










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.40 n.2 Pretoria Jan. 2010
REVIEW
Ruminal acidosis: a review with detailed reference to the controlling agent Megasphaera elsdenii NCIMB 41125
H.H. MeissnerI,*,**; P.H. HenningII; C.H. HornII,***; K-J. LeeuwI; F.M. HaggII; G. FouchéI
IAgricultural Research Council, Private Bag X2, Irene 0062, South Africa
IIMegastarter Biotech Pty LTD t/a MS Biotech, P.O. Box 10520, Centurion 0046, South Africa
ABSTRACT
Ruminal acidosis is discussed with reference to causes and economic and health implications. Distinction is made between the acute form which with proper adaptation to high energy diets is seldom encountered and the more problematic chronic or sub-acute form, commonly referred to as sub-acute ruminal acidosis (SARA). Apart from stepwise transition from roughage to concentrates, methods adopted to reduce SARA include grain treatment to reduce starch degradation, feed additives such as buffers to control ruminal pH, dicarboxylic acids to stimulate the growth of lactate utilisers, antibiotics such as virginiamycin and the ionophores which inhibit the growth of lactate producers, and direct-fed microbials (DFM's), some of which are lactate utilisers but used more often as stimulants of the major ruminal lactate utilisers Megasphaera elsdenii and Selenomonas ruminantium. Some of the feed additives are expensive and their effects on SARA mostly inconclusive. With regard to the ruminal lactate utilising bacteria, the potential of M. elsdenii to control lactic acid has been recognized and some success with patented strains has been achieved. However, these strains have not been commercialised because of one or more reasons which include inadequate growth rate, inability to multiply at a low ruminal pH, non-preferential use of lactate as primary substrate, inability to survive in sub-optimal anaerobic conditions, inhibition by ionophores, inadequate delivery methods to the ruminant and inability to keep on producing acetate when fibre digesters become inhibited. Megasphaera elsdenii NCIMB 41125, selected from the concentrate-fed rumen through stringent screening and a pH-auxostat technique, proved to meet most criteria mentioned above. In addition, the strain is unaffected by most anthelmintics and in-feed antibiotics. Research results show that: a) strain 41125 is highly successful in preventing ruminal pH decline and lactic acid accumulation to SARA levels; b) volatile fatty acid (VFA) production is similar to current in-feed products and the proportional contribution depends on substrate, dilution rate and pH. Propionate as preferred VFA can be promoted further by the synergistic benefits of strain 41125 with some antibiotics; c) feed intake may be enhanced but not consistently, apparently depending upon adaptation procedure followed and dietary composition; d) because of effective ruminal acidosis control, less roughage can be used during adaptation which is of economic benefit; e) animal health associated with the SARA-compromised immune system is improved with administration of strain 41125; f) in feedlot cattle carcass gain and carcass feed conversion may benefit by about 2% and in dairy cattle high producers may produce more milk because strain 41125 enables management to challenge these cows with higher levels of concentrate in total mixed rations (TMR's), and g) drenching sheep with strain 41125 before entering harvested maize fields prevents ruminal acidosis.
Keywords: Megasphaera elsdenii, ruminal acidosis, dairy cattle, beef cattle, sheep, DFM, antibiotics
1. Ruminal acidosis
Ruminal acidosis is considered the most important nutritional disorder in US feedlots (Stock & Britton, 2006) and the dairy industry (Oetzel, 2003). Donovan (1997) estimated the annual cost of sub-acute ruminal acidosis (SARA) to the US dairy industry at $ 500 million to $ 1 billion. The condition affects more than 20% of cows (Hall & Averhof, 2000; Plaizier et al., 2008). There is no reason to believe that the cost in South Africa is less devastating. In addition, apart from compromises to dairy cow health and economics, SARA is of concern for animal welfare reasons, since lameness and laminitis impact significantly on cow comfort and general well being (Hall & Averhof, 2000; Oetzel, 2003).
Ruminal acidosis occurs when there is a sudden excess intake of highly fermentable carbohydrates, primarily starches and sugars. This normally manifests when feedlot cattle without proper adaptation are rapidly transitioned from roughage to high concentrate diets, or when dairy cow intake during transition pre- and post-partum is erratic (Beauchemin & Penner, 2009). It is also a function of the total load of readily fermentable carbohydrates in the rumen (Oetzel, 2003), since dairy cows often experience SARA at the time of maximum carbohydrate consumption (Penner et al., 2007). From a rumen fermentation perspective the main cause of the onset of ruminal acidosis is an imbalance between lactic acid producer and utiliser numbers (Slyter, 1976), resulting in organic acid accumulation (Dawson & Allison, 1988), particularly lactic acid in severe (acute) cases (Nagaraja & Titgemeyer, 2007) and high concentrations of VFA in sub-acute (SARA) and chronic cases (Stone, 2004; Nagaraja & Titgemeyer, 2007; Beauchemin & Penner, 2009). Depending on the severity of the disorder, the result is a drop in ruminal pH to levels that impair proper ruminal and physiological functions.
The literature distinguishes between acute ruminal acidosis and SARA by the following definitions: Acute: ruminal pH <5 - 5.2 (Nocek, 1997; Owens et al., 1998) and lactic acid concentrations of 50 - 150 mM (Nagaraja & Titgemeyer, 2007); SARA: ruminal pH = 5 - 5.5 (Nagaraja & Titgemeyer, 2007), threshold in the dairy cow's pH = 5.8 (Zebeli et al., 2008; Beauchemin & Penner, 2009), or ruminal pH below 5.6 for more than 3 h/24 h (Plaizier et al., 2008) and VFA's at concentrations of 150 - 225 mM (Nagaraja & Titgemeyer, 2007), or daily mean ruminal pH not lower than 6.16 and not lower than 5.8 for 5.24 h/24 h, whereas effective fibre in the total mixed rations (TMR) of dairy cows should be 30 - 33% peNDF (Zebeli et al., 2008). The time threshold is crucial as SARA in dairy cows is characterized by short and long (>3 h) bouts of low ruminal pH; the long bouts of concern because they negatively affect fibre digestion, and decrease the absorptive capacity and damage the rumen epithelium. With time VFA's are absorbed, buffered or passed from the rumen causing the ruminal pH to rise. As a consequence, a cyclical pattern of ruminal pH occurs, causing repetitive periods of SARA (Beauchemin & Penner, 2009). In practice, because of efficient adaptation, acute ruminal acidosis is only encountered in ill-adapted individuals and during the first two to three days of concentrate feeding, whereas SARA is more widespread with major consequences to animal health, production and forage utilisation (e.g. in dairy cows).
2. Methods of addressing ruminal acidosis
Gradual adaptation from roughage to concentrate diets in feedlots is the most common method, but economically unwise because of lower gains and higher usage of expensive roughage (cost per MJ). Buffers and alkalinisers are partially successful (Beauchemin & Penner, 2009), but inconsistent in preventing the decrease in ruminal pH (Stokes et al., 1986; Boerner et al., 1987; Zinn & Boques, 1993). In addition, some of them may depress protozoa and diminish overall microbial activity. Supplementation with dicarboxylic acids such as aspartate, fumarate and malate may be useful as they stimulate the growth of Selenomonas ruminantium on lactate and control ruminal pH in vitro (Martin & Streeter, 1995). However, malate has not been effective in feedlot studies (Martin et al., 1999), whereas other reports suggest that the level of supplementation of malate required to obtain a production response will be cost-prohibitive (Kung et al., 1982; Devant et al., 2007). A promising new technology is steeping grain with lactic acid before feeding (Iqbal et al., 2009). Steeping of rolled barley grain in a TMR fed to dairy cattle modulated ruminal fermentation, decreased the VFA load and increased ruminal pH at critical stages to prevent SARA. The explanation is that the lactic acid decreased the soluble and increased the resistant starch, resulting in less starch degradation in the rumen and more starch digestion in the small intestine (Iqbal et al., 2009).
In the category "antibiotics", ionophores have been useful at high doses in controlling ruminal pH; the mode of action being inhibition of lactate-producing bacteria and reduction of meal size (Nagaraja et al., 1982; Burrin & Britton, 1986; Newbold & Wallace, 1988; Owens et al., 1998). Some though do not seem to be effective in controlling lactic acid (Steyn, 1996), or preventing the secondary growth of lactobacilli (Newbold & Wallace, 1988) or the accumulation of VFA in the rumen (Mutsvangwa et al., 2002; Erickson et al., 2003), and are therefore unlikely to alleviate or prevent SARA. Virginiamycin controls the growth of ruminal lactic acid-producing bacteria and thereby is successful in controlling ruminal pH and lactic acid accumulation (Nagaraja & Taylor, 1987; Coe et al., 1999; Rinttilä et al., 2009). However, under certain feeding conditions addition of virginiamycin did not reduce the risk of ruminal acidosis (Godfrey et al., 1995; Thorniley et al., 1998). Virginiamycin and the ionophores are selective inhibitors of gram-positive bacteria (Nagaraja & Taylor, 1987), and are the most in-feed antibiotics approved to control lactate-producing bacteria and liver abscesses in feedlot cattle. In vitro results of Apajalahti & Alaja (2008) and Rinttilä et al. (2009), however, suggested that the reduction in lactic acid content observed with the antibiotics AS-700, TM-200, zinc bacitracin, flavomycin and tylosin is merely the result of depression of overall bacterial fermentation.
Although antibiotics by and large have been effective in controlling ruminal acidosis, concern regarding their use in the US animal feed industry has increased (Krehbiel et al., 2003). In the European Union (EU) the use of antibiotic growth promoters has been banned and tighter control has been exerted on therapeutic antibiotics. The result is increased interest in the effects of direct-fed microbials (DFM's) on animal health and performance. In the rumen DFM's include bacteria, fungi and yeasts. In the context of ruminal acidosis, the major lactate utilising bacteria, Megasphaera elsdenii and S. Ruminantium, are obvious candidates. Of the two, M. elsdenii is more important (Counotte et al., 1981) because it utilises 60 to 95% of the lactate available in the rumen. It also shows preference for lactate as a substrate (Russel & Baldwin, 1978; Marounek et al., 1989), but has the ability to simultaneously metabolise lactate, glucose and maltose, and therefore competes with lactate-producers for substrate. S. Ruminantium, on the other hand, is unlikely to be effective as it undergoes catabolite repression (Russel & Baldwin, 1978) and is comparatively acid-intolerant (Mackie & Gilchrist, 1979).
Lactobacilli may assist in preventing ruminal acidosis because their presence may help the rumen population to adapt to measurable amounts of lactate (Yoon & Stern, 1995; Nocek et al., 2002). They may also provide a competitive advantage to M. elsdenii. However, because lactic acid is rapidly produced during concentrate exposure by the lactate-producing organisms in the rumen, quantitatively it is unlikely that the adaptation can be significant (Brown & Nagaraja, 2009). Propionibacterium is also a lactate utiliser (Krehbiel et al., 2003), but its role in shifting ruminal fermentation towards propionate production to the benefit of the host has rather been emphasized. In this context it should improve the VFA ratio if lactate is metabolized by lactate-utilising organisms. Theoretically, the combination of Propionibacterium and lactobacilli should be beneficial because in the presence of Lactobacillus strains more propionic acid and less butyric acid are produced (Krehbiel et al., 2003), whereas post-ruminally, lactobacilli as a further benefit to health and production display probiotic characteristics (Brown & Nagaraja, 2009). The results in this regard, however, are inconsistent. Krehbiel et al. (2003) and Brown & Nagaraja (2009) summarized several positive findings whereas other studies could find no evidence that Propionibacterium and Lactobacillus strains enhance ruminal fermentation (Raeth-Knight et al., 2007) or animal performance (Elam et al., 2003).
The most common fungus used in ruminant production is Aspergillus oryzae. Some strains stimulate the growth of M. elsdenii and S. ruminantium (Nisbet & Martin, 1993; Waldrip & Martin, 1993), thereby reducing the risk and severity of ruminal acidosis. Similarly, the yeast, Saccharomyces cerevisiae, stimulated the growth of M. elsdenii in vitro by providing critical nutrients (Callaway & Martin, 1997; Hutjens, 1999). S. cerevisiae may also have a direct effect on lactic acid production, since one strain proved to exceed Streptococcus bovis in utilisation of sugars (Chaucheyras et al., 1996). Production results, however, have been inconsistent (Brown & Nagaraja, 2009) and the commercial products are comparatively expensive (Hutjens, 1999).
Work on M. elsdenii has progressed significantly during the last 30 years, with patented strains showing promising characteristics (Robinson et al., 1992; Hino et al., 1994; Kung & Hession, 1995; Wiryawan & Brooker, 1995; Klieve et al., 2003), yet with very little application in the animal industry. Main reasons are one or more of the following: The strains were not isolated from rumens of animals on high concentrate diets; they are not ionophore resistant which is a prerequisite on feedlot and dairy diets; they have inadequate growth rates and biomass production; they do not preferentially use D- and L-lactate as substrate, or do not have the ability to grow on reducing sugars as well as on lactate which is necessary in the concentrate-fed rumen; most cannot maintain multiplication at a low ruminal pH and they predominantly produce propionate and butyrate rather than acetate (Horn et al., 2009b). At low ruminal pH, the activity of fibre-degrading bacteria that primarily ferments to acetate is decreased, which creates a shortfall in acetate availability to the host. The Megasphaera strain(s) must accommodate the shortfall by also fermenting lactate to substantial amounts of acetate. From a commercialization point of view difficulties have been experienced with stability and in maintaining viable cells because the organism is strictly anaerobic. A summary of results in relation to M. elsdenii and ruminal acidosis reveals the following: Kung & Hession (1995) found that strain B 159 prevented accumulation of lactic acid when challenged with highly fermentable carbohydrates; Robinson et al. (1992) demonstrated that strain 407A was promising in preventing lactic acid acidosis in steers; strain NIAH 1102 preferred lactate to glucose as substrate (Hino et al., 1994); strain YE 34 rapidly established a viable lactic acid-utilising bacterial population in the rumen of inoculated compared to uninoculated cattle (Klieve et al., 2003) and strain JDB 301 maintained rumen stability (pH >6.0) for days following grain feeding (Wiryawan & Brooker, 1995).
3. M. elsdenii strain NCIMB 41125 (previously known as isolate CH4)
Research on strain 41125 and similar M. elsdenii strains commenced in 1994 at Irene (Agricultural Research Council, Private Bag X2, Irene 0062, South Africa) with the aim of overcoming the limitations of the strains on the shelf, as discussed above. The strains were obtained from the rumens of concentrate-adapted fistulated dairy cows and slaughtered feedlot cattle, and were subjected to stringent selection criteria (ARC/Kemira Phosphates, 2006; Horn et al., 2009a). The strategy was to select those strains from the rumen content which could thrive under sustained, simultaneous pressure of a combination of discriminatory factors, using the pH-auxostat version of continuous culture. By that means and by keeping the pH differential between control point and the incoming poorly-buffered medium small, a further selection pressure was applied. This was singling out from the heterogeneous population, the fastest growing individuals under the given constant environmental conditions. These individuals determined the rate at which lactic acid was converted to weaker acids, and thus the rate of deviation from a pH set point. This, in turn, determined the rate at which fresh medium was to be added, i.e. the dilution rate of the system. In time the slower growing species and strains in the population were washed out of the bioreactor. Of the more promising thriving strains, strain 41125 excelled and met most of the set criteria (see "Characteristics of strain 41125" below).
3.1. Characteristics of strain 41125
Phylogenetically, comparative 16S rRNA sequencing results revealed that the M. elsdenii strains, which phenotypically are representative of a larger homogenous group, are 97 - 99% similar. With respect to the two reference type strains of M. elsdenii (ATCC 25940 and ATCC 17752), the major nucleotide sequence difference with strain 41125 occurs at nucleotide positions 529-536 and 1105-1120 (Wiederhold, 1994; ARC/Kemira Phosphates, 2006).
In terms of characteristics of significance to prevention or alleviation of ruminal acidosis, the following provides a summary:
- Strain 41125 displays a high growth rate of up to 0.938/h (ARC/Kemira Phosphates, 2006; Horn et al., 2009b) which is markedly faster than any other documented M. elsdenii strain, resulting in a biomass output of 0.39 g (L/h). Its biomass accumulation provides strain 41125 more tolerance to environmental stressors such as oxygen exposure which makes it highly robust for commercial use.
- Strain 41125 is able to metabolize lactate and produce fermentation end products at pH levels well below 5.5 and it is able to replicate at a measurable μmax at pH = 4.5 (ARC/Kemira Phosphates, 2006; Horn et al., 2009b). Indigenous and most patented strains of M. elsdenii have minimal activity and replication ability at a pH range of 4.5 - 5.5.
- Strain 41125 has the ability to convert lactate to (primarily) acetate and continues to do so at low pH levels (ARC/Kemira Phosphates, 2006; Horn et al., 2009b). The results furthermore show that the ratio among VFA's produced, particularly propionate, butyrate and valerate, depends on dilution rate and pH.
- The isolate is unaffected by ionophores, by most in-feed anthelmintics and antibiotics (ARC/Kemira Phosphates, 2006; Apajalahti & Alaja, 2008; Apajalathi et al., 2008; Rinttilä et al., 2009).
3.2. Control of pH and lactic acid
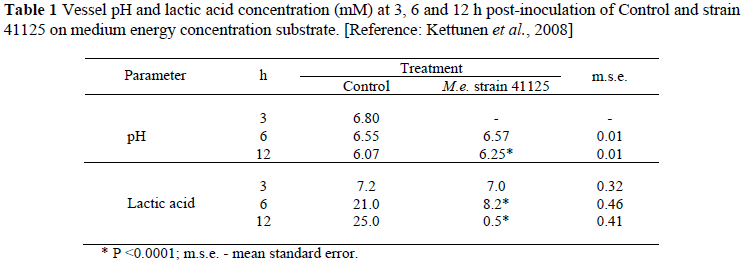
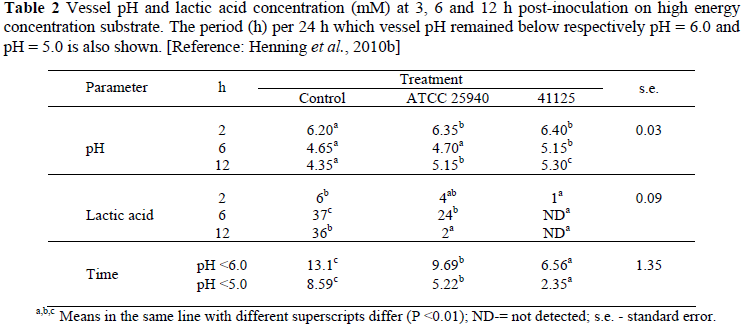
In in vitro simulation where the inoculum was based on a diet of 50% maize silage, 25% barley meal and 25% soyabean meal, i.e. medium energy as for dairy cows (Kettunen et al., 2008; Rinttilä et al., 2009), or a roughage-based substrate, supplemented with a high level of maltose just prior to strain 41125 administration, i.e. high energy as for feedlot cattle (Henning et al., 2010b), the trends corresponded. The results are shown in Tables 1 and 2, respectively. On the medium energy inoculum, strain 41125 was administered at a concentration of 3.1 × 106 cfu's/mL and on the high energy inoculum at a concentration of 5 × 105 cfu's/mL, respectively. On the medium energy inoculum, Control (containing no culture) was compared with strain 41125 and on the high energy inoculum with, additionally, a type strain of M. elsdenii (ATCC 25940).
Corresponding to the higher energy concentration in the inoculum medium, vessel pH declined to lower levels than in the case of the medium energy inoculum (Table 2 vs. Table 1). The decline was to less than pH = 5.5, simulating SARA according to definition, and even to less than 5 which corresponds with acute ruminal acidosis. Due to effective utilisation of accumulating lactic acid (Tables 1 and 2), strain 41125 prevented vessel pH to decrease to clinical levels. Plaizier et al. (2008) and Zebeli et al. (2008) maintained that the decline in ruminal pH per se as indication of SARA is not as important as the time per 24 h it remains below the indicator pH level. Consequently the time per 24 h vessel pH remained below respectively 6 (when ruminal cellulolytic activity begins to be impaired) and 5 (acute acidosis) was also calculated. The results in Table 2 suggest that in comparison to Control and the type strain, strain 41125 should prevent ruminal pH to decrease to the levels of concern.
In vivo results are expected to differ somewhat from those in vitro or batch culture because of confounding effects in the rumen of fermentation end product accumulation - disappearance ratios, dilution rate and passage from the rumen. One of the consequences is that the pH - lactic acid relationship may not be as strong as in batch culture because of variable VFA contribution. Also, SARA is mainly a function of VFA concentration (150 - 225 mM) (Nagaraja & Titgemeyer, 2007), often with lactic acid concentrations in the dairy cow of <1 mM (Beauchemin & Penner, 2009). In relation to strain 41125, Aikman (Aikman, 2008; Aikman et al., 2009a) accordingly measured detectable levels of lactic acid only in some cows of both Control and strain 41125 treatments. Nevertheless, ruminal pH tended to be longer below 5.6 in Control (4.03 h vs. 3.35 h for strain 41125), apparently attributable to a higher VFA load. The effect of highly fermentable carbohydrates on ruminal lactic acid levels is further illustrated in a study by Hagg et al. (2010) at the University of Pretoria. They fed two groups of dairy cows TMR's containing respectively 60 : 40 and 70 : 30 concentrate - roughage ratios and reported mean lactic acid concentrations (mM) of respectively 2.75 (Control) and 2.50 (strain 41125), and 5.80 (Control) and 4.32 (strain 41125), suggesting a slight (not significant) but consistent advantage to strain 41125 drenching. The 60 : 40 ratio corresponded with the highest energy concentration in the study of Aikman.
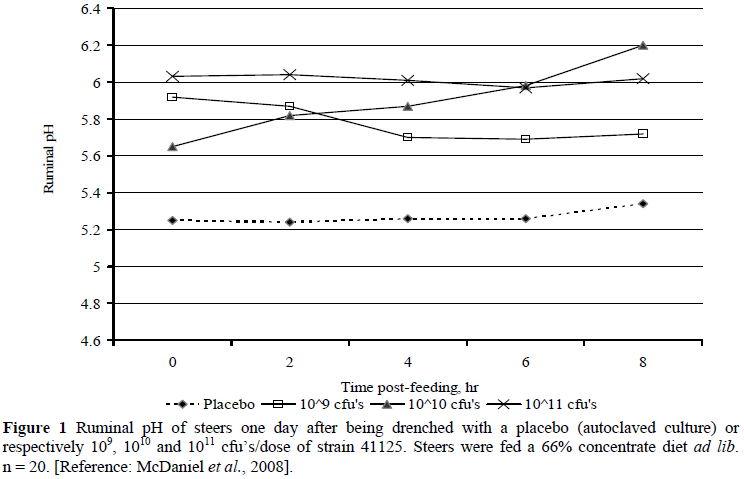
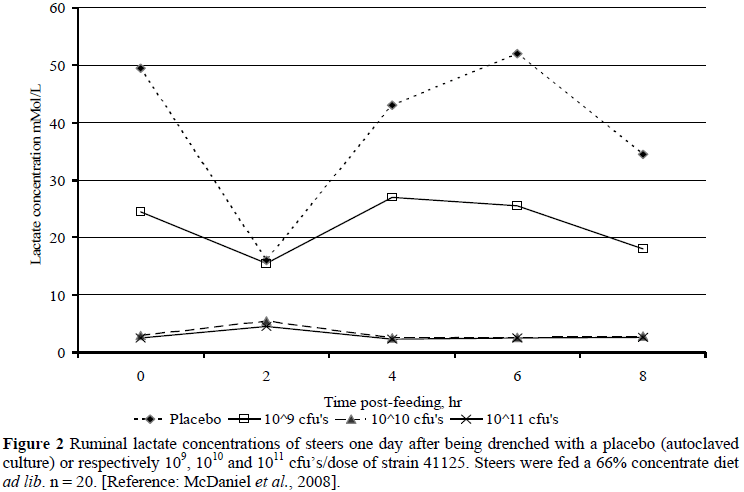
In in vivo feedlot simulation, the studies of Henning et al. (2010a; b) at Irene and McDaniel et al. (2008; 2009a; b) at Kansas State University (KSU) are illustrative. Henning et al. (2010b) fed sheep on roughage and then without adaptation 1000 g of maize followed by 300 g of maltose through the rumen cannula. One hour later they administered into the rumen either strain 41125 at 1011 cfu's or a placebo. Ruminal pH in Control remained below 5 from 8 h to 24 h post-administration whereas for strain 41125 ruminal pH remained above 5.5 for the entire period (P <0.001). Lactic acid in Control increased progressively to >55 mM at 10 h, whereas it remained below 10 mM in the strain 41125 administered treatment (P <0.001), confirming earlier results with a similar approach (Henning & Steyn, 1995; Steyn, 1996). With more typical feedlot diets with sheep and cattle, the trends corresponded (Henning et al., 2010a). For example, in the cattle trial ruminal pH approached 5 between days 9 and 17 post-drenching in Control in contrast to three strain 41125 treatments (respectively 109, 1010 and 1011 cfu's/dose), which averaged just above pH = 5.5. Lactic acid concentrations on days 2 and 3 post-drenching averaged 10.1 - 20.8 mM in Control vs. 0.28 - 0.47 mM in the strain 41125 administered treatments. Similar trials were conducted at KSU with US feedlot diets (McDaniel et al., 2008; 2009a; b), showing corresponding results the day after drenching (Figures 1 and 2). The KSU results suggest that the influence of strain 41125 on ruminal pH and removal of lactic acid are dose size dependent, but both appeared to level off between 1010 and 1011 cfu's/dose, which is supported by earlier work at KSU (2003, unpublished) and the data of Henning et al. (2010a). A dose of 1010 cfu's translates to a concentration in the rumen of cattle of the order of 105 cfu's/mL. In sheep the results of Henning & Steyn (1995) suggested that concentrations of 109 to 0.5 × 1010 cfu's/dose are optimal.
Since strain 41125 is administered almost immediately after first concentrate exposure, control of ruminal pH and lactic acid is most noticeable during the first 2 - 4 days after introduction of concentrates and drenching of the organism (Henning & Steyn, 1995; Steyn, 1996; McDaniel et al., 2008; McDaniel et al. 2009b; Henning et al., 2010a). This is a function of the size of the dose of strain 41125 introduced and the slower responding endogenous M. elsdenii strains, the latter which thereafter progressively populate the rumen environment. This was confirmed by quantitative rt-PCR and cell flow cytometry of rumen samples at Alimetrics, Finland (Rinttilä, 2008). Although the 2 - 4 day adaptation scenario is most critical, cattle are also vulnerable to SARA at later stages of the feeding period. This may occur in the feedlot when concentrates are increased from the grower to the finishing diet or when after a spell of low intake, cattle overeat. This may also occur in dairy cattle on diets containing progressively greater quantities of carbohydrate until peak consumption. The data of McDaniel et al. (2009b) suggested that strain 41125 keeps its competitive advantage and will outgrow other lactate utilising organisms whenever there is a sudden increase in concentrate (substrate) entering the rumen. The data on this subject should be expanded through dedicated investigations, including the long-term survival of strain 41125 in the rumen.
In conclusion, strain 41125 has been shown to rapidly metabolize lactic acid and stabilize ruminal pH. Dosing of ruminants with strain 41125 is valuable when they are comparatively quickly transitioned from roughage to concentrate diets. In contrast, other DFM's may be less effective in this regard: Apajalahti (2007) and Kettunen et al. (2008) evaluated products containing Lactobacillus species, Propionibacterium species and live yeast in a comparative batch culture study with strain 41125. They found very little evidence that these products enhance fermentation, supporting in vivo evidence of Raeth-Knight et al. (2007).
Therefore, in association, their results suggest that it is unlikely that these products administered either alone or together with strain 41125 will assist in combating or preventing acidosis. However, when strain 41125 was tested together with a yeast hydrolysate they found synergistic effects. This could mean that the efficiency of strain 41125 drenching may be enhanced if administered in the presence of particular nutrients, which supports the review on M. elsdenii by Brown & Nagaraja (2009). Also, these DFM's should not be ruled out on account of the results of Raeth-Knight et al. (2007) and Kettunen et al. (2008), since positive results have been reported (Krehbiel et al., 2003; Brown & Nagaraja, 2009) and patents registered. The subject of strain 41125 in the presence of other DFM's warrants further investigation.
3.3. Volatile fatty acids
Total ruminal VFA concentration at the recommended dose of strain 41125 does not appear to be altered compared to a Control (Steyn, 1996; KSU, 2003 [unpublished]; Aikman, 2008, Aikman et al. 2009a; Hagg et al., 2010; Henning et al., 2010a). However, in vitro, Apajalahti (2007) showed that where strain 41125 was introduced at very high levels (107 vs. 106 or 105 cfu's/mL) total yield of VFA's was reduced. In vivo, the results of McDaniel et al. (2008) and Henning et al. (2010a) (1011 vs. 1010 or 109 cfu's/mL) tended to support this observation. In the presence of the antibiotics, monensin, AS-700, TM-200, flavomycin and zinc bacitracin, strain 41125 produced less VFA's in in vitro than alone, because the antibiotics depressed microbial fermentation (Apajalahti & Alaja, 2008; Rinttilä et al., 2009). In contrast, virginiamycin had very little effect on the growth of strain 41125, whereas in the presence of tylosin, strain 41125 had a competitive advantage to other endogenous bacteria. Both reflected in VFA production and thus positive synergism.
The yield and proportional contributions of individual short chain fatty acids in the presence of M. elsdenii is influenced by pH (Counotte et al., 1981), substrate (Rogosa, 1984) and dilution rate. With regard to dilution rate, Horn et al. (2009b) demonstrated in vitro changes in yields of propionic, butyric and valeric acid (Table 3), the highest yield of propionic acid being at a dilution rate of 0.83/h.
With M. elsdenii in general, when pH decreases, fermentation is shifted from propionic acid (favourable) to butyric (less favourable) and valeric acid (Counotte et al., 1981; Marounek et al., 1989). Aikman ( 2008) and Aikman et al. (2009a) found that with dairy cows where ruminal pH remained around 6, strain 41125 compared to Control shifted fermentation to more propionic acid and a lower acetic - propionic acid ratio. Hagg et al. (2010), also with dairy cows but at a ruminal pH of around 5.8, found no significant difference between Control and strain 41125 treatments, whereas Kettunen et al. (2008) and Rinttilä et al. (2009) in vitro and Steyn (1996), KSU (2003, unpublished) and Henning et al. (2010a) in vivo (feedlot diets) at still lower (<5.5) pH's, reported a shift towards butyric and valeric acid with introduction of strain 41125. This suggests that: (a) fermentation of strain 41125 conforms to that of other M. elsdenii strains, and (b) when ruminal pH hovers around SARA levels, VFA composition may be less favourable. Thus, either ruminal pH must be elevated further or a propionic acid stimulator or both are required. Propionibacterium strains alone or in the presence of lactobacilli, selected antibiotics and ionophores are candidates, but since ionophores are routinely fed in feedlot trials where strain 41125 is tested, their effect is already in the results. In vitro, the results with a Propionibacterium strain in combination with a Lactobacillus strain showed no response (Apajalathi, 2007), whereas the antibiotics AS-700, TM-200, tylosin, virginiamycin and zinc bacitracin in the presence of strain 41125 shifted fermentation from butyrate to propionate (Apajalathi & Alaja, 2008; Rinttilä et al., 2009). The response of strain 41125 in conjunction with these products should receive further attention.
In the trial of McDaniel et al. (2008) butyric acid increased and propionic acid decreased when the highest dose size (1011 cfu's/mL) of strain 41125 was administered. The highest concentration of propionic acid and the most favourable acetic - propionic acid ratio were obtained with the medium dose size (1010 cfu's/mL), which corresponds with the recommended dose size. However, since ruminal pH, dilution rate and substrate are confounding factors the influence of strain 41125 on VFA concentrations and ratios remains unpredictable.
3.4. Dry matter intake
Feed intake is expected to be enhanced or to vary less when strain 41125 is administered, at least during the phase when the accumulation of lactic acid by the organism is prevented or removed. In practice, this phase coincides with the period when the diet of the animal is transitioned from roughage to concentrate, i.e. during adaptation. Some feedlot results do not support this expectation (Steyn & Henning, 1996; Leeuw et al., 2009; McDaniel et al., 2009b), but the majority of results where sheep and cattle were fed on station apparently conforms (Henning & Steyn, 1995; Drouillard, 2004; KSU, 2003 [unpublished]; Henning et al., 2010a; b). The intake advantage in some of these cases was maintained for even longer periods than during adaptation. For example, cattle drenched with strain 41125 in the Drouillard (2004) trial tended to maintain higher intakes throughout the experiment and their intakes were more consistent (P = 0.07) (Figure 3) than Control cattle. Henning et al. (2010b) reported a 16% decline in roughage intake in Control sheep following highly fermentable carbohydrate administration through the rumen cannula vs. only 9% in strain 41125 dosed sheep (P <0.05). In the experiments of Henning et al. (2010a) feed intake of lambs was 29% higher for strain 41125 dosed lambs than for Control between days 1 and 11 (P <0.006) and 6% higher between days 1 and slaughter at 50 days (P <0.01). In their cattle trial an average difference of about 21% was maintained with the difference on days 12, 25, 33 and 34 significant at P <0.01 and the difference at days 10, 28, 32 and 35 significant at P <0.05. Both papers of Henning and co-workers also reported less variation in day-to-day feed intake in the strain 41125 dosed animals. In this regard it is interesting that the study of Leeuw (unpublished) revealed less variation in intake for strain 41125 administered than monensin treatments between days 1 and 35.
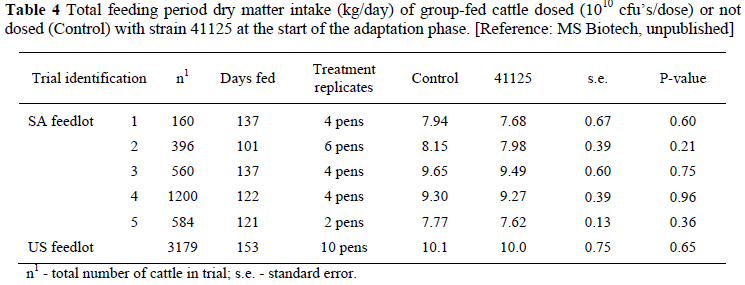
Feed intake in commercial feedlot trials as tested by a probability test does not show a significant advantage for strain 41125 dosed cattle, either in the adaptation phase or the total feeding period. Average feed intakes for the total feeding period are shown in Table 4.
There may be a number of reasons why intake is not always enhanced by strain 41125: It is a function of concentrate processing (Henning & Steyn, 1995), adaptation procedure (Drouillard, 2004) and feed energy concentrations, with commercial feedlots generally being more conservative in approach.
With TMR's fed to dairy cows drenching with strain 41125 resulted in a variable feed intake response (Aikman, 2008; Aikman et al., 2009b; Hagg et al., 2009; 2010). These authors fed two groups of cows for 60 - 100 days post-partum on diets with respectively high and low energy concentrations; each group having a Control and a strain 41125 administered treatment. Aikman (Aikman, 2008; Aikman et al., 2009b) in addition, also conducted a ruminal fermentation study for the first 28 days post-partum on the high energy concentration diet. When in the production study of Aikman the results of only the highest producers (presumably more prone to SARA) were considered for the first 21 days post-partum, intake was lower for the strain 41125 dosed cows than the Control cows (P <0.012) on the high energy concentration diet, but not significantly different on the low energy concentration diet. The results for the cows in the ruminal fermentation study which had similar production levels concurred (Figure 4). In contrast, the high producers on a diet with similar energy concentration at Irene (Erasmus, unpublished as reported by Hagg et al., 2009) showed the opposite for the entire 80 days of measurement; intake was higher on the strain 41125 administered treatment than for Control (P <0.06). As with feedlots, one suspects that the difference in feed intake response to strain 41125 drenching in dairy cow TMR diets is feed source and processing related: Aikman fed barley and wheat as energy sources, Erasmus fed maize meal and steam flaked sorghum and Hagg et al. (2010) only maize meal. Protein and NDF sources also differed markedly.
3.5. Adaptation and roughage use
Feedlot cattle are normally transitioned gradually from roughage to all concentrate diets over a period of 14 to 21 days and even longer in feeding systems where starter, grower and finisher diets are used. The adaptation phase is costly because cattle grow slowly, and because of the number of diets and feed sources required to be mixed. In addition, the roughage sources are expensive per unit available energy and because of high transport, storage and insurance costs. It therefore makes sense to limit the cost of adaptation by shortening the period and using less roughage. The question is whether drenching with strain 41125 can assist in reducing roughage.
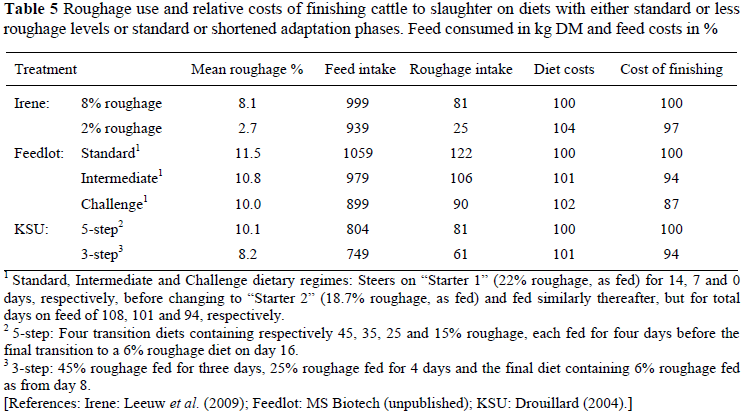
Table 5 shows data of trials where cattle were dosed with strain 41125 and which provide evidence of improved feed costs with either less roughage or enhanced adaptation. In a trial at Irene, Leeuw et al. (2009) fed steers diets with either 8% or 2% roughage in the final diet. Adaptation occurred over 14 days, commencing respectively with 18% and 12% roughage and finishing with 8% and 2%. In a second trial at a commercial feedlot the adaptation phase was shortened progressively, using the distinguishing terms for the three treatments of Standard, Intermediate and Challenge. In the third trial at KSU, Drouillard (2004) adapted the cattle either according to the standard 5-step procedure or an enhanced 3-step procedure. The data in Table 5 were calculated from the results and dietary costs of the trials.
The results suggest that it should be possible to reduce roughage acquisition for the feedlot by at least 25%, with substantial benefits to transport and storage costs.
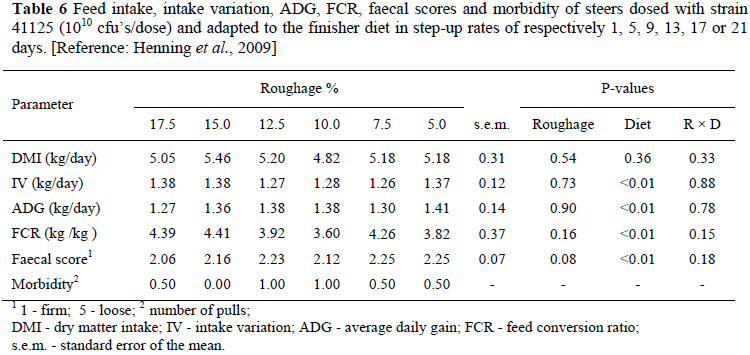
To test the hypothesis that drenching with strain 41125 may assist in the quest for less roughage during adaptation, a study was conducted at Irene with weaner steers in individual pens. They were dosed with strain 41125 on day 1 after all-roughage feeding and then fed diets either based on maize meal or hominy chop for 35 days (Henning et al., 2009). Six dietary step-up rates were employed, attained by mixing a basal starter diet with a finisher diet in the required ratio to obtain roughage levels of respectively 5, 7.5, 10, 12.5, 15 and 17.5% on day 1, whereafter the steers were transitioned to the finishing diet (5% roughage) on days 1, 5, 9, 13, 17 or 21. The combined results of the maize meal and hominy chop diets are shown in Table 6.
Dietary effects were mostly significant (Table 6) with hominy chop diets showing higher intakes and less intake variation than maize meal diets, a result which reflected in better ADG and FCR. The effect of step-up rate depicted in Table 6 as percentage initial roughage, however, was not significant, neither was the interaction between step-up rate and diet. Faecal score tended (P = 0.08) to be looser towards the shorter step-up rate, which with regression analysis was confirmed with maize meal but not with hominy chop diets. Faecal consistency, however, did not affect the other measured parameters in relation to step-up rate. The results suggest that by drenching with strain 41125, transition from roughage to concentrate finishing diets can be accomplished without gradual adaptation. The amounts of roughage consumed by the steers over the 35 day trial period were respectively 13.8, 12.8, 10.7, 8.89, 9.22 and 9.07 kg (step-up rate, P <0.01; diet, P = 0.81; interaction, P = 0.67) for the initial roughage levels of 17.5, 15.0, 12.5, 10.0, 7.5 and 5.0%, which confirms a saving in roughage usage. Although promising, the topic of faster adaptation and using less roughage requires further investigation because experience with regard to different feed sources, feed processing and feeding practices in feedlots is required; also because limited sheep data showed poorer feed intake and growth when strain 41125 was drenched to lambs receiving diets with 0 or 5% roughage than lambs receiving diets containing 10% roughage (Steyn & Henning, 1996).
3.6. Animal health
Ruminal acidosis has both immediate or short term and longer term (indirect) consequences. The short term effect is related to the discomfort, inflammation (Ametaj et al., 2009) and poorer or variable performance of the animal suffering from acidosis. The longer term effects may impact on associated conditions such as liver abscesses, continued inflammation and laminitis/lameness, and apparent non-associated conditions of diseases because of endotoxins which activate the immune system (Plaizier et al., 2008). In this context it is known that lipopolysaccharides and low ruminal pH from high proportions of grain in the diet are associated with increased permeability of the rumen and colon mucosa. This apparently results in increased endotoxins due to translocation or release of cytokines into the bloodstream (Gozho et al., 2006; Emmanuel et al., 2007; 2008).
One suspects that drenching with strain 41125 may reduce the incidence of these conditions or mitigate their effects. The results from commercial feedlot trials, however, are inconsistent. In dairy operations, a number of farmers who routinely dose with strain 41125 testified that one of the main benefits is a lower incidence of ill-health and veterinary costs. Limited evidence from on-station experiments apparently supports their observations. For example, in the dairy cow study of Hagg et al. (2010), the researchers had to withdraw respectively 4 and 7 cows in the low concentrate and high concentrate control treatments due to ill-health induced poor intake, and only one each in the corresponding strain 41125 administered treatments. In the feedlot study of Leeuw et al. (2009) the authors distinguished between steers treated for all (unspecified) cases of ill-health and steers exhibiting digestive disturbances such as diarrhoea and bloat. Their diets contained respectively 2 and 8% roughage in the finishing mix. On the 2% roughage diet, respectively 21 and 8 steers were treated for ill-health on the Control and strain 41125 administered treatments and 5 and 7 on the 8% roughage diet. The corresponding numbers treated for digestive disturbances were 15 and 5, and 4 and 4, respectively. The data reflected a significant diet × treatment interaction, with strain 41125 assisting in reducing cases of disturbance/ill-health on the 2% roughage (higher energy) diet.
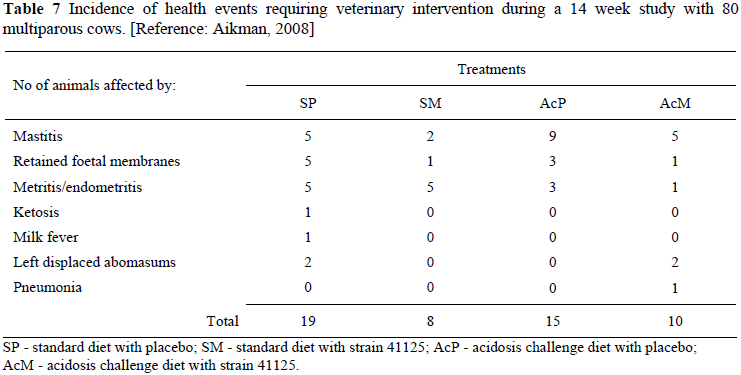
Aikman (2008) provided more detail by distinguishing between events of ill-health (Table 7). The diets fed to dairy cows were termed standard (S) and acidosis challenge (Ac) and they were dosed with either a placebo (P) or Megasphaera (M) (strain 41125) 3 and 12 days post-partum.
The data did not have sufficient observations to provide statistical analysis (Table 7), but the trends support the belief of dairy farmers. Thus, there is sufficient collateral evidence that drenching with strain 41125 improves animal health but dedicated long term trials are required to obtain conclusive results.
3.7. Animal production
3.7.1. Feedlot cattle
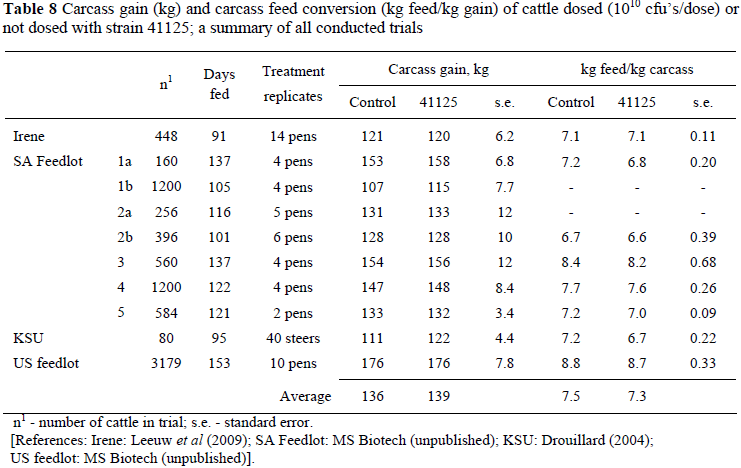
Brown & Nagaraja (2009) cited a study of a population-level analysis on performance response of DFM's in commercial feedlots, accommodating 118 feedlots that used DFM's and 149 that did not. Weight gains were 1.4 to 1.9% more rapid in DFM-fed cattle than in cattle not fed DFM's, whereas they required 1.9 to 3.9% less feed/unit gain. With high animal variation, different feeds and environmental conditions as confounding sources of variation, many large scale trials tested in this way or with meta-analysis are required to confirm such small differences statistically significant. Therefore, only trends for trials with treatments with or without strain 41125 can be reported here, supported by probability tests. Probability tests on the available data did not show trends for ADG or FCR, but did show positive responses for strain 41125 administered treatments (P <0.05) for carcass gain (2.2%) and feed required/kg carcass (2.7%) (Table 8). The magnitude of the difference appears to be similar to those of other DFM's.
3.7.2. Dairy cattle
Production responses to drenching with strain 41125 are most comprehensively reported in the study of Aikman (Aikman, 2008; Aikman et al., 2009b). In a 2 × 2 factorial design, cows in a 14-week production study were fed either a standard (D) or an acidosis challenge (Ac) diet and dosed either with a placebo (P) or Megasphaera (strain 41125) (M). In a 1 - 28 day post-partum ruminal fermentation study the Ac diet with or without strain 41125 was studied further. Results of the production study in Table 9 are reported for all cows (n = 80) and for the highest milk-producers (presumably more vulnerable to SARA) (n = 32; producing more than 10000 L/lactation). To have comparable data with the ruminal fermentation study, the authors also evaluated the production data of the high milk-producers between days 1 and 21 post-partum.
Dietary effects for most observations were significant (Table 9), as expected. Milk yield did not differ significantly between the placebo and strain 41125 administered treatments, but the high milk-producing cows on AcM tended to produce more milk than the high milk-producing cows on AcP, whereas the opposite was true for SM vs. SP (P = 0.08). When tested between days 1 and 21 post-partum, i.e. corresponding to the period of rapid increasing intake of concentrates, the high milk-producers dosed with strain 41125 had higher milk yields than the high milk-producers dosed with a placebo (P = 0.001), a result which was repeated for the high milk-producers in the ruminal fermentation trial (P = 0.05). The higher milk yields on AcM reflected in better efficiencies of production, partly because DM intake was lower, as discussed under section 3.5, and probably also because rumen fermentation enhanced propionic acid production (Aikman et al., 2009a). Propionic acid is a recognized glucose precursor in milk synthesis.
Corresponding milk yield results have been found by Erasmus as reported by Hagg et al. (2009), although in that trial the difference carried through to 80 days post-partum. With all 60 cows the difference between +strain 41125 and -strain 41125 approached significance (P = 0.10; 35.1 L/day vs. 33.1 L/day). With the 20 highest milk-producers on the higher energy diet, those on +strain 41125 produced 39.3 L/day vs. 35.9 L/day for cows on -strain 41125 (P = 0.06), in contrast to the 20 cows on the lower energy diet (35.2 L/day vs. 34 8 L/day) where the difference was not significant (P = 0.82). Thus, as with the Aikman (2008) results, the high milk-producing cows on the higher energy diet benefitted most from the administration of strain 41125. The results of Hagg et al. (2010) for high milk-producers in a similarly designed trial were not significantly different but did show corresponding trends. The average milk yields on their lower energy concentration diet were respectively 42.3 L/day for -strain 41125 and 43.7 L/day for +strain 41125, whereas on their high energy diet the yields were respectively 46.6 L/day for -strain 41125 and 47,8 L/day for +strain 41125. In conclusion, the three studies suggest that with strain 41125 drenching it is possible to feed TMR's with higher energy concentrations than what is normally recommended, to the benefit of milk yield of high producing cows.
On the AcM treatment of Aikman (Aikman, 2008; Aikman et al., 2009b) milk fat percentage in high producers (Table 9) was reduced in both the production trial and the ruminal fermentation trial, which partially can be explained by a lower acetate - propionate ratio (Aikman et al., 2009a) and the milk yield time-associated inverse relationship between milk yield and milk fat concentration. Hagg et al. (2009) reported the opposite in the Erasmus study, with the +strain 41125 treatments showing higher milk fat percentages, but with a significant treatment × dietary interaction, whereas Hagg et al. (2010) reported no significant difference in milk composition parameters. One suspects that fermentation substrate in the rumen is the main variable explaining the difference: Aikman fed barley and wheat as primary energy sources, Erasmus steam flaked sorghum and maize meal and Hagg et al. (2010) only maize meal.
Body weights and body condition score did not differ due to drenching treatment (Table 9). However, the change in body weight with lactation of the higher producers did, respectively, more positive for the higher energy diet (P = 0.02) and more positive for strain 41125 dosed cows. In support, Hagg et al. (2009) reported higher body weights (P = 0.02) and condition scores (P = 0.02) for strain 41125 dosed cows on the higher energy concentration diet, suggesting that because body reserves recover quickly, recycling and inter-calf period may benefit from drenching with strain 41125. These interpretations warrant further research.
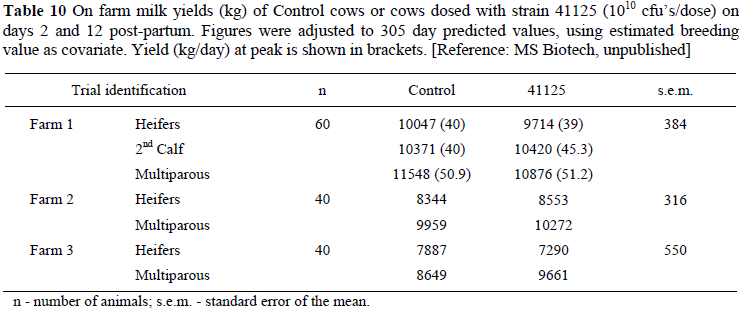
In farm trials milk yield responses after drenching cows with strain 41125 are inconsistent, probably because of limited degrees of freedom and varying environments, management and feeding conditions. The results, adjusted to 305 day predicted values, are presented in Table 10.
The overall average for 305 day predicted milk yields in Table 10 is 9544 kg for Control and 9541 kg for strain 41125 dosed cows, suggesting no benefits carried through to total lactation yields. Three comments are warranted: 1) If peak yields of Farm 1 are observed, the prediction to 305 day production with estimated breeding value as covariate might not have been as accurate as hoped for; 2) If so, it does appear that the response of heifers is variable, but cows primarily benefitted which may be expected, because some effects of SARA are cumulative; 3) The farmers participating in the investigation did not use high energy diets, which the on-station trials (Table 9) showed, are required if cows are to benefit from administration of strain 41125.
3.7.3. Sheep
The utilisation of maize residues by sheep after harvesting is a general practice by farmers in South Africa. Harvesting procedures waste a substantial amount of maize kernels which the sheep prefer in the first 1 - 2 weeks of introduction to the residues, often resulting in ruminal acidosis. Farmers manage the practice by providing hay and supplements before they expose the sheep to the residues, with variable success. The question is whether drenching with strain 41125 instead of adaptation to a concentrate supplement beforehand can prevent acidosis or mitigate the effects. Three on farm trials were conducted with weight gain as indicator; the results are summarized in Table 11.
Drenching with strain 41125 significantly improved weight gains of wethers and post-weaned lambs and reduced weight loss of ewes suckling lambs (Table 11). The suckling lambs apparently also benefitted, even though they were not dosed. In one of the comparisons on Farm 3, weaned lambs were dosed with rumen fluid of sheep adapted to maize feeding (Control) and compared with weaned lambs dosed with strain 41125. The lambs dosed with strain 41125 exhibited higher weight gains (P = 0.02) than the lambs which received rumen fluid. Health of the sheep was also improved: on Farm 1, 2% mortalities and 0.7% morbidities were recorded for Control and zero for both parameters on strain 41125 and on Farm 3, 8.9% mortalities were recorded for ewes on Control and 2.5% for ewes dosed with strain 41125. The results suggest that drenching sheep with strain 41125 before they enter harvested or un-harvested maize fields should be beneficial to health and productivity.
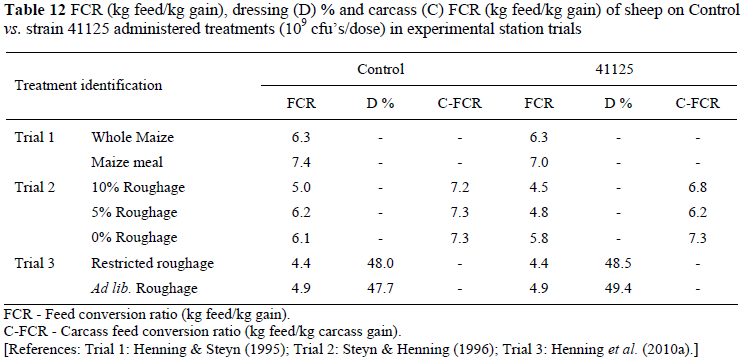
For sheep fed in feedlot simulation at experimental station in trials with small numbers, there is evidence that, as with feedlot cattle, feed efficiency and carcass growth may benefit from strain 41125 drenching by a small margin. The results of Henning & Steyn (1995) (Trial 1), Steyn & Henning (1996) (Trial 2) and Henning et al. (2010a) (Trial 3) at Irene are shown in Table 12.
4. General comments and conclusions
In terms of ability to control lactic acid accumulation in the rumen strain 41125 appears to be more successful than the DFM's cited. It also seems at least on par with in-feed antibiotics that inhibit growth of lactate-producing bacteria, which implies that it could be a worthy substitute once in-feed antibiotics, as in the EU, are banned elsewhere. Adaptation to concentrate diets is enhanced by drenching with strain 41125, which coincides with less roughage required. Production responses in feedlot cattle appear to be of the same order as for other DFM's, but conclusive data are lacking. Milk production is enhanced in the early stages of lactation, but milk composition responses appear to be influenced by TMR composition. Health and production of sheep introduced to maize fields without prior adaptation is improved. In relation to future investigations: 1) The synergism with selected antibiotics and possibly other DFM's provide the opportunity that the benefits of strain 41125 can be exploited further to optimise responses under different environmental and production systems; 2) There may be further production responses with strategic drenching of strain 41125 or in exploiting the long term competitive survival of the organism in the rumen, either in the absence or presence of synergistic products.
References
Aikman, P.C., 2008. Effect of administration of Megasphaera elsdenii NCIMB 41125 lactate utilizing bacteria in early lactation on the production, health and rumen environment of highly productive dairy cows fed a high concentrate diet. CEDAR Report no. 260, University of Reading, UK. pp. 1-100. [ Links ]
Aikman, P.C., Henning, P.H., Horn, C.H. & Humphries, D.J., 2009a. Effects of Megasphaera elsdenii NCIMB 41125 supplementation on rumen fermentation and pH in early lactation dairy cows. ADSA Conf., Abstract no. 33064. [ Links ]
Aikman, P.C., Henning, P.H., Horn, C.H. & Jones, A.K., 2009b. Effect of using Megasphaera elsdenii NCIMB 41125 as a probiotic on feed intake and milk production in early lactation dairy cows. In: Ruminant Physiology - Digestion, Metabolism and Effects of Nutrition on Reproduction and Welfare. Eds Chilliard, Y., Glasser, F., Faulconnier, Y., Bocquier, Y., Veissier, I. & Doreau, M., Wageningen Academic Publishers. pp. 110-111. [ Links ]
Ametaj, B.N., Koenig, K.M., Dunn, S.M., Yang, W.Z., Zebeli, Q. & Beauchemin, K.A., 2009. Backgrounding and finishing diets are associated with inflammatory responses in feedlot steers. J. Anim. Sci. 87, 1314-1320. [ Links ]
Apajalahti, J., 2007. Effect of Megamilk amendment on rumen fermentation kinetics. Research Report: Project KEM0017, Alimetrics Ltd., Espoo, Finland. [ Links ]
Apajalahti, J. & Alaja, S., 2008. Effect of antimicrobial compounds on the growth and metabolism of Megasphaera elsdenii CH4. Research Report: Project KEM0019, Alimetrics Ltd., Espoo, Finland. [ Links ]
Apajalahti, J., Henning, P.H., Horn, C.H., Alaja, S. & Kettunen, A., 2008. Effect of common antimicrobial agents on the growth and metabolism of Megasphaera elsdenii NCIMB 41125, a lactate utilizing probiotic for ruminants. INRA - RRI Symp., Abstract. [ Links ]
ARC/Kemira Phosphates, 2006. "Megasphaera elsdenii strain and its uses". US Patent Application Serial Number 10/521,847, MS Biotech (Pty) Ltd., Centurion, South Africa. [ Links ]
Beauchemin, K. & Penner, G., 2009. New developments in understanding ruminal acidosis in dairy cows. Tri-State Dairy Nutrition Conf., 21-22 April 2009. pp. 1-12. [ Links ]
Boerner, B.J., Byers, F.M., Schelling, G.T., Coppock, C.E. & Green, L.W., 1987. Trona and sodium bicarbonate in beef cattle diets: Effects on pH and volatile fatty acid concentrations. J. Anim. Sci. 65, 309-316. [ Links ]
Brown, M. & Nagaraja, T.G., 2009. Direct-fed microbials for growing and finishing cattle. Plains Nutrition Council Meeting, San Antonio, 10 April 2009. [ Links ]
Burrin, D.G. & Britton, R.A., 1986. Responses to monensin in cattle during subacute acidosis. J. Anim. Sci. 63, 888-893. [ Links ]
Callaway, E.S. & Martin, S.A., 1997. Effects of Saccharomyces cerevisiae culture on ruminal bacteria that utilize lactate and digest cellulose. J. Dairy Sci. 80, 2035-2044. [ Links ]
Chaucheyras, F., Fonty, G., Bertin, G., Salmon, J.M. & Gouet, P., 1996. Effects of a strain of Saccharomyces cerevisiae (levucell SC), a microbial additive for ruminants, on lactate metabolism in vitro. Can. J. Microbiol. 42, 927-933. [ Links ]
Coe, M.L., Nagaraja, T.G., Sun, Y.D., Wallace, N., Towne, E.G., Kemp, K.E. & Hutcheson, J.P., 1999. Effect of virginiamycin on ruminal fermentation in cattle during adaptation to a high concentrate diet and during an induced acidosis. J. Anim. Sci. 77, 2259-2268. [ Links ]
Counotte, G.H.M., Prins, R.A., Janssen, R.H.A.M. & deBie, M.J.A., 1981. Role of Megasphaera elsdenii in the fermentation of DL-[2-13C] lactate in the rumen of dairy cattle. Appl. Environ. Microbiol. 42, 649-655. [ Links ]
Dawson, K.A. & Allison, M.J., 1988. Digestive disorders and nutritional toxicity. In: The rumen microbial ecosystem. Ed. Hobson, P.N., London: Elsevier Applied Science. pp. 445-459. [ Links ]
Devant, M., Garcia, J.A. & Bach, A., 2007. Effect of pre- and postpartum malate supplementation to dairy cows on rumen fermentation and milk production in early lactation. J. Appl. Anim. Res. 31, 169-172. [ Links ]
Donovan, J., 1997. Subacute acidosis is costing us millions. Hoards Dairyman, Sept. 25, 1997. p. 666. [ Links ]
Drouillard, J.S., 2004. Oral dosing of feedlot cattle with Megasphaera elsdenii: impact on adaptation to high-concentrate diets. Research Report: Project no. 2003 - 12, Kansas State University, Manhattan. [ Links ]
Elam, N.A., Gleghorn, J.F., Rivera, J.D., Galyean, M.L., Defoor, P.J., Brashears, M.M. & Younts-Dahl, S.M., 2003. Effects of live cultures of Lactobacillus acidophilis (strains NP45 and NP51) and Propionibacterium freudenreicii on performance, carcass, and intestinal characteristics, and Escherichia coli strain 0157 shedding of finishing beef steers. J. Anim. Sci. 81, 2686-2698. [ Links ]
Emmanuel, D.G.V., Madsen, K.L., Churchill, T.A., Dunn, S.M. & Ametaj, B.N., 2007. Acidosis and lipopolysaccharide from Esherichia coli B: 055 cause hyperpermeability of rumen and colon tissues. J. Dairy Sci. 90, 5552-5557. [ Links ]
Emmanuel, D.G.V., Dunn, S.M. & Ametaj, B.N., 2008. Feeding high proportions of barley grain stimulates an inflammatory response in dairy cows. J. Dairy Sci. 91, 606-614. [ Links ]
Erickson, G.E., Milton, C.T., Fanning, K.C., Cooper, R.J., Swingle, R.S., Parrott, J.C., Vogel, G. & Klopfenstein, T.J., 2003. Interaction between bunk management and monensin concentration on finishing performance, feeding behavior, and ruminal metabolism during an acidosis challenge with feedlot cattle. J. Anim. Sci. 81, 2869-2879. [ Links ]
Godfrey, S.I., Rowe, J.B., Thorniley, G.R., Boyce, G.R. & Speijers, E.J., 1995. Virginiamycin to protect sheep fed wheat, barley or oats from grain poisoning under simulated drought feeding conditions. Aust. J. Agric. Res. 46, 393-401. [ Links ]
Gozho, G.N., Krause, D.O. & Plaizier, J.C., 2006. Rumen lipopolysaccharide and inflammation during grain adaptation and subacute ruminal acidosis in steers. J. Dairy Sci. 89, 4404-4413. [ Links ]
Hagg, F.M., Henning, P.H. & Meissner, H.H., 2009. Effect of Megasphaera elsdenii NCIMB 41125 lactate utilizing bacteria on the rumen environment, production and health of high producing dairy cows. Proc. Livestock Health and Production Group Congress, Mosselbay, South Africa. pp. 48-52. [ Links ]
Hagg, F.M., Erasmus, L.J., Henning, P.H. & Coertze, R.J., 2010. The effect of a probiotic (Megasphaera elsdenii NCIMB 41125) on the productivity and health of Holstein cows. S. Afr. J. Anim. Sci. 40, 101-112. [ Links ]
Hall, M.B. & Averhoff, K.S., 2000. The real costs of digestive upset. Proc. 37th Florida Dairy Production Conf., Gainesville, May, 2-3, 2000. pp. 99-104. [ Links ]
Henning, P.H. & Steyn, D.G., 1995. Efficacy of orally administered CH4 to prevent acidosis and enhance performance of lambs, and optimisation of the dosage size. Kynoch - AECI - ANPI Lactate-utiliser Project: Progress Report, ARC - ANPI, Irene. [ Links ]
Henning, P.H., Campbell, A.A., Hagg, F.M. & Horn, C.H., 2009. Effect of accelerated diet step-up rate on performance of feedlot steers dosed with Megasphaera elsdenii NCIMB 41125. In: Ruminant Physiology - Digestion, Metabolism and Effects of Nutrition on Reproduction and Welfare. Eds Chilliard, Y., Glasser, F., Faulconnier, Y., Bocquier, Y., Veissier, I. & Doreau, M., Wageningen Academic Publishers. pp. 78-79. [ Links ]
Henning, P.H., Horn, C.H., Leeuw, K-J. & Meissner, H.H., 2010a. Effect of ruminal administration of the lactate-utilizing strain Megasphaera elsdenii (Me) NCIMB 41125 on abrupt or gradual transition from forage to concentrate diets. Anim. Feed Sci. Technol. 157, 20-29. [ Links ]
Henning, P.H., Horn, C.H. Steyn, D.G. & Meissner, H.H., 2010b. The potential of Megasphaera elsdenii isolates to control ruminal acidosis. Anim. Feed Sci. Technol. 157, 13-19. [ Links ]
Hino, T., Shimada, K. & Maruyama, T., 1994. Substrate preference in a strain of Megasphaera elsdenii , a ruminal bacterium, and its implications in propionate production and growth competition. Appl. Environ. Microbiol. 60, 1827-1831. [ Links ]
Horn, C.H., Kistner, A. & Fouché, G., 2009a. The Auxostat as isolation and rapid selection tool for strict anaerobic minority organisms in complex ecosystems. Proc. 6th Internat. Symp. Anaerobic Microbiol., Liblice Chateau, Czech Republic, 17-20 June, p. 109 of Abstract book. [ Links ]
Horn, C.H., Kistner, A. & Fouché, G., 2009b. Selective enrichment, isolation and characterization of fast-growing acid-tolerant and ionophores-resistant lactate utilisers from rumen contents of animals on high-energy diets. In: Ruminant Physiology - Digestion, Metabolism and Effects of Nutrition on Reproduction and Welfare. Eds Chilliard, Y., Glasser, F., Faulconnier, Y., Bocquier, F., Veissier, I. & Doreau, M., Wageningen Academic Publishers. pp. 216-217.
Hutjens, M.F., 1999. How and when feed additives may or may not pay. Hoards Dairyman, 25 Sept. 1999. [ Links ]
Iqbal, S., Zebeli, Q., Mazzolari, A., Bertoni, G., Dunn, S.M., Yang, W.Z. & Ametaj, B.N., 2009. Feeding barley grain steeped in lactic acid modulates rumen fermentation patterns and increases milk fat content in dairy cows. J. Dairy Sci. 92, 6023-6032. [ Links ]
Kettunen, A. Henning, P.H., Horn, C.H., Alaja, S. & Apajalahti, J., 2008. Capability of Megasphaera elsdenii strain NCIMB 41125 and other probiotics to prevent accumulation of lactic acid in a rumen model. INRA - RRI Symp., Abstract. [ Links ]
Klieve, A.V., Hennessy, D., Ouwerkerk, D., Forster, R.J., Mackie, R.I. & Attwood, G.T., 2003. Establishing populations of Megasphaera elsdenii YE 34 and Butyrivibrio fibrisolvens YE 44 in the rumen of cattle fed high grain diets. J. Appl. Microbiol. 95, 621-630. [ Links ]
Krehbiel, C.R., Rust, S.R., Zhang, G. & Gilliland, S.E., 2003. Bacterial direct-fed microbials in ruminant diets: Performance response and mode of action. J. Anim. Sci. 81 (E. Suppl. 2), E120-E132. [ Links ]
Kung Jr, L. & Hession, A.O., 1995. Preventing in vitro lactate accumulation in ruminal fermentations by inoculation with Megasphaera elsdenii. J. Anim. Sci. 73, 250-256. [ Links ]
Kung Jr, L., Huber, J.T., Krummerey, J.D., Allison, L. & Cook, R.M., 1982. Influence of adding malic acid to dairy cattle rations on milk production, rumen volatile fatty acids, digestibility, and nitrogen utilization. J. Dairy Sci. 65, 1170-1174. [ Links ]
Leeuw, K-J., Siebrits, F.K., Henning, P.H. & Meissner, H.H., 2009. Effect of Megasphaera elsdenii NCIMB 41125 drenching on health and performance of steers fed high and low roughage diets in the feedlot. S. Afr. J. Anim. Sci. 39, 337-348. [ Links ]
Mackie, R.I. & Gilchrist, F.M.C., 1979. Changes in lactate-producing and lactate-utilizing bacteria in relation to pH in the rumen of sheep during stepwise adaptation to a high-concentrate diet. Appl. Environ. Microbiol. 55, 1570-1573. [ Links ]
Martin, S.A. & Streeter, M.N., 1995. Effect of malate on in vitro mixed ruminal microorganism fermentation. J. Anim. Sci. 73, 2141-2145. [ Links ]
Martin, S.A., Streeter, M.N., Nisbet, D.J. & Williams, S.E., 1999. Effects of DL-malate on ruminal metabolism and performance of cattle fed high-concentrate diets. J. Anim. Sci. 77, 1008-1015. [ Links ]
Marounek, M., Fliegrova, K. & Bartos, S., 1989. Metabolism and some characteristics of ruminal strains of Megasphaera elsdenii. Appl. Environ. Microbiol. 55, 1570-1573. [ Links ]
McDaniel, M.R., Higgens, J.J., Heidenreich, J.M., Shelor, M.K., Parsons, G.L., Henning, P.H. & Drouillard, J.S., 2008. Ruminal parameters of cattle drenched with a placebo, or live cultures of Megasphaera elsdenii strain CH4. Abstract, ASAS-ADSA Annual Congress. [ Links ]
McDaniel, M.R., Heidenreich, J.M., Higgens, J.J., Drouillard, J.S., Henning, P.H. & Horn, C.H., 2009a. Investigation of the effects of a viable strain of Megasphaera elsdenii NCIMB 41125 on ruminal pH and ruminal concentrations of organic acids following a carbohydrate challenge. Chapter 2 in: The effects of dosing cattle with Megasphaera elsdenii NCIMB 41125 prior to the introduction of a grain-rich diet. M.S. thesis of M.R. McDaniel, Kansas State University. pp. 35-76.
McDaniel, M.R., Higgens, J.J., Heidenreich, J.M., Shelor, M.K., Parsons, G.L., Henning, P.H. & Drouillard, J.S., 2009b. Quantitative detection of bacterial genomes and in vitro studies to determine capacity for lactate utilization following intraruminal dosing of cattle with Megasphaera elsdenii strain NCIMB 41125. Ch. 3 in: The effects of dosing cattle with Megasphaera elsdenii NCIMB 41125 prior to the introduction of a grain-rich diet. M.S. Thesis of M R. McDaniel, Kansas State University. pp. 77-97.
Mutsvangwa, T., Walton, J.P., Plaizier, J.C., Duffield, T.F., Bagg, R., Dick, P., Vessie, G. & McBride, B.W., 2002. Effects of a monensin controlled-release capsule or premix on attenuation of subacute ruminal acidosis in dairy cows. J. Dairy Sci. 85, 3454-3461. [ Links ]
Nagaraja, T.G. & Taylor, M.B., 1987. Susceptibility and resistance of ruminal bacteria to antimicrobial feed additives. Appl. Environ. Microbiol. 53, 1620-1625. [ Links ]
Nagaraja, T.G. & Titgemeyer, E.C., 2007. Ruminal acidosis in beef cattle: the current microbiological and nutritional outlook. J. Dairy Sci. 90 (E. Suppl.): E17-E38. [ Links ]
Nagaraja, T.G., Avery, T.B., Bartley, E.E., Roof, S.K. & Dayton, A.D., 1982. Effect of lasalocid, monensin or thiopeptin on lactic acidosis in cattle. J. Anim. Sci. 54, 649-658. [ Links ]
Newbold, C.J. & Wallace, R.J., 1988. Effects of the ionophores monensin and tetronasin on simulated development of ruminal lactic acidosis in vitro. Appl. Environ. Microbiol. 54, 2981-2985. [ Links ]
Nisbet, D.J. & Martin, S.A., 1993. Effects of fumarate, L-malate, and an Aspergillus oryzae fermentation extract on D-lactate utilization by the ruminal bacterium Selenomonas ruminantium. Curr. Microbiol. 26, 133. [ Links ]
Nocek, J.E., 1997. Bovine acidosis: Implication on laminitis. J. Dairy Sci. 80, 1005-1028. [ Links ]
Nocek, J.E., Kautz, W.P., Leedle, J.A.Z. & Allman, J.G., 2002. Ruminal supplementation of direct-fed microbials on diurnal pH variation and in situ digestion in dairy cattle. J. Dairy Sci. 85, 429-433. [ Links ]
Oetzel, G.R., 2003. Subacute ruminal acidosis in dairy cattle. Advances in Dairy Cattle 15, 307-317. [ Links ]
Owens, F.N., Secrist, D.S., Hill, W.J. & Gill, D.R., 1998. Acidosis in cattle: A review. J. Anim. Sci. 76, 275-286. [ Links ]
Penner, G.B., Beauchemin, K.A. & Mutsvangwa, T., 2007. Severity of ruminal acidosis in primiparous Holstein cows during the periparturient period. J. Dairy Sci. 90, 365-375. [ Links ]
Plaizier, J.C., Krause, D.O., Gozho, G.N. & McBride, B.W., 2008. Subacute ruminal acidosis in dairy cows: the physiological causes, incidence and consequences. Vet. J. 176, 21-31. [ Links ]
Raeth-Knight, M.L., Linn, J.G. & Jung, H.G., 2007. Effect of direct-fed microbials on performance, diet digestibility, and rumen characteristics of Holstein dairy cows. J. Dairy Sci. 90, 1802-1809. [ Links ]
Rinttilä, T., Kettunen, A. & Apajalahti, J., 2008. Development of CH4 strain-specific real-time PCR essay. Alimetrics Ltd., Espoo, Finland. [ Links ]
Rinttilä, T., Ristilä, R. & Apajalahti, J., 2009. Effect of four prophylactic antibiotics on the growth and metabolism of Megasphaera elsdenii CH4. Research Report: Project KEM0023. Alimetrics Ltd., Espoo, Finland. [ Links ]
Robinson, J.A., Smolenski, W.J., Greening, R.C., Ogilvie, M.L., Bell, R.L., Barsuhn, K. & Peters, J.P., 1992. Prevention of acute acidosis and enhancement of feed intake in the bovine by Megasphaera elsdenii 407A. J. Anim. Sci. 70, 310. (Abstract). [ Links ]
Rogosa, M., 1984. Anaerobic gram-negative cocci. In: Bergey's Manual of Systematic Bacteriology. Vol. 1. Eds Krieg, N.R. & Holt, J.G., Baltimore/London: Williams & Wilkins. pp. 680-685. [ Links ]
Russell, J.B. & Baldwin, R.L., 1978. Substrate preferences in rumen bacteria: Evidence of catabolite regulatory mechanisms. Appl. Environ. Microbiol. 36, 319-329. [ Links ]
Slyter, L.L., 1976. Influence of acidosis on rumen function. J. Anim. Sci. 43, 910-929. [ Links ]
Steyn, D.G., 1996. Efficacy of the lactate-utilizing rumen bacteria, Megasphaera elsdenii, in preventing rumen acidosis in lambs, in the absence of ionophores. Treatise, B.Tech. Agric. (Anim. Prod.), Technikon, Pretoria. [ Links ]
Steyn, D.G. & Henning, P.H., 1996. Evaluation of CH4 efficacy in lambs on concentrate diets with decreasing levels of roughage inclusion. Research Report on Project: Isolation, selection and cultivation of lactate-utilizing rumen bacteria for the treatment of chronic and acute lactic acidosis. ARC-ANPI, Irene, South Africa. [ Links ]
Stock, R. & Britton, R., 2006. Acidosis. In: Beef Cattle Handbook, Communication BCH-3500. University of Nebraska, USA. [ Links ]
Stokes, M.R., Vanoemark, L.L. & Bull, L.S., 1986. Effects of sodium bicarbonate, magnesium oxide, and a commercial buffer mixture in early lactation cows fed hay crop silage. J. Dairy Sci. 69, 1595-1603. [ Links ]
Stone, W.C., 2004. Nutritional approaches to minimize subacute ruminal acidosis and laminitis in dairy cattle. J. Dairy Sci. 87 (E. Suppl.), E13-E26. [ Links ]
Thorniley, G.R., Rowe, J.B., Cowcher, P.C. & Boyce, M.D., 1998. A single drench of virginiamycin to increase safety of feeding grain to sheep. Aust. J. Agric. Res. 49, 899-906. [ Links ]
Waldrip, H.M. & Martin, S.A., 1993. Effects of an Aspergillus oryzae fermentation extract and other factors on lactate utilization by the ruminal bacterium Megasphaera elsdenii. J. Anim. Sci. 71, 2770-2776. [ Links ]
Wiederhold, A.H., 1994. Isolation, selection and cultivation of lactic acid-utilizing rumen bacteria for the treatment of chronic and acute acidosis. MSc. thesis. University of Natal, Pietermaritzburg, RSA. [ Links ]
Wiryawan, K.G. & Brooker, J.D., 1995. Probiotic control of lactate accumulation in acutely grain-fed sheep. Aust. J. Agric. Res. 46, 1555-1568. [ Links ]
Yoon, I.K. & Stern, M.D., 1995. Influence of direct-fed microbials on ruminal microbial fermentation and performance of ruminants: A review. Asian-Australas. J. Anim. Sci. 8, 533-555. [ Links ]
Zebeli, Q., Dijkstra, J., Tafaj, M., Steingass, H., Ametaj, B.N. & Drochner, W., 2008. Modeling the adequacy of dietary fiber in dairy cows, based on the responses of ruminal pH and milk fat production to composition of the diet. J. Dairy Sci. 91, 2046-2066. [ Links ]
Zinn, R.A. & Borques, J.L., 1993. Influence of sodium bicarbonate and monensin on utilization of a fat-supplemented, high-energy growing-finishing diet by feedlot steers. J. Anim. Sci. 71, 18-25. [ Links ]
* Corresponding author. E-mail: heinzmeissner@vodamail.co.za
** Present address: 189 van Riebeeck Avenue, Centurion 0157, South Africa
*** Present address: Biotherapeutics, PO Box 207, Irene 0062, South Africa


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}