










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.5 Pretoria Jan. 2009
Volatile fatty acids in cattle supplemented with protein-enriched salt and sodium monensin
G.B. Neto#; A. Berndt; J.R. Nogueira; J.C. Nogueira; J.J.A.A. Demarchi
Agência Paulista de Tecnologia dos Agronegócios, São Paulo, Brasil
ABSTRACT
The effects of sodium monensin and protein supplements on volatile fatty acid production were studied in a 4 x 4 Latin Square design, using four female bovines equipped with rumen fistulae and fed Brachiaria brizantha Marandu cultivar hay. The treatments were organized in a 2 X 2 factorial array (with or without 160 mg of sodium monensin/animal/day and with or without protein supplements). Protein supplementation increased the concentration of acetic acid and sodium monensin reduced the concentrations of acetic and butyric acids and the acetic : proprionic ratio, both with and without protein supplements, except for the material collected at 8 h. Sodium monensin reduced the total concentration of fatty acids only in the material collected at 0 h, while protein supplementation increased the concentration of fatty acids at 4, 6 and 8 h. Used together, they increased the molar concentration of propionate in the rumen fluid, without decreasing the total concentration of fatty acids.
Keywords: Marandu grass, ionophore
Introduction
Adding ionophores to mineral salt can increase the efficiency of energy metabolism in the rumen or organism, thus increasing efficiency of animal production. The mechanism that leads to improved performance when ionophores are used is not yet completely understood. However, it is known that the metabolism of bacteria in the rumen is affected by its toxic effect on microorganisms, resulting in alterations in the molar concentration of fatty acids. The toxicity of these compounds arises from their capacity to penetrate into biological membranes and subsequently alter the flux of ions from and into the cell (Ipharraguerre & Clark, 2003). According to Chalupa (1977) the effect that monensin in increasing propionate and decreasing acetate and methane is advantageous metabolically, since propionate production is energetically more efficient. On the other hand Oliveira et al. (2005) showed a reduction of the voluntary feed intake in cattle fed monensin, and Dye et al. (1988) observed that adding ionophores to the diet linearly reduced the total fatty acid levels. However, the response to the use of ionophores varies due to several factors, including the concentration and quality of fibre in the diet, the amount and quality of the nitrogen source, the period of adaptation to the product, the time when the sample was collected, the type and concentration of the ionophore, as well as the presence of fat, buffers, etc. One of the options to reduce the seasonality impact in the production of forage for feeding bovine is closing the pasture at the end of the growth period to accumulate forage to supply the animals in the drought period. This technique makes forage with a low nutritive value available due to the advance in the physiological age of the plant. Tropical grasses generally have low protein levels and a high proportion of cell wall components. Milford & Minson (1965) showed that the voluntary intake was positively correlated with the crude protein content when less than 7% in the dry matter. At this level the voluntary intake of dry matter by ruminants could be limited by a lack of crude protein. However, the protein intake can be increased by supplementing the diet with protein or non-protein nitrogen. In this situation, protein-enriched salt associated with ionophores can result in the intake of a larger amount of dry matter and increased efficiency in its digestion. The aim of this study was to examine the effects of sodium monensin and protein supplementation on the concentration of fatty acids in bovines fed Brachiaria brizantha Marandu cultivar hay.
Material and Methods
This experiment was conducted at the Agência Paulista de Tecnologia dos Agronegócios, São Paulo, Brasil. Four dry, mixed-breed cows, which had a mean live weight of ca. 500 kg, had all been fitted with rumen fistula from the Kehl Industry and Trade LTDA - Italo Paino street, 700, zipcode 13564-610, city São Carlos, State São Paulo, Brazil. These cows were fed exclusively with Brachiaria brizantha Marandu cultivar hay, along with the supplements: mineral salt, protein-enriched salt, mineral salt with monensin or protein-enriched salt plus monensin. The concentrations (g/kg) of crude protein, ether extract, ash, acid detergent fibre, neutral detergent fibre, cellulose, lignin and hemicellulose in dried hay were 39, 9, 68, 495, 804, 407, 63 and 309, respectively. The treatment without protein supplementation and without monensin consisted of supplementation with 48 g of a mineral salt alone. Whenever the interaction was not significant, the probability of main effects was used, which were monensin (with or without) and protein-enriched salt (with or without). All the treatments were administered daily into the rumen through the fistulae at 8:00.
The protein supplementation was based on the requirements (g/kg) of 78.6, 510, 2.5 and 1.5 of crude protein, total digestible nutrients, Ca and P, respectively (NRC, 1996). The proteinated salt was supplied at a rate of 250 g/day. The concentration (g/kg) of dry matter, crude protein, TDN, ether extract, crude fibre, Ca and P in the proteinated salt were 953.4, 1122.4, 431.7, 23.8, 17.2, 23.8 and 20.4, respectively, and was composed of soyabean meal, protenose, urea and mineral salt at percentages of 25.8, 25.0, 30.0 and 19.2, respectively. The monensin was supplied at a rate of 160 mg/day by introducing 32 g of Bovicorte Premium® mineral (NUTRACT), which contains 0.5% sodium monensin.
The experiment was conducted during four periods of 25 days each; the first 19 days of each trial was for the animals to adapt to the diets. During the 20th day, rumen fluid was collected 0, 1, 2, 3, 4, 6 and 8 h after morning feeding at 8:00. The collection at 0 h was done immediately before the animals were treated. The fatty acids were measured with gas chromatography, according to Erwin et al. (1961).
A 4 x 4 Latin square in a 2 x 2 factorial design was used, with four animals and four treatments (with or without monensin and with or without protein supplements). These analyses separate the effects of monensin, of protein supplements, and the interaction between these factors. Whenever the interaction was significant (P <0.05), the effects of monensin within protein supplementation and vice versa were tested through orthogonal contrasts. Whenever the interaction was not significant, the probability of main effects was used. In the first contrast, there was a separate monensin effect (0 vs. 160 mg monensin/animal/day). In the second contrast, there was a separate protein-enriched salt effect (0 vs. 250 g protein-enriched salt/animal/day). The statistical analyses were done using the Statistical Analysis System (SAS, 2003) It was used in the CONTRAST command of PROC GLM (Nogueira, 2004).
Results and Discussion
Monensin and protein supplementation affected the concentration of acetic acid. The monensin reduced and the protein supplementation increased acetate concentration at four of the seven sampling times (P <0.05) (Table 1).
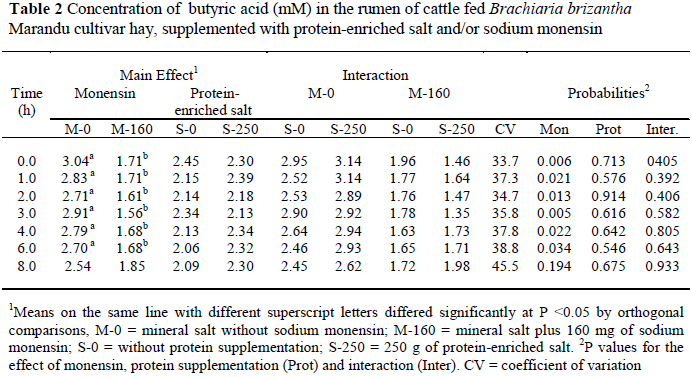
Sodium monensin reduced the concentration of butyric acid at six of the seven sampling times (P < 0.05); protein supplementation, however, had no effect on butyric acid concentration (P > 0.05) (Table 2).
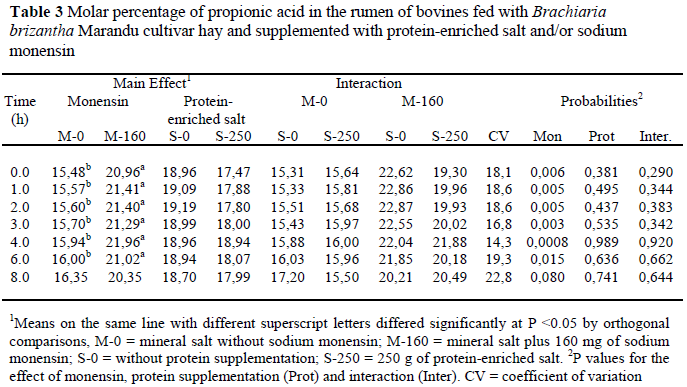
There was a tendency (P = 0.06) for protein supplementation to increase the concentration of proprionic acid. The propionate levels, with and without protein supplements, were 9.39 and 11.86 mM, respectively, 4 h after feeding. Monensin increased the molar percentage of propionic acid (Table 3).
Monensin significantly (P <0.05) reduced the concentration of fatty acids in the collection made at 0 h (68 vs. 58 mM) and also reduced (P <0.05) the acetic : propionic acid ratio from 5.2 to 3.8. Protein supplementation increased (P <0.05) the total fatty acid concentration from a mean of 48 to 62 mM in the collections made at 4, 6 and 8 h without significantly altering the acetic : propionic ratio. The low level of CP (39 g/kg) harms the microbial growth and it reduces in situ digestibility. There was significant (P <0.05) interaction between protein-enriched salt and monensin on the forage dry matter in situ digestibility with 96 h of incubation. While the monensin reduced the digestibility (43.7 vs. 30.8%), in diets with monensin, the protein-enriched salt increased the digestibility (26.1 vs. 35.6%). The increase of the digestibility by protein-enriched salt caused an increased fatty acids production. However, the monensin did not stop exerting an effect on the fatty acids proportion. Yang & Russel (1993) also observed that monensin increased propionate and reduced acetate concentrations, both with and without protein supplementation. According to Chen & Wolin (1978), these effects are due to the inhibitory action on the bacteria that produce formate and hydrogen, while they stimulate those that produce succinate and propionate; which has double the advantage metabolically, since the production of propionate is energetically more efficient than the production of acetate (Chalupa, 1977). We found that monensin altered the proportion between acetic, propionic and butyric acids from 80 : 16 : 4 to 75 : 22 : 3, respectively. Richardson et al. (1976) observed an increase in energy due to change in molar concentration from 60 : 30 : 10 (acetic, propionic and butyric acids) to 52 : 40 : 8 of 5.6% in crude energy in the form of fatty acids, since the theoretical conversion of energy contained in a hexose (2.81 MJ/mole) to energy contained in acetic, propionic and butyric acids corresponds to 1.76, 3.07 and 2.19 MJ/mole, respectively, in fermented hexose. Based on this conversion proposed by Richardson et al. (1976), the change in the proportion of the fatty acids that we observed would result in an increase of 3.7% of crude energy in the form of fatty acids, both with and without protein supplementation. This is similar to what was found by Chalupa et al. (1980) who observed a 2 to 6% increase in the fermentation efficiency as a function of the increase in propionate and the decrease in acetate, and by Whetstone et al. (1981), who found that the changes in the fatty acid ratios increased fermentation efficiency from 1.8 to 5.6%. Raun et al. (1976) found an increase of 3 to 6% in metabolizable energy, due to the alteration in the molar proportion of fatty acids, resulting in weight gains of up to 11% and increased feed conversion efficiency.
Conclusions
Sodium monensin increased the concentration of propionic acid in the rumen. Protein supplements increased fatty acid concentrations, without impeding the effect that sodium monensin has on reducing the acetic acid : propionic acid ratio. Adding these two supplements together increased the molar concentration of propionate in the rumen fluid without reducing the total fatty acid concentration.
References
Chalupa, W., 1977. Manipulating rumen fermentation. J. Anim. Sci. 45, 585-599. [ Links ]
Chalupa, W., 1980. Chemical control of rumen microbial metabolism. Digestive Physiology and Metabolism in Ruminants, AVI Publishing Co., Inc., Westport, CT. p. 325 [ Links ]
Chen, M. & Wolin, M.F., 1978. Effect of monensin and lasalocid on the growth of rumen and methane bacteria. Am. Soc. Microbiol., 78th Ann. Meet. (Abstr.). p. 88. [ Links ]
Dye, B.E., Amos, H.E. & Froetschel, M.A., 1988. Influence of lasalocid on rumen metabolites, milk production, milk composition and digestibility in lactating cows. Nutr. Rep. Int. 38, 101-115. [ Links ]
Erwin, E.S., Marco, G.J. & Emery, E.M., 1961. Volatile fatty acid analyses of blood and rumen fluid by gas chromatography. J. Dairy Sci. 44, 1768-1771. [ Links ]
Ipharraguerre, I.R. & Clark, J.H., 2003. Usefulness of ionophores for lactating dairy cows: A review. Anim. Feed Sci. Technol. 106, 39-57. [ Links ]
Milford, R. & Minson, S.J., 1965. The relation between the crude protein content of tropical pasture plants. J. Br. Grassl. Soc. 20, n.3, pp. 1977-1979. [ Links ]
Nogueira, M.C.S., 2004. Orthogonal contrasts: definitions and concepts. Sci. Agric. 61, 118-124. [ Links ]
NRC, 1996. Nutrient Requirements of Beef Cattle (7th ed). National Academy Press, Washington D.C., USA. [ Links ]
Oliveira, M.V.M., Lana, R.P., Jham, G.N., Peereira, J.C., Pérez, J.R.O. & Valadares, S.C.F., 2005. Effects of different dietary levels of monensin and protein on intake and ruminal fermentation in bovines. Braz. J. Anim. Sci. 34, 1763-1774. [ Links ]
Raun, A.P., Cooley, C.O., Potter, E.L., Rathmather, R.P. & Richardson, L.F., 1976. Effect of monensin on feed efficiency of feedlot cattle. J. Anim. Sci. 43, 670-677. [ Links ]
Richardson, L.F., Raun, A.P., Potter, E.L. & Cooley, C.O., 1976. Effect of monensin in rumen fermentation in vitro and in vivo. J. Anim. Sci. 43, 657-664. [ Links ]
SAS, 2003. Statistical Analysis Systems user's guide. SAS Institute Inc., Raleigh, North Carolina, USA. [ Links ]
Whetstone, H.D., Davis, C.L. & Bryant, M.P., 1981. Effect of monensin on breakdown of protein by ruminal microorganisms in vitro. J. Anim. Sci. 53, 803-809. [ Links ]
Yang, C.M.J. & Russel, J.B., 1993. The effect of monensin supplementation on ruminal ammonia accumulation in vivo and the numbers of amino acid-fermenting bacteria. J. Anim. Sci. 71, 3470-3476. [ Links ]
# Corresponding author. E-mail: geraldobalieiro@apta.sp.gov.br


{kind=link}
{kind=link}
{kind=link}