 397-404. [ Links ]
Cendrowska, M.C., Kasprowicz, A. & Michalowski, T., 2006. Preliminary assessment of the capability of the rumen bacterium, Butyrivibrio fibrisolvens, to utilize fructose polymers for growth. J. Anim. Feed Sci. 15, 11-14. [ Links ]
Cobos, M.A., García, L.E., González, S.S., Bárcena, J.R., Hernández, D.S. & Pérez-Sato, M., 2002. The effect of shrimp shell waste on ruminal bacteria and performance of lambs. Anim. Feed Sci. Technol. 95, 179-187. [ Links ]
Danscher, A.M., Enemark, J.M.D., Telezhenko, E., Capion, N., Ekstrøm, C.T. & Thoefner, M.D., 2009. Oligofructose overload induces lameness in cattle. J. Dairy Sci. 92, 607-616. [ Links ]
Erwin, E.S., Marco, G.J. & Emery, E., 1961. Volatile fatty acid analysis of blood and rumen fluid by gas chromatography. J. Dairy Sci. 44, 1768-1771. [ Links ]
Gibson, G.R. & Roberfroid, M.B., 1995. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 125, 1401-1412. [ Links ]
Gressley, T.F. & Armentano, L.E., 2007. Effects of low rumen-degradable protein or abomasal fructan infusion on diet digestibility and urinary nitrogen excretion in lactating dairy cows. J. Dairy Sci. 90, 1340-1353. [ Links ]
Heinrichs, A.J., Jones, C.M. & Heinrichs, B.S., 2003. Effects of mannan oligosaccharide or antibiotics in neonatal diets on health and growth of dairy calves. J. Dairy Sci. 86, 4064-4069. [ Links ]
Hungate, R.E., 1969. A roll tube method for cultivation of strict anaerobes. In: Methods in Microbiology 3. Eds Norris, J.R. & Ribbons, D.W., Academy Press, New York, USA. pp. 117-132. [ Links ]
Hussein, S.H., Flickinger, E.A. & Fahey, G.C., 1999. Pet food applications of inulin and oligofructose. J. Nutr. 129, 1454S-1456S. [ Links ]
Li, G.H., Liu, B., Qu, M.R., Zhang, X.F., Liu, G.B., Gao, M. & Zhang, A.Z., 2007. Effects of soybean oligosaccharides infusion in different parts of the gastrointestinal tract on several immune indices in sheep. Chin. J. Anim. Sci. 19, 1-7. [ Links ]
Ling, B.M., Qiu, M.R., Lu, D.X. & Liu, G.B., 2007. Effects of functional oligosaccharides on the rumen fermentation of the growing sheep in vitro. Chin. J. Anim. Sci. 19, 129-134. [ Links ]
McCullough, H., 1967. The determination of ammonia in whole blood by direct colorimetric method. Clin. Chem. 17, 297-304. [ Links ]
Mwenya, B., Santoso, B., Sar, C., Pen, B., Morikawa, R., Takaura, K., Umetsu, K., Kimura, K. & Takahashi, J., 2005. Effects of yeast culture and galacto-oligosaccharides on ruminal fermentation in Holstein cows. J. Dairy Sci. 88, 1404-1412. [ Links ]
Niss, D.B., Herskin, M.S., Danscher, A.M. & Thoefner, M.B., 2009. Short communication: Rising and lying behavior of heifers before and after alimentary oligofructose overload. J. Dairy Sci. 92, 617-620. [ Links ]
Nousiainen, K., Shingfield, J. & Huhtanen, P., 2004. Evaluation of milk urea nitrogen as a diagnostic of protein feeding. J. Dairy Sci. 87, 386-398. [ Links ]
Öztürk, H., 2009. Effects of chicory inulin on ruminal fermentation in vitro. Ankara Üniv. Vet. Fak. Derg. 56, 171-175. [ Links ]
Shibata, T.H., 1985. Studies on the utilization of pasture herbage nitrogen by rumen microorganisms 2. Effects of carbohydrate and lipid material on the microbial utilization of rumen ammonia in vitro. Grassl. Sci. 31, 322-331. [ Links ]
Swanson, K.S., Grieshop, C.M., Flickinger, E.A., Bauer, L.L., Healy, H-P., Dawson, K.A., Merchen, N.R. & Fahey, G.C., 2002. Supplemental fructooligosaccharides and mannanoligosaccharides influence immune function, ileal and total tract nutrient digestibilities, microbial populations and concentrations of protein catabolites in the large bowel of dogs. J. Nutr. 132, 980-989. [ Links ]
Van Soest, P.J., Robertson, J.B. & Lew, B.A., 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 77, 3583-3597. [ Links ]
# Corresponding author. E-mail: jpinos@uaslp.mx


{kind=link}
{kind=link}
{kind=link}