










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.5 Pretoria Jan. 2009
Monensin and protein supplements on methane production and rumen protozoa in bovine fed low quality forage
G.B. Neto#; A. Berndt; J.R. Nogueira; J.J.A.A. Demarchi; J.C. Nogueira
São Paulo Agribusiness Technology Agency, São Paulo, Brazil
ABSTRACT
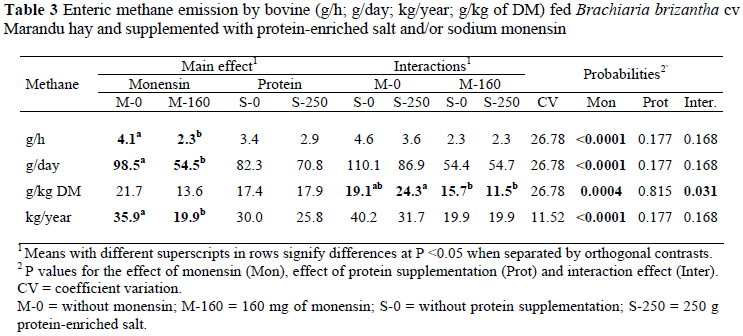
This study aimed at evaluating the effect of sodium monensin and protein supplementation in the enteric methane production and the population of ruminal protozoa in bovine fed Brachiaria brizantha Marandu cultivar hay. The treatments were: mineralized salt, mineralized salt with monensin, protein-enriched salt and protein-enriched salt with monensin. The experimental outline was in a 4 x 4 Latin square. The protein supplementation increased the dry matter intake and the total concentrations of rumen ciliate protozoa. Monensin eliminated the rumen fauna and reduced the methane production. Methane production was 19.13; 15.73; 24.35 and 11.52 g/kg of ingested DM when mineralized salt, mineralized salt with monensin, protein-enriched salt and protein-enriched salt with monensin were supplied, respectively. The association between the inputs allows for a reduction in the methane production without a reduction in dry matter intake.
Keywords: Marandu hay, enteric methane, ionophore, ciliate protozoa, SF6 tracer technique
Introduction
One of the options to reduce the seasonality impact in the production of forage for feeding bovine is closing the pasture at the end of the growth period to accumulate forage to supply the animals in the drought period. This technique makes forage with a low nutritive value available due to the advance in the physiological age of the plant. In this situation, protein supplementation associated with ionophores can result in the intake of a larger amount of dry matter and increased efficiency in its digestion. The inclusion of ionophores in mineralized salt may change the fermentation pattern and increase the energy use of the diet.
The exact mechanism that leads to the improved performance when using ionophores has not been fully clarified yet. Ionophores are substances obtained through the fermentation by bacteria from the Streptomyces cinnamonensis group, being highly lipophilic and toxic to many microorganisms and are commonly used as coccidiostat drugs in poultry production. The toxicity of the ionophores to determined microorganisms can reduce the energetic loss of the diet through enteric methane production. Wedegaertner & Johnson (1983) observed that the effect of monensin in the reduction of the methane production and the loss of energy in the faeces and urine increased the retention of energy in bovines by 19%. Moreover, the methane expelled by the ruminants is presently the focus of criticism, since it presents a greenhouse effect 21 times greater than CO2 (UNFCC, 2005), a lifespan of 14 years in the atmosphere (IPCC, 1995) and an annual emission estimate, of the Brazilian herd alone, of 9.38 Tg of CH4/year (MCT, 2006).
The present study aimed at evaluating the effects of sodium monensin and protein supplementation in the enteric methane production and the population of ruminative ciliated protozoa in bovine fed with Brachiaria brizantha Marandu cultivar hay.
Material and Method
The experiment was conducted at the São Paulo Agribusiness Technology Agency, São Paulo, Brazil. Four dry, cross-bred bovine females with an average live weight of 500 kg, and fitted with ruminal cannulae were used. The animals were exclusively fed with Brachiaria brizantha Marandu cultivar hay plus the four treatments: mineralized salt, protein-enriched salt, mineralized salt with monensin and protein-enriched salt with monensin. The concentration of crude protein, ether extract, minerals, acid detergent fibre, neutral detergent fibre, cellulose, lignin and hemicellulose in dried hay were 39, 9, 68, 495, 804, 407, 63 and 309 g/kg, respectively. The control received only 48 g of the mineral salt with the basic diet. All the treatments were given daily at 08:00 through the fistula directly into the rumen.
The protein supplementation was based on NRC requirements (NRC, 1996) of 78.6 g CP; 510 g total digestible nutrients (TDN); 2.5 g Ca and 1.5 g P/kg feed, and was supplied at 250 g/day. The composition of the protein supplement was 95.3% DM, and 112.2 CP, 431.7 g TDN, 23.8 g EE, 17.2 g crude fibre, 23 8 g Ca and 20.4 g P/kg and consisted of soya meal, protenosis, urea and a mineralized salt at proportions of 25.8, 25.0, 30.0 and 19.2%, respectively. The sodium monensin was supplied at a rate of 160 mg/day by introducing 32 g of Bovicorte Premium® mineral (NUTRACT), which contains 0.5% sodium monensin.
The experiment was performed in four, 25-day periods, and the first 19 days of each period focused on the adaptation to the diet. During the 20th day, rumen fluid samples were collected for the rumen ciliate protozoa count at 0, 1, 2, 3, 4, 6 and 8 hours after the morning feeding, done at 8:00. The 0-hour collection was performed immediately before the animals received the treatment. Between the 21st and 25th days, gas samples were collected to evaluate the methane production according to Johnson & Johnson (1995). The methane and SF6 concentrations were determined using gas chromatography. The dry matter intake was evaluated daily throughout the adaptation phase and entire experiment period.
The animals were designed into a 4 x 4 Latin square within a 2 x 2 factorial arrangement, with four animals and four treatments (with or without monensin and with or without protein supplementation). The analyses separate the effect of the monensin, the effect of the protein supplementation and the interaction between these factors as variation sources. Whenever the interaction was significant (P <0.05), the effects of the monensin within the protein supplementation and vice versa were tested through orthogonal contrasts. Whenever the interaction was not significant, the probability of the main effects was used.
Results and Discussion
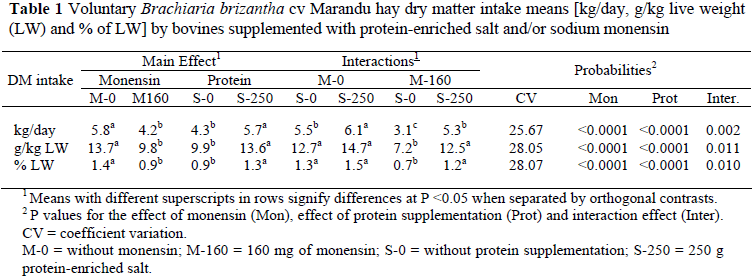
The low nutritive value of the forage resulted in low DM intake, 5.0 kg/day in average. There was significant interaction between the effects of the protein supplementation and monensin on the voluntary DM intake (Table 1). The protein supplementation did not allow the monensin to reduce the DM intake in g/kg of live weight (LW) and in percentage of the live weight (%LW) (Table 1).
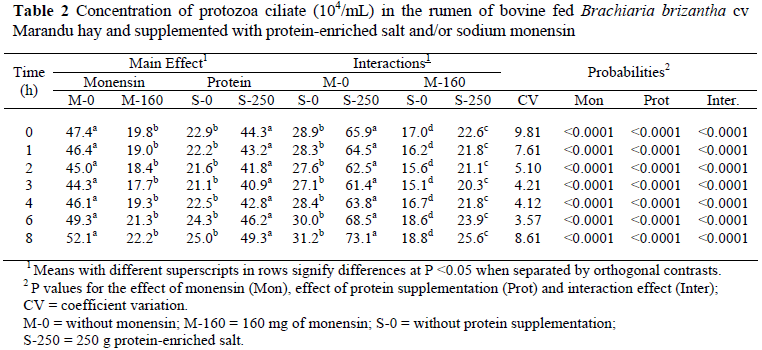
There was a significant interaction between the effects of monensin and the protein supplementation on the population of ciliate protozoa in the rumen (Table 2). Monensin reduced the ciliate protozoa in the rumen and the protein supplementation increased the same, nonetheless, without avoiding the reduction caused by monensin (Table 2). Guan et al. (2006) also observed a reduction in the ciliate protozoa count when they supplied monensin to the animals.
The protozoa can use most of the carbohydrates for growth and this process produces hydrogen. The hydrogen produced by the ciliate protozoa is used by methanogenic Archaea that live in symbiosis with the protozoa and are responsible for the formation of methane. Thus, the protozoa can be responsible for up to 37% of methanogenesis (Williams & Coleman, 1997) and their elimination would result in the reduction of the methane production in the rumen.
The mean of six studies has indicated that monensin can reduce methane emissions by 25%, and varied between 4% and 31% (Schelling, 1984). In the present study, monensin reduced methane production by 44% (P <0.001) compared to the treatments without monensin (Table 3). However, 55% of the methane emission reduction caused by monensin is due to the reduction in DM intake and only 45% is due to specific effects on ruminal fermentation (O'Kelly & Spiers, 1992). Analyzing the methane emissions in g/kg of DM ingested, a significant interaction (P <0.05) was observed between the effects of monensin and protein supplementation. The reduction in the methane production, in g/kg of DM, caused by monensin ingested was 52% with protein supplementation and 17% without protein supplementation, demonstrating a positive association of the inputs. The annual emission of 40 kg of methane in the control treatment was below the value of 56 kg mentioned by the Intergovernmental Panel Climate Change (IPCC, 2006). Nevertheless, methane production in the month of August reported by Demarchi et al. (2003) for animals maintained in Brachiaria brizantha cv Marandu pastures was of 102 g/day, in agreement with the value of 110.2 g/day observed in the control treatment (Table 3). The authors attributed the lower emission to the low nutritive value of the forage for that time of year. The ciliate protozoa in the rumen population was positively correlated with methane production (r = 65%, P <0.05), a result coherent with the monensin effect in the elimination of ciliate protozoa from the rumen and reduction in methane production.
Conclusions
Monensin reduces methane production and eliminates the ciliate protozoa from the rumen. The protein supplementation increases the protozoa ciliate population and avoids the reduction in dry matter intake caused by monensin. The association between the inputs allows for a reduction in methane production without a reduction in dry matter intake.
References
Demarchi, J.J.A.A., Manella, M.Q. & Lourenço, A.J., 2003. Daily methane emission at different seasons of the year by Nellore cattle in Brazil grazing Brachiaria brizantha cv. Marandu. In: IX WCAP, 2003, Porto Alegre - RS, Brazil. Proceedings, Porto Alegre: WAAP, 3 p. [ Links ]
Guan, H., Wittenberg, K.M. & Ominski, K.H., 2006. Efficacy of ionophores in cattle diets for mitigation of enteric methane. J. Anim. Sci. 84, 1896-1906. [ Links ]
Intergovernmental Panel on Climate Change, 1995. Climate Change: Radiative Forcing of Climate Change and an Evaluation of the IPCC IS92 Emission Scenarios. Cambridge: University Press. 339 pp. [ Links ]
Intergovernmental Panel on Climate Change, 2006. Guidelines for National Greenhouse Gas Inventories. Chapter 10: Emissions from livestock and Manure Management. [ Links ]
Johnson, K.A. & Johnson, D.E., 1995. Methane emissions from cattle. J. Anim. Sci. 73, 2483-2492. [ Links ]
MCT - Ministério da Ciência e Tecnologia, 2006. Primeiro Inventário Brasileiro de Emissões Antrópicas de Gases de Efeito Estufa - Emissões de Metano pela Pecuária, Brasília, Brasil. [ Links ]
NRC, 1996. Nutrient Requirements of Beef Cattle (7th ed.). National Academy Press, Washington D.C., USA. [ Links ]
O'Kelly, J.C. & Spiers, W.G., 1992. Effect of monensin on methane and heat productions of steers fed lucerne hay either ad libitum or at the rate of 250 g/h. Aust. J. Agric. Res. 43, 1789-1793. [ Links ]
Schelling, G.T., 1984. Monensin mode of action in the rumen. J. Anim. Sci. 58, 1518-1527. [ Links ]
UNFCCC. Key GHG Data. Bonn: UNFCCC. 2005. [ Links ]
Wedegaertner, T.C. & Johnson, D.E., 1983. Monensin effects on digestion, methanogenesis and heat increment of a cracked corn-silage diet fed to steers. J. Anim. Sci. 57, 168-177. [ Links ]
Williams, A.G. & Coleman, G.S., 1997. The rumen protozoa. In: The Rumen Microbial Ecosystem. Eds Hobson, P.N. & Stewart, C.S., Blackie Academic, London, UK. 2, 73-139. [ Links ]
# Corresponding author. E-mail: geraldobalieiro@apta.sp.gov.br


{kind=link}
{kind=link}
{kind=link}