










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.40 n.3 Pretoria Jan. 2010
Chemical characteristics of red hartebeest (Alcelaphus buselaphus caama) meat
L.C. HoffmanI,#; K. SmitI,II; N. MullerII
IDepartment of Animal Sciences, Stellenbosch University, Private Bag X1, Matieland, Stellenbosch 7602, South Africa
IIUniversity of Stellenbosch, Department of Food Science, Private Bag X1, Matieland 7602, South Africa
ABSTRACT
The aim of this study was to determine the effect of region (Qua-Qua, Maria Moroka, Sandveld and Tussen die Riviere) and gender on carcass characteristics and chemical composition of meat from the red hartebeest. The parameters measured on 29 red hartebeest included body weight, carcass weight, dressing percentage, proximate composition, fatty acid, cholesterol, amino acid and mineral content. The average carcass weight of males (79.3 kg) was significantly higher than that of females (56.0 kg); however, there was no difference in dressing percentages. The animals obtained from the Qua-Qua region had the highest lipid content (1.3 g/100 g meat sample) compared to hartebeest sampled in other regions. Differences in concentrations of individual fatty acids, amino acids and minerals were minor and of no practical value. The ratio of polyunsaturated to saturated fatty acids (0.78) is above the recommended 0.7 and therefore has positive implications for human health. These nutrient values determined in meat from red hartebeest could be incorporated into human food composition tables. These results will also be of value to the South African game meat exporters who may wish to market their products in terms of their nutritional value in order to compete with meat from domestic livestock or international venison products. In such cases, it is required by law (both locally and internationally) to provide adequate nutritional labelling.
Keywords: Game meat, venison, fatty acids, cholesterol, amino acids, minerals, sustainable utilisation
Introduction
Game meat is usually sold per animal or per kilogram (Hoffman & Wiklund, 2006). It is therefore important to know which animals will give the highest dressing percentage or yield of meat (Issanchou, 1996). Von La Chevallerie (1970) noted that the live weight of an animal alone does not reflect its meat production potential. However, carcass weight can give a good indication of an animal's meat production potential if sufficient data on the carcass composition is available. A number of researchers have found that wild ungulates often produce dressing percentages similar to, if not higher than many domestic livestock species which results from the higher proportion of muscular tissue present in the former carcasses (Ledger, Sachs & Smith, 1967; Skinner, 1984). The ability of wild ungulates to compete with domestic livestock, however, is most notable in their ability to reach mature weights at a younger age under harsher extensive farming conditions (Skinner, 1984). Furthermore, Berry (1986) noted that game meat production was the most profitable strategy for game farms in terms of return per kilogram of biomass when compared to live sales, trophy hunting and recreational hunting.
According to Onyango et al. (1998) the decrease in the per capita supply of high quality protein and the increase in the world's population have intensified the need to search for other sources of protein. Although there is an increase in the demand for game meat, there is a tendency for the meat to be sold under the generic name of "game meat" or "venison", with no specific indication of species. A possible reason for generic marketing is the fact that nutritional information on the meat of most game species is unavailable (Hoffman & Wiklund, 2006).
South African game meat is often described as an organic product, as no fertilizers or growth stimulators are used in the production system (Pauw, 1993). The negative health image that many consumers associate with red meat is due to its predominantly saturated animal fat, which is associated with health risks (Schönfeldt, 1993). The current dietary recommendations emphasize an increased intake of polyunsaturated fatty acids and a decreased intake of saturated fatty acids. Meat from free-ranging game such as eland is rich in polyunsaturated fatty acids (Lawrie, 1979), and meat from pasture-fed animals such as reindeer has a high percentage of a-linolenic acid (C18:3n-3) (Wiklund et al., 2001). An increased consumption of meat from free-ranging or pasture-fed animals could contribute to an intake of more polyunsaturated fatty acids (Simopoulos, 2002).
Red hartebeest (Alcelaphus buselaphus caama) are found in regions of the North Western Cape (South Africa), Botswana, Namibia and in Zimbabwe (Zaburnis & Cross, 1974). This species has also been introduced to reserves in KwaZulu-Natal and other national parks in South Africa. Red hartebeest occupy ridge and kopje summits and are known to maintain their territories for long periods. In 1994 the estimated number of red hartebeest in the Limpopo Province was 15 000 (Van der Waal & Dekker, 2000). This indicates that there is an enormous potential for red hartebeest meat production.
The purpose of this study was to determine the effect of region and gender on carcass characteristics and chemical composition (proximate, cholesterol, fatty acids, amino acid and mineral content) of red hartebeest meat.
Materials and Methods
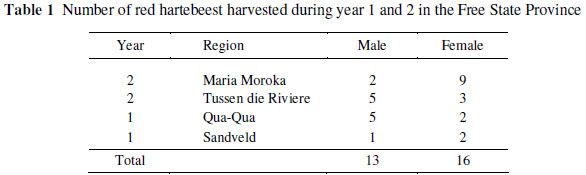
Red hartebeest, representing both genders, were obtained during two years (year 1 and year 2) from four nature reserves in the Free State Province, South Africa (Table 1). Due to unforeseen circumstances it was not possible to obtain the carcass measurements from Sandveld and some of the animals from Tussen die Riviere. All the animals were obtained between May to September (autumn to spring). The animals were selected randomly and either shot in the head or neck with .274 or .270 calibre rifles during organized game cropping operations. No trophy animals (identified by their horn size) were shot. All the animals were exsanguinated in the field as soon as possible after shooting. In year 1 red hartebeest were shot during the daytime and during the night in year 2 although this is not expected to interact with the regional effect since time of shooting may affect the ante-mortem stress experienced by the animals but not any of the variables measured in this investigation. Since all animals were obtained during the same season, the year in which they were obtained is also not expected to interact with region.
After bleeding, the carcasses were weighed for determination of "live weight" whereafter the carcasses were dressed (removal of head, feet, all intestines as well as all internal organs including the kidneys) and weighed for determination of dressed weight. The dressout percentage was calculated as a percentage of the dressed weight divided by the live weight. For chemical analyses, the m. longissimus dorsi of each animal was cut and removed from between the 6th rib and anterior to the 5th lumbar vertebrae. This game species does not have any subcutaneous fat and thus only the visible collagen tissue was removed from the muscle prior to further processing. Proximate analysis was conducted on minced (three times through 2 mm sieve) samples of all the animals. The moisture, protein and ash content (g/100 g meat) were determined according to standard techniques of the AOAC (1997), and the lipid by means of solvent (chloroform/methanol) extraction (Lee et al., 1996).
The fatty acid, cholesterol, amino acid and mineral concentration of samples from male and female animals from Maria Moroko and Tussen die Riviere were determined as described by Koep et al. (2007) and Mostert & Hoffman (2007). Fatty acid methyl esters were analysed by gas chromatography (GC) (Varian Model 3300 equipped with flame ionisation detection) using 60 m BPX70 Capillary columns of 0.25 mm internal diameter (SGE, Australia). Gas flow rates were: hydrogen, 25 mL/min; and hydrogen carrier gas 2-4 mL/min. Temperature programming was linear at 3 ºC/min, with an initial temperature of 150 ºC, a final temperature of 220 ºC, an injector temperature of 240 ºC and a detector temperature of 250 ºC. The FAME were identified by comparison of the retention times to those of a standard FAME mixture (Nu-Chek-Prep Inc., Elysian, Minnesota). From the same lipid extraction used for fatty acid determination, a sub-sample was used for cholesterol determination by GC (Varian Model 3700, equipped with flame ionization detection). The amino acid composition was determined using a modification of the method of Bidlingmeyer et al. (1984) on a defatted, dried meat sample using a Waters high performance liquid chromatography system (1525 HPLC with a binary gradient delivery, 717 auto-sampler and Injector, 1500 column heater, 2487 dual wavelength UV detector) and a Breeze data workstation (Waters, Millford, MA, USA). Mineral concentrations were measured on a Varian (Victoria, Australia) Liberty Series II sequential inductively coupled plasma atomic emission spectrophotometer.
A two factor factorial experiment was performed in a completely randomized design with an unequal number of random replications. The factors were the four regions (Maria Moroka, Tussen die Riviere, Qua-Qua and Sandveld) and the two sexes. A single carcass was considered to be an experimental unit and the data subjected to an analysis of variance using SAS version 8.2 (SAS, 1999). Student's t-Least Significant Differences were calculated at the 5% confidence level to compare treatment means (Ott, 1998).
Results and Discussion
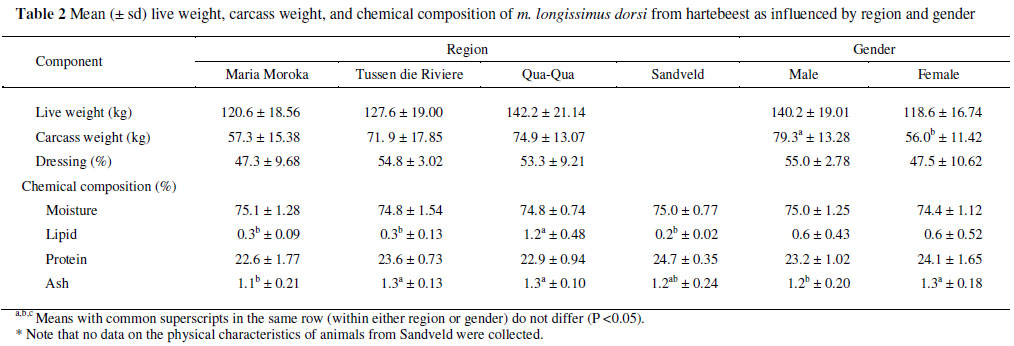
Carcass weight was higher (P <0.05) in males than in females, but no differences (P >0.05) were found in live weight or dressing percentage (Table 2). In comparison with females, the male animals usually have a higher weight in most game species, because of their thicker necks and heavier forequarters (Von La Chevallerie, 1970). The reason for the male animals in this investigation being lighter than expected was due to the fact that animals that were not trophy animals (normally heavier) were selected for culling as the four regions sell trophy animals for higher revenue. The dressing percentage of red hartebeest from this study is comparable with the 50% reported for blesbok (Damaliscus pergagus phillipsi) (Van Zyl & Ferreira, 2001) and 53% duiker (Sylvicapra grimmia) (Ferreira & Hoffman, 2001), but lower than the 58% found for impala (Aepyceros melampus) (Van Zyl et al., 1969; Hoffman, 2000; Hoffman et al., 2005a) and 59% for kudu (Tragelaphus strepsiceros) (Mostert & Hoffman, 2007). Ledger et al. (1967) reported the carcass weight for red hartebeest males as 81.5 kg and 73.2 kg for females. This difference compared to present results could be due to the differences in the methodology of determining this parameter, e.g. which organs (kidneys, etc.) were included in the carcass weight or, as already explained, due to the fact that non trophy animals were shot. The phenomenon that neither the live weight nor the dressing percentage differed between the genders, but the carcass weight did, may be explained by the high variations (Table 2) noted in these measurements, especially for the dressing percentage. Region had no effect on any of these measurements (Table 2).
Although differences (P <0.05) were found for lipid content between the regions, the mean values are still low compared to other farmed animal species. The regional differences found for the lipid value could possibly be ascribed to the high number of animals cropped in a specific region. It is also noteworthy that the muscle lipid content of the animals from Qua-Qua was not only higher than that from the other regions but these animals also tended to exhibit heavier carcass weights. It would be interesting to see whether this trend (positive correlation between carcasses weight and muscle lipid content) holds true if a larger sample of animals was investigated. In this study the lipid content in meat was similar in males and females. Although not different between genders in kudu (Mostert & Hoffman, 2007), the lipid content of meat was found to be higher in females than males for impala (Hoffman, 2000; Van Zyl & Ferreira, 2004; Hoffman et al., 2005a), springbok (Antidorcas marsupialis) (Van Zyl & Ferreira, 2004; Hoffman et al., 2007a) and blesbok (Van Zyl & Ferreira, 2004). Significant differences were found for ash content between region and gender. These differences are, however, difficult to explain since no other proximate differences were noted except for differences in lipid content between regions and also because literature on the proximate composition of African ungulates is lacking. If the sample size had been larger, differences in ash content may have become insignificant. The protein content of red hartebeest meat is similar to that reported for the m. longissimus dorsi of the impala (23.8 g/100 g; Hoffman, 2000), common duiker (25.7 g/100 g; Hoffman & Ferreira, 2004) and kudu (24.0 g/100 g; Mostert & Hoffman, 2007).
Table 3 illustrates that palmitic acid (C16:0) and stearic (C18:0) acid are the major constituents of the saturated fatty acid component and oleic acid (C18:1n-9) the major monounsaturated fatty acid in meat from the hartebeest. This is in accordance to the fatty acid profile of game meat and other red meat types (Aidoo & Haworth, 1995). However, in the case of beef (Enser et al., 1998), palmitic acid (29%) and oleic acid (42%) are both higher than that of the meat of red hartebeest. Fatty acid composition of meat is currently receiving attention because of its implications for human health. The aim is to bring the ratio of polyunsaturated to saturated fatty acids of meat closer to the recommended value of above 0.7 as well as the n-6:n-3 ratio to less than 5.0 (Sanudo et al., 2000; Raes et al., 2004). Red hartebeest had a mean ratio of polyunsaturated to saturated fatty acids of 0.78 whilst the n-6:n-3 ratio was 2.71. This n-6:n-3 ratio compared favourably with that of the springbok m. longissimus dorsi which varied from 3.02 to 3.35, with an average ratio of 3.2 (Hoffman et al., 2007b) and compares well to values of 0.58 and 1.81 reported in the meat of a wide range of African game ungulates (Hoffman & Wiklund, 2006).
No differences were found for cholesterol content within the subgroups (Table 3). In this study the highest cholesterol value for red hartebeest was 59.9 mg/100 meat sample, compared to 89.6 mg/100 g found for South African beef (Schönfeldt, 1993) and 54 5 -59.3 mg/100 g muscle reported for springbok (Hoffman et al., 2007b). The lower cholesterol content of red hartebeest can be considered an important quality attribute, and should be emphasised in the marketing of the meat of this species as game meat.
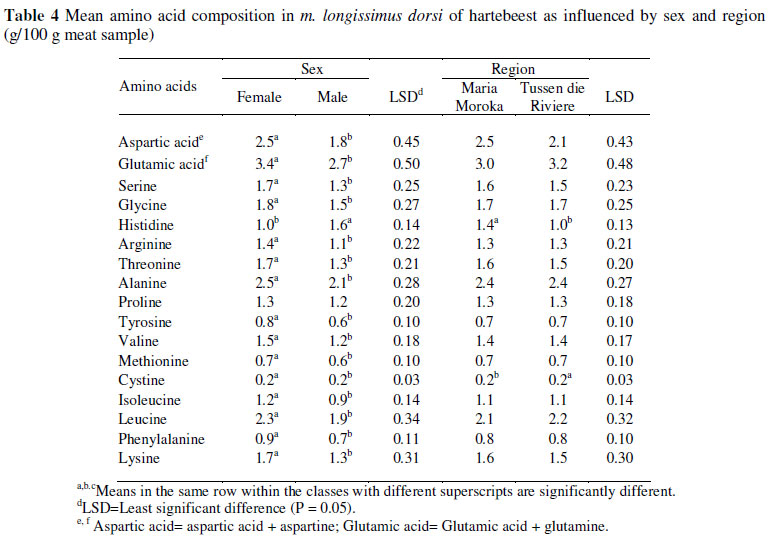
Other than histidine the meat from the female animals measured higher (P S0.05) values for all the amino acids when compared to the male animals (Table 4). It is also noteworthy that females had higher (P >0.05) total protein values than males (Table 2). Within region, differences (P S0.05) were found for histidine and cystine. The animals from MM had a higher histidine value (1.37 mg/100 g muscle) when compared to the animals from TDR (1.0 mg/100 g muscle). The red hartebeest from TDR had a higher isoleucine (0.2 mg/100 g muscle) value than the animals from MM (0.2 mg/100 g muscle). As for springbok (Hoffman et al., 2007a) and impala (Hoffman et al., 2005b), the two amino acids presented at highest levels were glutamic and aspartic acid, with leucine the most abundant essential amino acid.
No differences (P >0.05) for minerals measured (Table 5) were found between regions and gender. This phenomenon is most probably attributed to the high variation in the mineral content of the muscle. This high variation also makes it difficult to calculate daily human dietary intake for game meat. The iron values reported by Lawrie (1979) and Lushbough & Schweigert (1960) for beef steak (4.3 mg/100 g) and beef loin (2.5 mg/100 g) were much lower than that noted here. The higher iron content of game meat could be attributed to a higher myoglobin content as wild ungulates are known to have more myoglobin than domesticated and farmed animals (Hoffman et al., 2005b).

Conclusions
From the results of this study it is clear that region and gender had little effect on the proximate composition, fatty acid, cholesterol, and mineral composition of meat from hartebeest. The differences found within and between the main effects are small (and confounded by high variations) and need not be taken into account for e.g. labelling. Overall, red hartebeest meat has a favourable fatty acid composition and is thus a healthy alternative meat source as well as being low in fat. The chemical composition of red hartebeest meat reported in the current study can be used for human food composition tables and to market game meat in terms of species specific products.
Acknowledgments
The help of Frans Marais, the culling team and the Free State Department of Tourism and Conservation and the staff of the four nature reserves, is much appreciated.
References
AOAC., 1997. Official Methods of Analysis. 16th ed. Association of Official Analytical Chemists Inc. Arlington, Virginia, USA. [ Links ]
Berry, M.P.S., 1986. A comparison of different wildlife production enterprises in the Northern Cape Province, South Africa. S. Afr. J. Wildl. Res. 16, 124-128. [ Links ]
Bidlingmeyere, B., Cohen, S.A. & Tarvin, T.L., 1984. Rapid analysis of amino acids using pre-column derivatisation. J. Chromat. 336, 93-104. [ Links ]
Enser, M., Hallett, K.G., Hewett, B., Fursey, G.A.J., Wood, J.D. & Harrington, G., 1998. Fatty acid content and composition of UK beef and calf muscle in relation to production system and implications for human nutrition. Meat Sci. 49, 329-341. [ Links ]
Hoffman, L.C., 2000. The yield and carcass chemical composition of impala (Aepyceros melampus), a southern African antelope species. J. Sci. Food Agric. 80, 752-756. [ Links ]
Hoffman L.C. & Ferreira, A.V., 2004. Chemical composition of two muscles of the common duiker (Sylvicapra grimmia). J. Sci. Food Agric. 84, 1541-1544. [ Links ]
Hoffman, L.C. & Wiklund, E., 2006. Game and venison - meat for the modern consumer. Meat Sci. 74, 197-208. [ Links ]
Hoffman, L.C., Kritzinger, B. & Ferreira, A.V., 2005a. The effects of sex and region on the carcass yield and m longissimus lumborum proximate composition of impala. J. Sci. Food Agric. 85, 391-398. [ Links ]
Hoffman, L.C., Kritzinger, B. & Ferreira, A.V. 2005b. The effects of region and gender on the fatty acid, amino acid, mineral, myoglobin and collagen contents of impala (Aepyceros melampus) meat. Meat Sci. 69, 551-558. [ Links ]
Hoffman, L.C., Kroucamp, M. & Manley, M., 2007a. Meat quality characteristics of springbok (Antidorcas marsupialis). 2. Chemical composition as influenced by age, gender and production region. Meat Sci. 76, 762-767. [ Links ]
Hoffman, L.C., Kroucamp, M. & Manley, M., 2007b. Meat quality characteristics of springbok (Antidorcas marsupialis) 3. Fatty acid composition as influenced by age, gender and production region. Meat Sci. 76, 768-773. [ Links ]
Issanchou, S., 1996. Consumer expectations and perceptions of meat and meat quality. Meat Sci. 74, 197-208. [ Links ]
Koep, K.S.C., Hoffman, L.C., Dicks, L.M.T. & Slinde, E., 2007. Chemical composition of meat and blubber of the cape fur seal (Arctocephalus pusillus pusillus). Food Chem. 100, 1560-1565. [ Links ]
Lawrie, R.A., 1979. Meat Science. 3rd ed. Oxford: Pergamon Press. United Kingdom. [ Links ]
Ledger, H.P., Sachs, R. & Smith, N.S., 1967. Wildlife and food production. Wld Rev. Anim. Prod. 3, 13-36. [ Links ]
Lee, C.M., Trevino, B. & Chaiyawat, M., 1996. A simple and rapid solvent extraction method for determining total lipids in fish tissue. J. AOAC Int. 79, 487-492. [ Links ]
Lushbough, C.H. & Schweigert, B.S., 1960. The nutritive content and the nutritional value of meat and meat products. In: American Meat Science Foundation, The Science of Meat and Meat Products. San Francisco, California: W.H. Freeman and Company, USA. pp. 185-210. [ Links ]
Mostert, R. & Hoffman, L.C., 2007. Effect of gender on the meat quality characteristics and chemical composition of kudu (Tragelaphus strepsiceros), an African antelope species. Food Chem. 104, 565-570. [ Links ]
Onyango, C.A., Izumimoto, M. & Kutima, P.M., 1998. Comparison of some physical and chemical properties of selected game meats. Meat Sci. 49, 117-125. [ Links ]
Ott, R.L., 1998. An Introduction to Statistical Methods and Data Analysis. Belmont, California: Duxbury Press, USA. [ Links ]
Pauw, J., 1993. The development of venison marketing. Forum: The Venison Industry. Irene Animal Production Institute: Agricultural Research Council. pp. 3-12. [ Links ]
Raes, K., De Smet, S. & Demeyer, D., 2004. Effect of dietary fatty acids on incorporation of long chain polyunsaturated fatty acids and conjugated linoleic acid in lamb, beef and pork: A review. Anim. Feed Sci. Technol. 113, 199-221. [ Links ]
Sanudo, C., Enser, M.E., Campo, M.M., Nute, G.R., Maria, G., Sierra, I. & Wood, J.D., 2000. Fatty acid composition and sensory characteristics of calf carcasses from Britain and Spain. Meat Sci. 54, 339-346. [ Links ]
Schönfeldt, H., 1993. Nutritional content of venison. Forum: The Venison Industry. Irene Animal Production Institute: Agricultural Research Council. pp. 51-56. [ Links ]
SAS, 1999. SAS/STAT User's Guide., Version 8, Volume 2. SAS Institute Inc. Cary, N.C., USA. [ Links ]
Simopoulos, A.P., 2002. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed. Pharm. 56, 365-379. [ Links ]
Skinner, J.D., 1984. Selected species of ungulates for game farming in Southern Africa. Acta Zool. Fenn. 172, 219-222. [ Links ]
Van der Waal, C. & Dekker, B., 2000. Game ranching in the Northern Province of South Africa. S. Afr. J. Wildl. Res. 30, 151-156. [ Links ]
Van Zyl, J.H.M., Von La Chevallerie, M. & Skinner, J.D., 1969. A note on the dressing percentage in the Springbok and Impala. Proc. S. Afr. Soc. Anim. Prod. 8, 199-199. [ Links ]
Van Zyl, L. & Ferreira, A.V., 2001. Chemical composition and body component distribution in the springbok, blesbok and impala. In: Sustainable Conservation in Practice. Eds Ebedes, H., Reilly, W., Van Hoven, W. & Penzhorn, B., Proc. 5th Int. Wildlife Ranching Symp., 2001, Pretoria, South Africa. pp. 78-86. [ Links ]
Van Zyl, L. & Ferreira, A.V., 2004. Physical and chemical carcass composition of springbok (Antidorcas marsupialis), blesbok (Damaliscus dorcas phillipsi) and impala (Aepyceros melampus). Small Rumin. Res. 53, 103-109. [ Links ]
Von La Chevallerie, M., 1970. Meat production potential from wild ungulates. Proc. S. Afr. Anim. Prod. 9, 73-87. [ Links ]
Wiklund, E., Pickova, J., Sampels, S. & Lundström, K., 2001. Fatty acid composition of m. longissimus lumborum, ultimate muscle pH values and carcass parameters in reindeer (Rangifer tarandus tarandus L.) grazed on natural pasture or fed a commercial feed mixture. Meat Sci. 58, 293-298. [ Links ]
Zaburnis, E.A. & Cross, R., 1974. A Field Guide to Antelope of South Africa. Natal: Wildlife Society, South Africa. [ Links ]
# Corresponding author. E-mail: lch@sun.ac.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}