










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkSouth African Journal of Animal Science
versão On-line ISSN 2221-4062
versão impressa ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 no.4 Pretoria Jan. 2009
Dietary vitamin E (α-tocopheryl acetate) and organic selenium supplementation: performance and antioxidant status of broilers fed n-3 PUFA-enriched feeds
H. Basmacıoğlu MalayoğluI,#; S. ÖzkanI; S. KoçtürkII; G. OktayII; M. ErgülI
IFaculty of Agriculture, Department of Animal Science, Ege University, İzmir, Turkey
IIFaculty of Medicine, Department of Biochemistry, Dokuz Eylül University, İzmir, Turkey
ABSTRACT
The experiment was conducted to study the effects of organic selenium (Se-enriched yeast) and vitamin E (α-tocopheryl acetate) supplementation, alone or together, on the performance and antioxidant status of broilers fed diets enriched in n-3 PUFAs using fish oil. Day-old Hubbard-JV strain broiler chicks allocated to one of the following treatments: (1) a commercial basal diet containing 0.15 mg inorganic Se /kg as sodium selenite and 50 mg α-tocopheryl acetate (α-TA)/kg feed acted as the control; (2) VE200: Control diet supplemented with 200 mg α-TA/kg feed; (3) OrgSe0.15: Control with inorganic Se replaced with 0.15 mg organic Se/kg; (4) OrgSe0.30: Control with inorganic Se replaced with 0.30 mg organic Se/kg feed; (5) VE200+OrgSe0.15, (6) VE200+OrgSe0.30. Body weight (BW) and body weight gain (BWG) to 42 d were significantly improved with vitamin E or organic Se at 0.30 mg/kg (but not at 0.15 mg/kg) when supplemented individually, but not in combination, except that VE200+OrgSe0.15 improved 28 and 35-d body weights significantly compared with the control. Feed intake (FI), feed conversion ratio (FCR), mortality, carcass characteristics and relative organ weights, except for the spleen, were unaffected by any of the supplementation treatments. The significant positive effect of VE200 on BWG was observed after 21 d. Relative spleen weight was significantly higher in broilers fed VE200 compared to the other treatments. The antioxidative effects of organic Se and vitamin E, alone or together, were not evident in diets enriched in n-3 PUFAs using 1.5% fish oil.
Keywords: Fish oil, n-3 PUFAs, broiler, vitamin E, organic selenium, antioxidant
Introduction
The production of reactive free radicals (ROS) as by-products of metabolism that have the potential to damage or destroy cellular structures is in a dynamic equilibrium under normal conditions in living organisms. This dynamic equilibrium is provided by a balance between antioxidants and pro-oxidants (Koinarski et al., 2005). However, stress factors such as nutritional (low digestible feed, feeds rich in polyunsaturated fatty acids (PUFAs), mycotoxins and oxidized oil, vitamin E and selenium (Se) deficiency, vitamin A excess, the presence of heavy metals and other toxicants), environmental (high or low ambient temperatures, transportation) and pathogenesis of numerous diseases including parasitic infections, have a negative impact on this antioxidant/pro-oxidant balance (Surai, 2002b; Koinarski et al., 2005). The imbalance between an antioxidant and pro-oxidant system is named oxidative stress. In commercial poultry production, oxidative stress has been associated with the deterioration of many physiological functions including health, growth, reproduction and immunity. Intracellular defence mechanisms against ROS-induced damage may be classified as non-enzymatic (ascorbate, α-tocopheryl, carotenoids and glutathione) or enzymatic antioxidants, such as superoxide dismutase (SOD), catalase and glutathione peroxidases (GSHPx) (Surai, 2002a; b).
In recent years health conscious consumers have stepped up their demands for specific foods or physiologically active food components; these being the so-called functional or designer foods. Meat or eggs enriched with N-3 PUFA's [eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)] have the potential to be regarded as credible functional foods, because of considerable evidence of their health benefits, such as reducing coronary artery diseases, hypertension, diabetes, as well as certain inflammatory diseases such as arthritis and dermatitis (Simopoulos, 2000). Numerous studies have focused on the enrichment of poultry meat and eggs with n-3 PUFAs (Sirri et al., 2002; Basmacıoğlu et al., 2003). The n-3 PUFA content of poultry products can be increased effectively through dietary manipulation, either directly using fish oil (Basmacıoğlu et al., 2003) and marine algae (Sirri et al., 2002) or indirectly by increasing levels of precursor n-3 PUFAs such as alpha-linolenic acid, in the form of linseeds, flaxseed or similar oils (López-Ferrer et al., 1999). Fish oil is a common feed ingredient used to increase n-3 PUFA content of meat and eggs. However, the use of fish oil at concentrations greater than 1 to 2% in poultry diets may introduce several organoleptic problems in the final product that compromise meat or egg consumption (Hargis & Van Elswyk, 1993).
However, n-3 PUFA-enriched diets increase both the susceptibility of cellular membranes to the induction of oxidative stress in animal organisms (Miret et al., 2003) and the susceptibility of products to lipid oxidation, which can impair meat and egg quality such as off-tastes and off-odours, thereby reducing consumer acceptability (Sparks, 2006). Husveth et al. (2000) reported that there was a significant positive correlation between liver thiobarbituric acid-reactive substances (TBARS) and n-3 PUFA content, and suggested that feeding oils rich in n-3 PUFAs increase tissue concentrations of these fatty acids. The consequence is increased tissue lipid peroxidation and reduced antioxidative status of broiler chicks. In this respect, dietary antioxidants such as vitamin E, carotenoids, herbal extracts and selenium (Se) are beneficial in preventing these detrimental effects (Fellenberg & Speisky, 2006). Vitamin E and Se are key components of the antioxidant system, reducing lipid peroxidation (Combs, 1981).
Vitamin E is one of the antioxidants widely used in poultry diets and has been proposed as a major antioxidant in plasma membranes of all cells and sub-cellular organs, functioning as a chain-breaker and free radical scavenger. Poultry cannot synthesize vitamin E and its concentration is reduced under stress conditions. As dietary PUFA levels increase, the alpha-tocopherol content of organs and products decreases. The undesirable oxidative effects of n-3 PUFAs on rat erythrocytes were prevented when sufficient vitamin E was supplied (Ando et al., 1998).
Selenium plays an important role in the antioxidant defence system due to its requirement by the Se-dependent GSHPx, which is involved in cellular antioxidant protection. It has been suggested that there is a synergistic relationship between Se and vitamin E, because GSHPx continues the work of vitamin E by detoxifying hydroperoxides. Recent understanding of antioxidant system functions and new discoveries regarding the GSHPx enzyme family are the basis for further development in the Se nutrition of poultry (Surai, 2002b).
The recommended Se concentration in broiler diets is 0.15 mg/kg throughout the growth period (NRC, 1994). Dietary antioxidant supplementation with levels exceeding nutritional requirement improved the bird's antioxidant defence system and decreased oxidative stress. Inorganic (selenite, selenate, selenide) and organic forms of Se (Se enriched yeast, Se enriched algae) may be used as supplements. Traditionally, the Se supplement in animal diets' has been the inorganic form, as sodium selenite. Recently, however, organic sources of Se have been explored as an alternative to inorganic supplementation. The amount of Se available for assimilation by the tissue is dependent on the source, and it has been shown that organic Se is deposited into the animal tissue more efficiently than inorganic Se (Choct et al., 2004).
The effects of supplementing n-3 PUFA-enriched feeds with vitamin E or Se on antioxidant status have been studied in the mouse (İbrahim et al., 1997), rat (Sen et al., 1997) and hamster (Poirier et al., 2002). However, there is a general lack of information about the effects of vitamin E or organic Se, alone or combined, on broiler performance and antioxidant status under nutritional oxidative stress conditions related to feeds enriched in n-3 PUFAs.
The objective of this study was to determine the effect of supplemental dietary antioxidants on oxidative stress conditions resulting from enriching feeds with n-3 PUFAs. Thus, the study aimed at evaluating the effects of organic Se at two dietary levels (0.15 and 0.30 mg/kg) and vitamin E as α-tocopheryl acetate (α-TA) at 200 mg/kg, supplemented alone or together, on the performance and antioxidant status of broilers fed diets enriched with n-3 PUFAs supplied as fish oil at a level of 1.5%.
Materials and Methods
A total of 540 unsexed Hubbard-JV broiler chicks were obtained from a commercial hatchery. One-day-old chicks were weighed, wing-banded and randomly distributed to the six treatment groups with three replicates. Each of the 18 floor pens was equipped with wood shaving as litter. Housing density was 12 birds/m2 and a fluorescent lighting schedule of 23 h light and 1 h dark was used during the experiment with an average light intensity of 20 lux. Birds were given one of six feeds in mash form. The ingredients and calculated nutritional composition of commercial basal feeds are given in Table 1. The feeds were formulated to meet or exceed minimum National Research Council recommendations (NRC, 1994) for all nutrients. The starter diet (0 to 21 d) contained 220 g crude protein and 12.6 MJ/ metabolisable energy (ME)/kg and the finisher (22 to 42 d), 198 g crude protein and 13.0 MJ ME/kg.
These basal feeds, used as the control, were supplemented with a vitamin premix containing 50 mg α-TA/kg feed and a mineral premix containing 0.15 mg inorganic Se (sodium selenite)/kg feed. Organic Se, as Se-enriched yeast (Sel-Plex®), was provided by Alltech Inc. Company, Turkey. The organic Se (Se-yeast) contained 1 000 mg Se/kg and it was added at either 0.15 or 0.3 g/kg feed mixture. Inorganic Se (sodium selenite) was replaced with organic Se in the mineral premix.
Vitamin E at a level of 200 mg/kg feed was added to the vitamin premix (contained 50 mg α-TA/kg) and thus, diets with vitamin E contained a total of 250 mg α-TA/kg. All premixes used in the study were prepared by a commercial company (Eryaş Ltd., İzmir-Turkey).
Thus, the experimental diets included (1) a control being the commercial basal diet containing 0.15 mg/kg inorganic Se as sodium selenite and 50 mg/kg α-tocopheryl acetate (α-TA); (2) VE200: The control diet supplemented with 200 mg α-TA/kg feed; (3) OrgSe0.15: The control with inorganic Se replaced with 0.15 mg organic Se/kg; (4) OrgSe0.30: The control with inorganic Se replaced with 0.30 mg organic Se/kg feed; (5) VE200+OrgSe0.15; (6) VE200+OrgSe0.30.
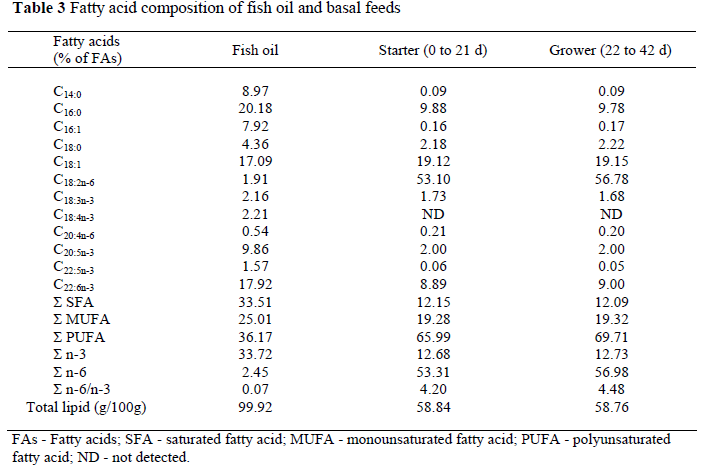
Selenium and vitamin E (α-tocopherol) content of the experimental feeds are presented in Table 2. All experimental feeds contained fish oil (15 g/kg) to increase the n-3 PUFAs content. The fatty acid composition of fish oil, and the basal feeds used in the study are given in Table 3.
Body weight (BW) of broilers in each pen was measured individually on a weekly basis. Chicks were weighed to ± 0.1 g at hatch and to 1 g thereafter. At the same time feed intake (FI) was recorded to the nearest g and feed conversion ratio (FCR) was calculated on a pen basis (kg feed/kg gain). Mortality was recorded daily.
At 42 days, six birds (three males and three females per treatment) were randomly selected from each treatment group. Blood samples obtained from the brachial veins were collected into tubes containing anticoagulant for assays of SOD and GSHPx. After obtaining blood samples, the birds were humanely killed by cervical dislocation for organ sampling. Organs (intestinal, gizzard, proventriculus, liver, heart and spleen) were dissected and weighed to the nearest 0.1 g. Organ weights were presented as relative to body weight (g/100 g BW). Liver samples were washed with phosphate buffer, pH 7.4, and immediately placed in plastic bags in dry ice and stored at -80 ºC for assay of SOD.
At the end of the trial six birds (three males and three females per treatment) were randomly selected from each treatment group and slaughtered to determine carcass characteristics. All birds were fasted for 12 h prior to slaughter, then weighed. Thereafter, birds were subjected to standard commercial evisceration procedures. Carcass yield (without feathers, head, feet and digestive organs) was expressed as a percentage of the live body weight just before slaughter, and carcass components (breast and thigh) were expressed as relative to body weight (g/100 g BW).
Standard proximate analysis techniques were used to determine nutrient concentrations, while starch, sugar, total calcium and phosphorus were also analyzed using the VDLUFA methods (Naumann & Bassler, 1993). Lipids in the samples were extracted according to the method of Bligh & Dyer (1959). Fatty acid methyl esters of feed were determined on extracted lipids according to the method modified by Tokuşoğlu & Ünal (2003). Lipids extracted from feed samples were refluxed using 0.05N NaOH with methanol and then derivatised with boron trifluoride (BF3) - methanol complex. This solution was then fractionated with saturated NaCl; the methyl ester (ME) phase was separated, and anhydrous sodium sulphate (Na2SO4) added to the final solution. Final extract was degassed for up to 2 min to eliminate oxygen. Fatty acid profiles were determined by gas chromatography [Equipment: Perkin Elmer (auto system), column: SGE (BP70X), 60 m-capillary; 0.25 µm film thickness; 0.25-mm diameter WCOT fused-silica), detector: flame ionization detector (FID), column temperature: ramp 1:165 ºC (increase with 1 ºC/min) (2 min isotherm), ramp 2:225 ºC (increase with 2 ºC/min) (30 min isotherm), detector temperature: 250 ºC, injector temperature: 250 ºC, elution time: 35 min, split ratio: 50:1, carrier gas: Helium (He), flow rate:1.5 mL/min].
Vitamin E concentrations in the experimental diets were determined according to the modified method of Qureshi et al. (2000). High performance liquid chromatographic (HPLC) conditions were performed for the α-tocopherol analysis. The HPLC system (Shimadzu) was equipped with an autosampler (SIL-10AD vp), system controller (SCL-10 Avp), pump (LC-10 ADvp), degasser (DGU-14A) and column oven (CTO-10 Avp), and the column temperature was 40 ºC. A fluorescence detector was used with wavelengths set at 295 nm for extinction and 330 nm for emission. Tocopherols were separated on a normal phase column (Luna, 150 x 4.6 mm i.d., 5 µm particle size) with mobile phase flow rate 1.2 mL /min. The mobile phase was a mixture of heptanes: THF (95:5) (v/v). The diets were analyzed for Se by the methods of AOAC (2005) using an atomic absorption spectrophotometer. Before analysis, all feed samples were wet digested using a mixed solution of concentrated perchloric and nitric acids. Lipid peroxidation in liver was estimated based on TBARS. Samples were evaluated for malondialdehyde (MDA) production using a spectrophotometric assay for TBA (Draper & Hadley, 1990). The extinction coefficient of 153/mM/cm at 532 nm for the chromophore was used to calculate the MDA-like TBA. The total protein content was determined by the method of Bradford (1976) using bovine serum albumin (BSA) as standard.
The activity of SOD in whole blood and tissues was determined by spectrophotometry with the use of a Ransod kit (Randox Laboratories Ltd., UK) and the method described by McCord & Fridovich (1969). The activity of blood GSHPx was determined by spectrometry with the use of Ransol kit (Randox Laboratories Ltd., UK) and the method described by Paglia & Valentine (1967). Antioxidant enzyme activities in blood were presented as unit per mL. SOD activity in liver was presented as unit per wet tissue.
General Linear Model using the SPSS (11.0)® statistic package (SPSS, 2001) was applied to data with a model including treatment and sex effects and interactions between them. When the effect was significant, the differences between treatment group means were separated by Duncan's multiple range tests. Results are presented as least square means and standard error of means. Statements of statistical significance are based on P <0.05.
Results and Discussion
Dietary analysis showed that the Se content of the control diet (supplemented with 0.15 mg inorganic Se/kg), OrgSe0.15 (0.15 mg organic Se/kg) and OrgSe0.30 was 0.26, 0.27 and 0.51 mg Se/kg of feed, respectively in starter diets, and 0.24, 0.22 and 0.45 mg Se/kg of feed, respectively in grower diets (Table 2).
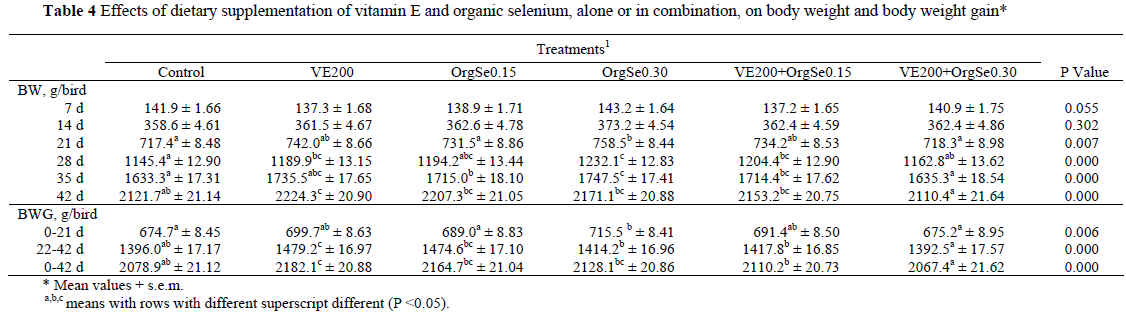
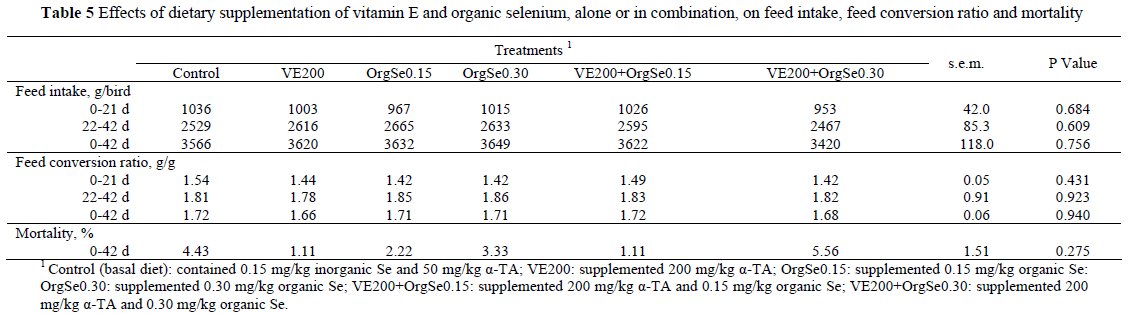
The average BW of chicks at hatching was 42.7 ± 0.13 g and did not differ statistically among treatment groups (data not shown). In the present study, the antioxidant supplementation to diets enriched with n-3 PUFAs improved (P <0.05) BW and BWG of the broilers at 21, 28, 35 and 42 d. However, FI, FCR and mortality were not affected by antioxidant supplementation (Tables 4 and 5).
The highest level of organic Se (0.30 mg/kg) resulted in increases (P <0.05) in the BW of the birds from 21 to 35 d, while the lower level (0.15 mg/kg) produced heavier birds only at 35 d. A significant improvement (P <0.05) in BWG was evident only during the early period of growth (0 - 21 d), and only on the higher concentration of Se. Organic Se supplementation did not affect FCR or FI.
Growth performance was the same on both Se sources (inorganic as selenite and organic as Se-enriched yeast) at the level of 0.15 mg/kg, suggesting that organic Se used at that concentration is no more bioavailable than inorganic Se used at the same level. However, when organic Se at 0.15 mg/kg was combined with vitamin E (VE200+OrgSe0.15), a significantly higher (P <0.05) BW at 28 and 35 d was noted compared with the control, whereas no significant improvement was observed relative to the feeds supplemented with organic Se or vitamin E alone. No significant improvement resulted from the supplementation of vitamin E and Se at 0.30 mg/kg compared with the other treatments.
Although Se levels in feeds used in the present study exceeded the NRC (1994) recommendation of 0.15 mg for broilers, the lower levels used (measured to be 0.26 - 0.27 mg/kg feed) were not sufficient to allow broiler chicks to attain the BWG of those fed OrgSe at 0.30 mg/kg feed, and this was irrespective of the source of supplementation.
Edens (2001) and Surai (2006) noted that the improved growth rate of broilers fed an organic Se supplemented diet could be related to the increased concentrations of the active form of thyroid hormone in the serum of chickens supplemented with organic Se as well as to the immunomodulating properties of Se.
Many of the studies related to organic Se were conducted under normal (Choct et al., 2004; Payne & Southern, 2005) or oxidative stress (Mahmoud & Edens, 2005; Özkan et al., 2007) conditions. Mahmoud & Edens (2005) evaluated the effect of organic Se on performance and different physiological parameters of broilers either in pathogenic Escherichia coli-challenged or under heat-stressed conditions. They found that feeds supplemented with 0.20 mg organic Se/kg feed, improved BW, FCR and mortality at 42 days of age. Özkan et al. (2007) found that birds fed diets supplemented with organic Se at 0.30 mg/kg feed and raised in cold conditions tended to be heavier and have higher daily BWG than birds fed diets supplemented with inorganic Se. However, they found that FCR was not affected by treatment during the trial. Surai (2006) suggested that the growth response of birds to Se supplementation would also depend on the birds' stress level, with stressed birds being more responsive. Some reports (Choct et al., 2004; Payne & Southern, 2005) indicated that dietary supplementation of Se (organic or inorganic) did not affect BW or BWG of broilers reared at optimum (no oxidative stress) conditions. In contrast, organic Se supplementation increased BW of broilers at 42 days of age (Edens, 2001) and improved feed conversion (Naylor et al., 2000) in broilers reared under normal commercial conditions. In the present study, birds fed VE200 (supplemented with vitamin E only) had higher (P <0.05) BWG than birds fed the control diet after 21 d (22 - 42 and 0 - 42 d, Table 4). However, FI and FCR were unaffected by this supplementation (Table 5).
There is considerable inconsistency in the literature pertaining to this subject: In our previous study (Basmacıoğlu et al., 2004), vitamin E supplementation to diets enriched with n-3 PUFAs significantly increased BWG from 22 - 42 d, but did not affect FI, FCR or mortality. In contrast, it was noted that less food was consumed by hamsters fed fish oil-containing diets supplemented with vitamin E than unsupplemented diets, while BWG was not affected by vitamin E supplementation (Poirier et al., 2002). In a study conducted with layer hens, it was found that vitamin E supplementation at different levels to diets enriched with PUFAs did not affect performance (Meluzzi et al., 2000; Sijben et al., 2002) and in contrast to these studies, Scheideler & Froning (1996) noted that vitamin E supplementation (50 IU/kg) to a diet enriched with n-3 PUFAs significantly improved egg production in layer hens. While some reports (Coetzee & Hoffman, 2001; Guo et al., 2001; Özkan et al., 2007) describe a lack of response in performance to supplementation with vitamin E under normal conditions, others have shown improvements in performance. For example, Kennedy et al. (1991) reported that FCR was improved by 0.8% and the average weight per bird was increased by 1.4% compared with those receiving normal diets. Chung & Boren (1999), who evaluated the benefits of vitamin E supplementation in broiler diets, observed that 240 mg vitamin E/kg improved FCR by 2.3% compared to the control group (33 mg VE/kg), and Swain et al. (2000) showed that vitamin E supplementation at levels of 150 and 300 IU/kg resulted in significant increases in BWG and FCR compared with the control diet (no VE supplementation) at 42 d under normal conditions. Established NRC requirements (NRC, 1994) may be optimal for growth but not necessarily for best health, disease resistance and oxidative stress conditions. It is of particular interest to further examine the role that vitamin E plays on the immune system when poultry are under oxidative stress. Mcllroy et al. (1993) concluded that dietary supplementation with vitamin E at levels exceeding nutritional requirements improved the birds' antioxidant defence system and decreased oxidative stress.
Even the positive effects of vitamin E or organic Se on weight gain disappeared when used in combination in the study. These results agree with those of Özkan et al. (2007), who reported that vitamin E supplementation at 200 mg/kg with organic or inorganic Se at 0.30 mg/kg had no significant positive effect on BWG or FCR. These reports are in contrast with those of Stanley et al. (1998) who noted that 0.1 mg/kg of organic Se with 500 mg/kg vitamin E supplementation decreased mortality and incidence of pulmonary hypertension syndrome in birds reared under cold stress and fed aflatoxin-contaminated feeds. Roch et al. (2000) also noted that the combination of 0.3 mg organic Se/kg with 250 mg vitamin E/kg improved FCR cold-stressed broilers.
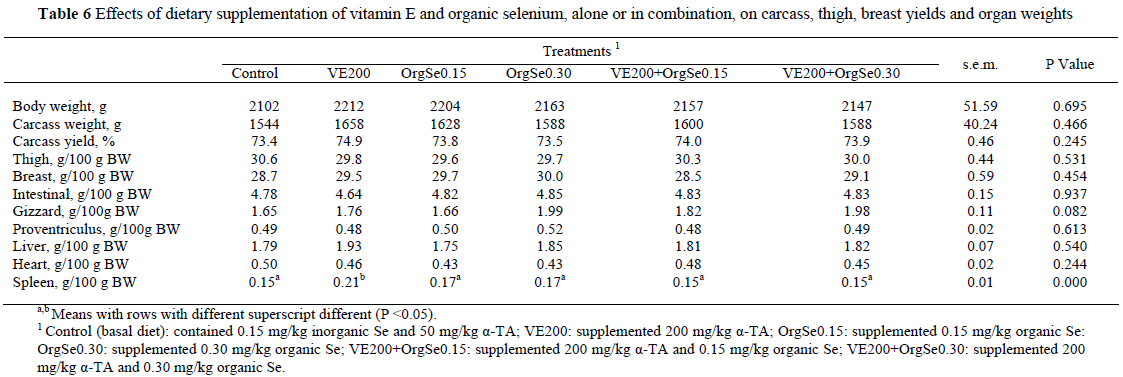
In the present study, vitamin E or organic Se supplementation to diets enriched with n-3 PUFAs did not affect the carcass characteristics or some organ weights (except for spleen weights) in broilers (Table 6). Relative spleen weight of birds fed VE200 was significantly higher than that of birds fed the other treatment diets (0.15 - 0.17 in the other treatments group vs. 0.21 g/100 BW in the VE200 treatment group).
Few studies have investigated the effects of dietary antioxidant supplementation on cut-up yields of broilers under stress conditions. In the studies related to Se supplementation, it is noted that form of dietary supplementation also appears to affect cut-up yields of broilers. For example, Naylor et al. (2000) noted that organic Se supplementation improved eviscerated weight and breast yield of broilers. In another study (Edens, 2001), percentage of carcass weight increased when organic Se was added to the diet. However, pectoralis major yield decreased as a result of organic Se supplementation in the same study. Surai (2002a) noted that the effect of Se on meat yield could be due to changes in thyroid hormone metabolism or a result of changes in broiler feathering. However, in this study, the supplementation of organic Se to n-3 PUFA-enriched diets did not affect meat yield (Table 6).
In the present study, relative spleen weight significantly increased in broilers fed VE200 which may indicate an improvement in the immune system. A similar result was reported by Konjufca et al. (2004). They found that vitamin E supplementation as Dl-α-tocopherol did not increase relative organ weight, except for the spleen. They concluded that the increase in spleen weight with supplemental vitamin E was likely to represent an increase in the number of lymphocytes.
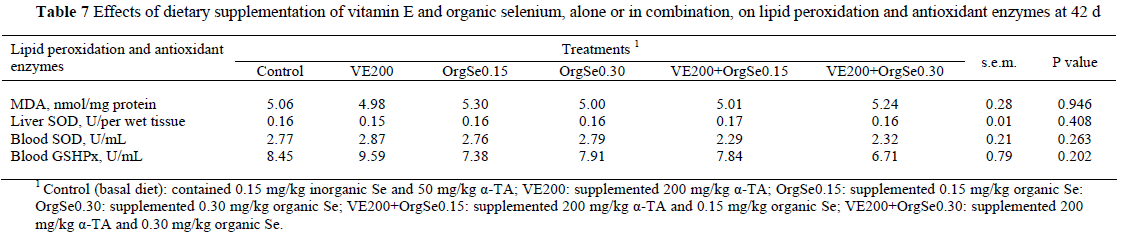
The vitamin E and organic Se supplementation to diets enriched with n-3 PUFAs did not significantly prevent lipid peroxidation in liver or increase antioxidant enzyme activities (SOD and GSHPx) in liver and blood (Table 7).
In general, studies have been conducted to determine the effects of supplemental antioxidants to PUFA-enriched diets with fish oil on oxidative stress in rats or mice, but few studies conducted with broilers have been reported in the literature. In addition, the results from studies are inconsistent. İbrahim et al. (1997) noted that these inconsistencies may be due to differences in the species of experimental animals used (rats vs. mice), or to dietary composition or duration of the experiment. Polavarapu et al. (1998) noted that species difference and supplemental level of fish oil may be important in this regard. Moreover, Poirier et al. (2002) reported that tissue and species-specific responses of antioxidant enzymatic defences and lipid peroxidation can occur with antioxidant supplementation.
Oxidative stress may not have been induced in this study by the addition of fish oil at a level of 1.5%. Therefore, the positive effect of antioxidants might not have been seen in these broilers. We conclude that the effect of antioxidants would depend on the birds' stress level. In a study conducted with broilers, Eid et al. (2006) reported that the decline in GSHPx activity at high levels (>2.5%) of fish oil could be explained by the presence of high levels of free radicals which induced oxidative stress in the body due to high levels of n-3 PUFAs. İbrahim et al. (1997) who measured the effect of dietary lipid (8% fish and lard oils), vitamin E and iron on hepatic antioxidant status in mice, reported that vitamin E supplementation to diets with fish oil did not significantly alter hepatic SOD and GSHPx. However, Miret et al. (2003) reported that SOD and GSHPx activities were lower in erythrocytes of rats fed fish oil at a level of 5%, and that vitamin E supplementation to diets containing 5% fish oil increased these antioxidant enzyme activities in erythrocyte membranes, but that each enzyme responded in a specific manner. No studies have reported the effect of organic Se supplementation to n-3 PUFA-enriched diets as oxidative stress factor in broilers. Thus, the results obtained from this study could be compared with results of studies conducted under environmental stress. Mahmoud & Edens (2005), who replaced inorganic Se with 0.2 mg/kg of organic Se in the diet of growing chicks, reported that organic selenium supplementation was associated with significantly increased plasma GSHPx activity in chickens reared under environmental oxidative stress. However, in the present study no additional benefit over that attainable from feeding either vitamin E or organic Se alone was obtained from feeding the organic Se supplement in combination with the vitamin E supplement on the antioxidant enzymes. Özkan et al. (2007) had noted previously that 200 mg/kg vitamin E supplementation with organic Se at level of 0.30 mg/kg did not cause any positive effect on liver GSHPx activity in cold-stressed broilers. However, Roch et al. (2000) found that diets supplemented with 0.3 mg/kg of organic Se plus 250 mg/kg vitamin E increased plasma GSHPx levels and decreased ascites mortality in cold-stressed broilers.
Conclusions
Supplementation of vitamin E at 200 g/kg or organic Se at level of 0.30 mg/kg to broiler diets enriched with n-3 PUFAs significantly increased BWG, but did not affect FI, FCR, mortality, carcass characteristics or organ weights, with an exception of spleen weight. The positive effect of organic Se on growth performance may be attributed to its ability to increase the active form of thyroid hormone and to its immunomodulating properties. Vitamin E supplementation significantly increased relative spleen weight, which indicated a benefit for the immune system. Although organic Se and vitamin E improved growth in broilers in this trial, their antioxidative effects were not evident in diets enriched in n-3 PUFAs using 1.5% fish oil. We conclude that 1.5% fish oil may not cause significant oxidative stress in living organism.
In further studies the effects of organic Se and vitamin E on metabolic hormones and immunity should be investigated, and higher levels of fish oil or PUFAs should be considered when measuring the effects of antioxidant supplementation to diets enriched in PUFAs using fish oil. In addition, the effect on the oxidative stability of meat from broilers fed n-3 PUFA-enriched feed, supplemented with vitamin E or organic Se, alone or in combination, could be investigated.
Acknowledgements
This study was funded by the Scientific Research Projects Committee at Ege University (Project number 2003-ZRF-021). We wish to thank Alltech Turkey for supplying Sel-Plex and Eryaş firm, Turkey for preparing mineral-vitamin premixes used in the study.
References
Ando, K., Nagata, K., Beppu, M., Kikugawa, K., Kawabata, T., Hasegawa, K. & Suzuki, M., 1998. Effect of n-3 fatty acid supplementation on lipid peroxidation and protein aggregation in rat erythrocyte membranes. Lipids 33, 505-512. [ Links ]
AOAC, 2005. Official Methods of Analysis, Association of Official Analytical Chemists. Arlington, Virginia, USA. [ Links ]
Basmacıoğlu, H., Çabuk, M., Ünal, K., Özkan, K., Akkan, S. & Yalçın, H., 2003. Effects of dietary fish oil and flaxseed on cholesterol and fatty acid composition of egg yolk and blood parameters of laying hens. S. Afr. J. Anim. Sci. 33, 265-273. [ Links ]
Basmacıoğlu, H., Tokuşoğlu, Ö. & Ergül, M., 2004. The effect of oregano and rosemary essential oils or alpha-tocopheryl acetate on performance and lipid oxidation of meat enriched with n-3 PUFAs in broilers. S. Afr. J. Anim. Sci. 34, 197-210. [ Links ]
Bligh, G.E. & Dyer, J.W., 1959. A rapid method of total lipid extraction and purification. Biochem. Cell Biol. 37, 911-917. [ Links ]
Bradford, M.M., 1976. A rapid and sensitive method for quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248-252. [ Links ]
Choct, M., Naylor, A.J. & Reinke, N., 2004. Selenium supplementation affects broiler growth performance, meat yield and feather coverage. Br. Poult. Sci. 45, 677-683. [ Links ]
Chung, T.K. & Boren, B., 1999. Vitamin E use in commercial flocks examined. Feedstuffs 6, 11-14. [ Links ]
Coetzee, G.J.M. & Hoffman, L.C., 2001. Effects of dietary vitamin E on the performance of broilers and quality of broiler meat during refrigerated and frozen storage. S. Afr. J. Anim. Sci. 31, 158-172. [ Links ]
Combs Jr., G.F., 1981. Influences of dietary vitamin E and selenium on the oxidant defence system of the chick. Poult. Sci. 60, 2098-2105. [ Links ]
Draper, H.H. & Hadley, M., 1990. Malondialdehyde determination as index of lipid peroxidation. Meth. Enzymol. 186, 421-431. [ Links ]
Edens, F.W., 2001. Involvement of Sel-Plex in physiological stability and performance of broiler chickens. Biotechnology in Feed Industry. Proc. 17th, Alltech Ann. Symp. 349-376. (Nottingham, Nottingham University Press). [ Links ]
Eid, Y., Ebeid, T., Saleh, A. & El-Hamid, H. ABD., 2006. Effects of n-3 enriched egg production on lipid peroxidation and antioxidative status in laying hens. XII European Poultry Conference, Verona-Italy, 10-14 September. [ Links ]
Fellenberg, M.A. & Speisky, H., 2006. Antioxidants: their effects on broiler oxidative stress and its meat oxidative stability. Wrld's Poult. Sci. J. 62, 53-70. [ Links ]
Guo, Y., Tang, Q., Yuan, J. & Jiang, Z., 2001. Effects of supplementation with vitamin E on the performance and the tissue peroxidation of broiler chicks and the stability of thigh meat against oxidative deterioration. Anim. Feed Sci. Technol. 89, 165-173. [ Links ]
Hargis, P.S. & Van Elswyk, M.E., 1993. Manipulating the fatty acid composition of poultry meat and eggs for the healthy conscious consumer. Wrld's Poult. Sci. 49, 251-264. [ Links ]
Husveth, F., Manila, H.A., Gaal, T., Vajdovich, P., Balogh, N., Wagner, L., Loth, I. & Nemeth, K., 2000. Effects of saturated and unsaturated fats with vitamin E supplementation on the antioxidant status of broiler chicken tissues. Acta Vet. Hung. 48, 69-79. [ Links ]
İbrahim, W., Lee, U.S., Yeh, C.C., Szabo, J., Bruckner, G. & Chow, C.K., 1997. Oxidative stress and antioxidant status in mouse liver: effects of dietary lipid, vitamin E and iron. J. Nutr. 127, 1401-1406. [ Links ]
Kennedy, D.G., Goodall, E.A., Mcllroy, S.G., Bruce, D.W. & Rice, D.A., 1991. The effects of increased vitamin E supplementation of profitable commercial broiler production. Proc. Nutr. Soc . 50, 197A. [ Links ]
Koinarski, V., Georgieva, N., Gadjeva, V. & Petkov, P., 2005. Antioxidant status of broiler chickens, infected with Eimeria acervulina. Revue Méd. Vét. 156, 498-502. [ Links ]
Konjufca, V.K., Bottje, W.G., Bersi, T.K. & Erf, G.F., 2004. Influence of dietary vitamin E on phagocytic functions of macrophages in broilers. Poult. Sci. 83, 1530-1534. [ Links ]
López-Ferrer, S., Baucells, M.D., Barroeta, A.C., Blanch, A. & Grashorn, M.A., 1999. N-3 enrichment of chicken meat using fish oil: Alternative substitution with rapeseed and linseed oils. Poult. Sci. 78, 356-365. [ Links ]
Mahmoud, K.Z. & Edens, F.W., 2005. Influence of organic selenium on hsp 70 response of heat-stressed and enteropathogenic Escherichia coli-challenged broiler chickens (Gallus gallus). Comp. Biochem. Physiol. Part C 141, 69-75. [ Links ]
McCord, J.M. & Fridovich, J., 1969. Superoxide dismutase: An enzymatic function for Erythrocuprein. J. Biol. Chem. 244, 6049-6055. [ Links ]
Mcllroy, S.G., Goodall, E.A., Rice, D.A., McNulty, M.S. & Kennedy, D.G., 1993. Improved performance in commercial flocks with subclinical infectious bursal disease when fed diets containing increased concentrations of vitamin E. Avian Path. 22, 81-94. [ Links ]
Meluzzi, A., Sirri, F., Manfreda, G., Tallarico, N. & Franchini, A., 2000. Effects of dietary vitamin E on the quality of table eggs enriched with n-3 long-chain fatty acids. Poult. Sci. 79, 539-545. [ Links ]
Miret, S., Sáiz, M.P. & Mitjavilo, M.T., 2003. Effects of fish oil- and olive oil-rich diets on iron metabolism and oxidative stress in the rat. Br. J. Nutr. 89, 11-18. [ Links ]
Naumann, C. & Bassler, R., 1993. Die chemische Untersuchung von Futtermitteln VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs-und Forschungsanstalten)-Verlag, Darmstadt. [ Links ]
Naylor, A.J., Chost, M. & Jacques, K.A., 2000. Effects of selenium source and level on performance and meat quality in male broilers. Poult. Sci. 79 (Suppl.), 117. [ Links ]
NRC, 1994. Nutrient Requirements of Poultry, 9th ed., National Academic Science, Washington, D.C., USA. [ Links ]
Özkan, S., Basmacıoğlu-Malayoğlu, H., Yalçın, S., Karadaş, F., Koçtürk, S., Çabuk, M., Oktay, G., Özdemir, S., Özdemir, E. & Ergül, M., 2007. Dietary vitamin E (α-tocopherol acetate) and selenium supplementation from different sources: performance, ascites-related parameters and antioxidant status in broilers reared at low and optimum temperatures. Br. Poult. Sci. 48, 580-593. [ Links ]
Paglia, D.E. & Valentine, W.N., 1967. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Clin. Med. 70, 158-168. [ Links ]
Payne, R.L. & Southern, L.L., 2005. Comparison of inorganic and organic selenium sources of broilers. Poult. Sci. 84, 898-902. [ Links ]
Poirier, J., Cockell, K., Hidiroglou, N., Madere, R., Trick, K. & Kubow, S., 2002. The effects of vitamin E and selenium intake on oxidative stress and plasma lipids in hamsters fed fish oil. Lipids 37, 1125-1133. [ Links ]
Polavarapu, R., Spitz, D.R., Sim, J.E., Follansbec, M.H., Oberley, L.W., Rahemtulla, A. & Nanji, A.A., 1998. Increased lipid peroxidation and impaired antioxidant enzyme function is associated with pathological liver injury in experimental alcoholic liver disease in rats fed diets high in corn oil and fish oil. Hepatology 27, 1317-1323. [ Links ]
Qureshi, A.A., Mo, H., Packer, L. & Peterson, D.M., 2000. Isolation and identification of novel tocotrienols from rice bran with hypocholesterolemic, antioxidant, and antitumor properties. J. Agric. Food Chem. 48, 3130-3140. [ Links ]
Roch, G., Boulianne, M. & De Roth, L., 2000. Effects of dietary vitamin E and selenium source on incidence of ascites, growth performance and blood parameters cold-stressed broilers. Poult. Sci. 79 (Suppl. 1), 41. [ Links ]
Scheideler, S.E. & Froning, G.W., 1996. The combined influence o dietary flaxseed variety, level, form, and storage conditions on egg production and composition among vitamin E-supplemented hens. Poult. Sci. 75, 1221-1226. [ Links ]
Sen, C.K., Atalay, M., Ågren, J., Laaksonen, D.E., Ray, S. & Hönninen, O., 1997. Fish oil and vitamin E supplementation in oxidative stress at rest and after physical exercise. J. Appl. Physio , 83, 189-195. [ Links ]
Sijben, J.W.C., Schrama, J.W., Nieuwland, M.G.B., Hovenier, R., Beynen, A.C., Verstegen, M.W.A. & Parmentier, H.K., 2002. Interactions of dietary polyunsaturated fatty acids and vitamin E with regard to vitamin E status, fat composition and antibody responsiveness in layer hens. Br. Poult. Sci. 43, 297-305. [ Links ]
Simopoulos, A.P., 2000. Symposium: Role of poultry products in enriching the human diet with n-3 PUFA. Human requirements for n-3 polyunsaturated fatty acids. Poult. Sci. 79, 961-970. [ Links ]
Simopoulos, A.P. & Salem Jr, N., 1992. Egg yolk as a source of long-chain polyunsaturated fatty acids in infant feeding. Am. J. Cli. Nutr. 55, 411-414. [ Links ]
Sirri, F., Tallarico, N., Meluzzi, A. & Franchini, A., 2002. Long-chain n-3 PUFA enrichment of broiler chicken tissues using dehydrated algae Schizochytrium sp. Arch. Geflügelk. 66, 151 (Abstr. [ Links ]).
Sparks, N.H.C., 2006. The hen's egg-is its role in human nutrition changing? Wrld's Poult. Sci. 62, 308-315. [ Links ]
SPSS, 2001. Statistical software for windows version 11.0 Microsoft® [ Links ]
Stanley, G.V., Chakwu, H. & Thompson, D., 1998. Singly and combined effects of organic selenium (Se-Yeast) and vitamin E on ascites reduction in broilers. Proc. Poult. Sci. Association, Abstr. 111. [ Links ]
Surai, P.F., 2002a. Selenium in poultry nutrition. 1. Antioxidant properties, deficiency and toxicity. Wrld's Poult. Sci. J. 58, 333-347. [ Links ]
Surai, P.F., 2002b. Selenium. Natural Antioxidants in Avian Nutrition and Reproduction, Nottingham, Nottingham University Press, UK. [ Links ]
Surai, P.F., 2006. Selenium in Nutrition and Health, Nottingham University Press, Nottingham, United Kingdom. [ Links ]
Swain, B.K., Jahri, T.S. & Majumdar, S., 2000. Effects of supplementation of vitamin E, selenium and their different combinations on the performance and immune response of broilers. Br. Poult. Sci. 41, 287-292. [ Links ]
Tokuşoğlu, Ö. & Ünal, K., 2003. Biomass nutrient profiles of three microalgae: Spirulina Platensis, Chlorella Vulgaris and Isochrisis Galbara, L. Food Sci. 68, 144-1148. [ Links ]
# Corresponding author. E-mail: hatice.basmacioglu@ege.edu.tr


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}