










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.5 Pretoria Jan. 2009
Modelling the long-term consequences of undernutrition of cows grazing semi-arid range for the growth of their progeny
F.D. Richardson#
Department of Mathematics and Applied Mathematics, University of Cape Town, Private Bag X3, Rondebosch 7701, South Africa
ABSTRACT
Undernutrition of cows grazing semi-arid range as a result of drought or heavy stocking rates reduces prenatal and pre-weaning growth of their calves. A mechanistic model has been developed that simulates the productivity of vegetation and cattle on semi-arid savanna rangeland in southern Africa. When cows are stocked at 0.200 cows/ha instead of 0.123 cows/ha the model predicted that empty body weights of their progeny at birth and weaning would be reduced by 2.9 and 43.8 kg, respectively. If all young animals are stocked at 0.22 animals/ha after weaning, differences in empty body weight at weaning are predicted to persist almost unchanged for a further two years. The model also predicted that undernutrition of cattle aged 21 months would be followed by partial compensatory growth. Comparison of simulated results with data from experiments in Zimbabwe indicates that the model realistically simulates the effects of stocking rate on cattle growth.
Keywords: Semi-arid range, mechanistic model, cow nutrition, calf feeding, compensatory growth
Introduction
Cattle grazing semi-arid range may be subjected to low intakes of metabolisable energy and protein as a result of drought, high stocking rates, or during the dry season low digestibility and low protein content of the forage (Butterworth, 1984). The growth and milk yield of cattle are reduced during such periods of undernutrition. Growing animals respond in different ways to nutritional restriction because of differences in age at the start of undernutrition, the severity and duration of undernutrition and the availability and composition of food during rehabilitation (O'Donovan, 1984). Severe chronic undernutrition of cattle in early life reduces growth and leads to smaller animals at any age (Morgan, 1972; Greenwood & Café, 2007). Undernutrition of breeding cows during the last trimester of gestation leads to a reduction in birth weight (Richardson et al., 1979) and in the cow's milk yield (Richardson et al., 1977) as a result of the effect of undernutrition on udder development (Mellor & Murray, 1985) so that pre-weaning growth is also reduced. On the other hand, compensatory growth may occur in rangeland cattle subjected to undernutrition after weaning. For example, Ainslie (1958) reported that when steers were subjected to different planes of nutrition between weaning at 8 - 9 months of age and 12 months the difference of 23 kg between group means was eliminated when all animals grazed together without being given any supplementary food. In this paper a mechanistic model of a rangeland production system has been used to explain the different responses to retardation of growth that have been recorded in the literature.
Materials and Methods
A comprehensive mechanistic model of forage and individual animals and the interactions between them has been developed using published information and established theory (Richardson et al., 2000). The model partitions daily rainfall between runoff, infiltration and drainage and also simulates the loss of soil moisture by evaporation and transpiration. Forage production is modelled in relation to soil moisture and the present potential for growth. Prediction of metabolisable energy intake is an important component of the model. The simulated intake of growing animals is the smallest of those predicted by three different mechanisms (Poppi et al., 1994). These are the rate at which the animal is able to harvest forage, the physical capacity of the digestive system and the animal's potential for growth. The rate of harvesting depends on the breadth of the dental arcade and the maximum number of bites per day which increases from zero in calves less than seven days old to a maximum of 38 000/day at 16 weeks. The rate of harvesting of forage is also dependent on forage density assuming a sigmoid response to calculate an adjustment factor, Y, Equation 1 (Thornley, 1998).

Where X is forage density, kg/ha; k is the forage density at which intake is half of maximum potential and q is a steepness parameter. In the model the upper limit to voluntary intake imposed by the capacity of the digestive system, Ids, is determined from faecal DM output, FDM,, and the metabolisability of the diet, Qd,Equation 2. The factor 0.82 is the ratio between metabolisability and digestibility of forage diets (Kahn & Spedding, 1984).

Dental arcade breadth in continuously growing animals increases with live weight and mature weight, Figure 1, but does not decrease if the animal loses weight. Digestive system capacity is measured as potential faecal DM output that is related to live weight in animals that are gaining weight (Figure 1) (Butterfield, 1988). Faecal output reaches a maximum when live weight is 0.75 of mature weight. If live weight increases beyond 0.75 of that at maturity then the capacity of the digestive system decreases due to the accumulation of fat in the body cavity. This decrease is reversible if the animal loses weight. Loss in weight in animals less than 0.75 of mature weight does not lead to decline in faecal output.
Metabolisable energy intake is partitioned between maintenance, accretion/depletion of body protein and fat, conceptus growth and milk production, using the method of Finlayson et al. (1994). The model simulates the effect of nutrition on udder development during the final 100 days of pregnancy as this has a major effect on subsequent milk yield. In this paper the model is used to study the effects of undernutrition of breeding cows on the prenatal, pre-weaning and post-weaning growth of their calves.
Results and Discussion
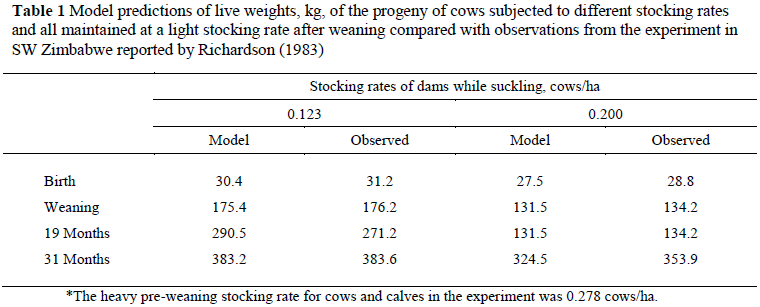
The model was evaluated by comparing model output with the results of an experiment in South Western Zimbabwe that examined the performance of individual cows subjected to widely different stocking rates, 0.123 (LSR) or 0.278 (HSR) cows/ha, during successive years. Calves were weaned individually at 270 days of age and allocated to post-weaning stocking rate groups balanced for dam's stocking rate, sex and body weight at 90 d (Richardson, 1983). The post-weaning stocking rates were 0.22 and 0.43 animals/ha. The experiment was carried out on a large heterogeneous area of land that included rocky hillsides, flat land and fertile riverine areas, while the grass growth model was developed from experiments on a uniform area of flat land. Every effort had been made to allocate equitable areas of each type of land to each stocking rate treatment, but the proportion of high quality grazing allocated to the heavy stocking rate treatment may have been greater than that for the light stocking rate. Consequently, simulations of the HSR treatment were performed using two values for stocking rate, 0.278 and 0.200 cows/ha. Model predictions of cow and calf live weights were closer to the observed values if the stocking rate of 0.200 cows/ha was used. The performance of cows and suckling calves was simulated for the years, 1978/9 and 1979/80, using the recorded rainfall data. During the 1979/80 the model predicted that the calves of LSR calves were heavier at birth than those of HSR cows. The differences in live-weight increased progressively until weaning at 270 days of age (Table 1). The simulated effect of stocking rate on live weights at birth and weaning were closely similar to those observed in the field experiment (Table 1) (Richardson, 1983). The model predicts that LSR calves consumed more milk and herbage, 747 and 551 kg, respectively, than HSR calves, 602 and 428 kg. Among animals stocked at 0.22 animals/ha after weaning the model predicted that animals whose dams had been stocked lightly would increase their weight advantage slightly until slaughter at two years seven months (Table 1), while the experimental results showed that that the calves of LSR calves maintain their weight advantage during the same period. The small progeny of HSR cows consumed less forage between weaning and slaughter, 2564 kg, than the progeny of LSR cows, 2961 kg.
The model was then used to study whether animals aged 21 months that had attained a weight of 278 kg and subjected to undernutrition during their second dry season that caused a loss of 40 kg would exhibit compensatory growth. The model predicted that when the restricted and unrestricted animals grazed together as one herd, the difference in live weight would be reduced to 27 kg within 140 days. The reason for this partial compensation was that the maturity of capacity of the digestive system was similar for both the restricted and unrestricted animals until they were 736 days old. Consequently due to their lighter weight and lower maintenance expenditure they were able to use a larger portion of their dietary energy for gain than their larger unrestricted contemporaries. Partial compensation of this magnitude by unsupplemented animals when compared with those given a protein supplement was reported by Murray et al. (1936).
Conclusion
The model predicts that potential forage intake in growing cattle is a function of maturity of live weight but does not decrease if an animal loses weight. This explains why the capacity for compensatory growth varies with age when under-nutrition occurs. The model predicts that on semi-arid range the calves of heavily stocked cows do not compensate for the effects of pre-weaning under-nutrition even at light stocking rates.
References
Ainslie, K.S., 1958. A study of the influence of seasonal fluctuations in the nutritive value of natural grazing in the medium rainfall areas of Southern Rhodesia on the growth and development to the beef steer and carcass. Rhod. Agric. J. 55, 286. [ Links ]
Butterfield, R.M., 1988. New Concepts of Sheep Growth. Department of Veterinary Anatomy, University of Sydney, Australia. [ Links ]
Butterworth, M.H. (1985). Beef Cattle Nutrition and Tropical Pastures. Longman, London, UK. [ Links ]
Finlayson, J.D., Cacho, O.J. & Bywater, A.C., 1995. A simulation model of grazing sheep: 1. Animal growth and intake. Agric. Syst. 48, 1-25. [ Links ]
Greenwood, P.L. & Café, L.M., 2007. Prenatal and pre-weaning growth and nutrition of cattle: long-term consequences for beef production. Animal 1, 1283-1296. [ Links ]
Kahn, H.E. & Spedding, C.R.W., 1984. A dynamic model for the simulation of cattle herd production systems: 2-an investigation of various factors influencing the voluntary intake of dry matter and the use of the model in their validation. Agric. Syst. 13, 63-82 [ Links ]
Mellor, D.J. & Murray, L., 1985. Effects of maternal nutrition on udder development during late pregnancy and on colostrum production in Scottish Blackface ewes with twin lambs. Res. Vet. Sci. 39, 230-234. [ Links ]
Morgan, J.H.L., 1972. Effect of plane of nutrition in early life on subsequent live-weight gain, carcass characteristics and eating quality of meat in cattle. J. Agric. Sci., Camb. 78, 417-423. [ Links ]
Murray, C.A., Romyn, A.E., Haylett, D.G. & Ericksen, F., 1936. The supplementary feeding of mineral and protein to growing cattle in Southern Rhodesia and its relation to the production of beef steers. Rhod. Agric. J. 33, 422-440. [ Links ]
O'Donovan, P.B., 1984. Compensatory gain in cattle and sheep. Nutr. Abstr. Rev. Series B 54, 389-410. [ Links ]
Poppi, D.P., Gill, M. & France, J., 1994. Integration of theories of intake regulation in growing ruminants. J. Theoret. Biol. 167, 129-145. [ Links ]
Richardson, F.D., 1983. Short and long-term influences of undernutrition on range cattle production. Zimbabwe Agric. J. 80, 175-182. [ Links ]
Richardson, F.D., Hahn, B.D. & Schoeman, S.J., 2000. Modelling nutrient utilization by livestock grazing semi-arid rangeland. In: Modelling Nutrient Utilization in Farm Animals. Eds McNamara, J.P., France, J. & Beever, D., CABI, Wallingford, Oxon. pp. 263-280. [ Links ]
Richardson, F.D., Oliver, J. & Clark, G.P.Y., 1977. Analysis of some factors which affect the productivity of beef cows and of their calves in a marginal rainfall area of Rhodesia. 2. The Yield and composition of the milk of suckling cows. Anim. Prod. 25, 359-372. [ Links ]
Richardson, F.D., Oliver, J. & Clarke, G.P.Y., 1979. Analysis of some factors which affect the productivity of beef cows and of their calves in a marginal rainfall area of Rhodesia. 3. Factors affecting calf birth weight, growth to 240 days and the use of concentrates by cows and their calves. Anim. Prod. 28, 199-211. [ Links ]
Thornley, J.H.M., 1998. Grassland Dynamics: An ecosystem simulation model. CAB International, Wallingford, Oxon. [ Links ]
# Corresponding author. E-Mail: fd.rich@mweb.co.za


{kind=link}
{kind=link}