










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.2 Pretoria Feb. 2009
SHORT COMMUNICATION
Investigation of effects of three candidate genes on leg action and fat deposition traits in pigs
M.L. MakgahlelaI; B. FanII, III, #; Z.-Q. DuII; M.F. RothschildII
IAgricultural Research Council, Private Bag X2, Irene 0062, South Africa
IIDepartment of Animal Science and Centre for Integrated Animal Genomics, Iowa State University, Ames, 50011, USA
IIIKey Laboratory of Agricultural Animal Genetics, Breeding and Reproduction, Ministry of Education & College of Animal Science & Technology, Huazhong Agricultural University, Wuhan, 430070, P.R. China
ABSTRACT
Data from 188 sows were used in the current study to examine the effects of high mobility group AT-hook1 (HMGA1), transcription factor 7-like-2 (TCF7L2) and insulin-like growth factor binding protein 3 (IGFBP3) genes on leg action and fat deposition traits, and further to explore the possible relationships between these genes on both traits. The candidate genes used in the study are known for their roles in fat deposition and growth. Overall leg action was scored on a scale of 1 (good movement) to 9 (leg weakness). Fatness traits included 10th rib backfat (BF10), adjusted 10th rib backfat to 125 kg (adjBF10) and last rib backfat (last BF), measured by ultrasonic imaging approach. The association analyses between single nucleotide polymorphisms (SNPs) and traits were performed using PROC MIXED procedures of SAS. The results showed that the associations between HMGA1, TCF7L2 and IGFBP3 genotypes with fat deposition traits were mostly suggestive in this limited data set. Leg action was also suggestively associated with IGFBP3 gene effects but was not associated with HMGA1 and TCF7L2 genes. Thus, IGFBP3 AA homozygote individuals tended to have had better movements (5.40), and were fatter when compared to GG homozygotes (5.84). The results from this study suggest a possible association between the IGFBP3 gene effects on both leg action and fatness. Therefore, further studies must be carried out in several populations, and using larger data to demonstrate these results conclusively.
Keywords: Candidate genes, overall leg action, fatness, association, pig
Leg and feet soundness play a major economic role in the commercial pig industry. Next to reproductive failure, feet and leg abnormalities are the most frequent reasons for culling of young sows (Boyle et al., 1998). Culling rates due to these problems accounted for 11% of the reasons for total disposal in Ireland, and range from 10 to 40% in Canada (Chagnon et al., 1991) and the USA (Irwin et al., 2000). In Denmark, up to 72% of culls were attributed to locomotive system problems (Kirk et al., 2005). Genetic improvement for leg action traits has in the past been challenging, since these traits have been shown to exhibit low to moderate heritability (Webb et al., 1983; Huang et al., 1995; Jorgensen & Andersen, 2000; Nikkilä et al., 2008). High fat deposition in pigs has been found to be associated with low lean meat production (Chen et al., 2002). Genetic progress for reduced fatness, using traditional breeding methods has been successful. Conversely, selection for reduced fatness has been shown to have antagonistic relationships with leg action traits (Huang et al., 1995; Stern et al., 1995; Jorgensen & Andersen, 2000; Nikkilä et al., 2008). Marker assisted selection (MAS) may be an effective strategy in improving these traits of economic importance and at the same time overcome pleoitropic effects in genes.
The current study was carried out to examine the effects of high mobility group AT-hook1 (HMGA1), transcription factor 7-like-2 (TCF7L2) and insulin-like growth factor binding protein 3 (IGFBP3) genes on leg action and fat deposition in pigs, and, furthermore, to explore the possible relationships between these genes on both traits. In previous studies, porcine HMGA1 and TCF7L2 genes had been shown to have associations with fat measurements, and were mapped on Sus scrofa chromosomes 7 and 14, respectively (Kim et al., 2004; Du et al., 2008). Insulin-like growth factor binding protein 3 is a member of the IGFBP family. Binding proteins prolong the half-life of insulin-like growth factors, and inhibit or stimulate the growth promoting effects of the IGFs on the cell culture (Duan, 2002; Mohan & Baylink, 2002; Mote, 2008).
A total of 188 commercial Large White and Landrace intercross females from Newsham Choice Genetics were used in this study. When the animals weighed about 125 kg, overall leg action was measured on a scale of 1 (excellent movement) to 9 (severe leg weakness) by an experienced scorer. Fatness traits including 10th rib backfat (BF10), adjusted 10th rib backfat (AdjBF10) and last rib backfat (Last BF) were measured using an ultrasonic imaging approach with a Pie Medical 200 (Classic Medical Supply Inc., Tequesta, FL). Standard protocols were followed to extract DNA from the sows using the DNeasy 96 Blood and Tissue kit (Qiagen, Valencia, CA, USA). The detailed information about single nucleotide polymorphisms (SNPs) and genotyping analyses is presented in Table 1. The association analyses between SNPs and traits were carried out using the PROC MIXED procedures of SAS (2006). Statistical models of analyses included genetic line, scoring date and genotype as fixed effects; sire within line as a random effect and body weight as a covariate.
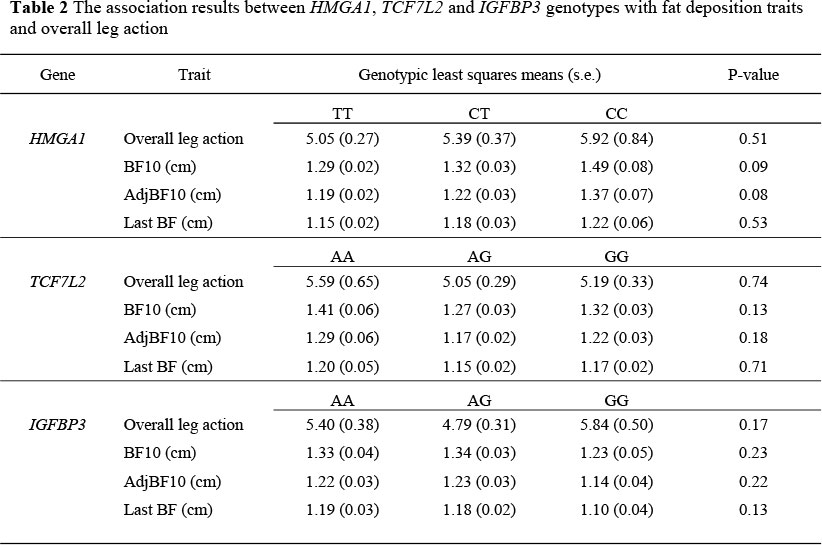
The association results between HMGA1, TCF7L2 and IGFBP3 genotypes with fat deposition traits and leg action score are presented in Table 2. Leg action was suggestively associated with the IGFBP3 gene (P <0.2), but was not associated with HMGA1 and TCF7L2 genes (P >0.2). On the basis of these results the association between overall leg action and IGFBP3 indicates that, in this population, AA homozygote individuals had better movements (5.40) compared to GG homozygotes (5.84). The association between the HMGA1 gene with BF10 and adjBF10 were suggestive (P <0.1), but was not indicative with Last BF (P >0.2). Thus, CC homozygote individuals deposited more fat compared to lean TT homozygotes. Kim et al. (2004) reported significant associations between this gene and backfat measurements in Landrace, Large White and Duroc x Large White populations. However, they found no associations between HMGA1 and backfat measurements in a Duroc population. These differences in gene markers and their effects on traits in different populations could be in part due to the genetic background and population sizes considered. The TCF7L2 gene also had a suggestive effect on BF10 and adjBF10 (P <0.2), but had no association with last BF (P >0.2). On the contrary, Du et al. (2008) found significant associations between TCF7L2 with last BF (P <0.05) and BF10 (P <0.01) in a Berkshire and Yorkshire intercross population.
The candidate genes HMGA1, TCF7L2 and IGFBP3 have been demonstrated in other pig populations for their transcriptional regulation roles in fat deposition and growth (Duan, 2002; Mohan & Baylink, 2002; Kim et al., 2004; Du et al., 2008; Mote. 2008). Observations in the field have long suggested that there may be unfavourable genetic correlations between fat deposition traits and overall leg action in pigs. In the present study the results from the association analyses between HMGA1, TCF7L2 and IGFBP3 genes were suggestive for most fat deposition traits. However, IGFBP3 was the only candidate gene found to have a suggestive effect on leg action (P <0.2), probably due to a limited dataset. On the basis of these findings, allele A in IGFBP3 is desirable for leg action but unfavourable for fatness. Thus, IGFBP3 transcription factors involved in the regulation of fat deposition may also function in the transcriptional activation of leg soundness. Although this association was indicative in this study, it is in agreement with previous findings that the genetic correlations between leg action and fat deposition measurements are antagonistic in pigs (Huang et al., 1995; Jorgensen & Andersen, 2000; Nikkilä et al., 2008). On the other hand, the association between the favourable alleles for both leg action and fatness in HMGA1 and TCF7L2 (T and G, respectively) suggests that for these genes less fatness may be associated with better leg soundness. These associations would be ideal in pig breeding since the objective is to breed pigs that deposit less fat and have good overall leg action.
Previously, HMGA1, TCF7L2 and IGFBP3 gene effects on leg action have not been reported for pigs. Results from this study suggest a possible association between the effects of these candidate genes and both leg action and fatness. Because of the limited sample size, further investigations are required in several populations, and using larger population size, to demonstrate these associations conclusively.
Acknowledgements
The authors would like to thank members of M.F. Rothschild's laboratory for their assistance in this research.
References
Boyle, L., Leonard, F.C., Lynch, B. & Brophy, P., 1998. Sow culling patterns and sow welfare. Irish Vet. J. 51, 354-357. [ Links ]
Chagnon, M., D'Allaire, S. & Drolet, R., 1991. A prospective study of sow mortality in breeding herds. Can. J. Vet. Res. 55, 180-184. [ Links ]
Chen, P., Baas, T.J., Mabry, J.W., Dekkers, J.C.M., & Koehler, K.J., 2002. Genetic parameters and trends for lean growth rate and its components in U.S. Yorkshire, Duroc, Hampshire, and Landrace pigs. J. Anim. Sci. 80, 2062-2070. [ Links ]
Du, Z.-Q., Fan, B., Zhao, X., Amoako, R. & Rothschild, M.F., 2008. Association analyses between type 2 diabetes genes and obesity traits in pigs. Obes. 17, 330-334. [ Links ]
Duan, C., 2002. Specifying the cellular response to IGF signals roles of IGF binding proteins. J. Endocrinol. 175, 41-54. [ Links ]
Huang, S.Y., Tsou, H.L., Kan, M.T., Lin, W.K. & Chi, C.S., 1995. Genetic study of leg weakness and its relationship with economic traits in central tested boars in subtropical area. Livest. Prod. Sci. 44, 53-59. [ Links ]
Irwin, C.K., Geiger, J.O., Pretzer, S.D. & Henry, S., 2000. Identifying causes of sow mortality rates in sow herds. Proc. of the 16th Congr. of the Inter. Pig Vet. Soc., 17-20 September, Melbourne, Australia, 294. [ Links ]
Jørgensen, B., & Andersen, S., 2000. Genetic parameters for osteochondrosis in Danish Landrace and Yorkshire boars and correlations with leg weakness and production traits. Anim. Sci. 71, 427-434. [ Links ]
Kim, K.S., Thomsen, H., Bastiaansen, J., Nguyen, N.T., Dekkers, J.C., Plastow, G.S. & Rothschild, M.F., 2004. Investigation of obesity candidate genes on porcine fat deposition quantitative trait loci regions. Obes. Res. 12, 1981-1994. [ Links ]
Kirk, R.K., Svensmark, B., Ellegaard, L.P. & Jensen, H.E., 2005. Locomotive disorders associated with sow mortality in Danish pig herds. J. Vet. Med. 52, 423-428. [ Links ]
Mohan, S. & Baylink, D.J., 2002. IGF-binding proteins are multifunctional and act via IGF dependent. J. Endocrinol. 175, 19-31. [ Links ]
Mote, B.E., 2008. The use of genetic markers to improve sow productive life and genetic abnormalities. PhD (Agric) thesis, Iowa State University, USA. [ Links ]
Nikkilä, M., Stalder, K., Mote, B., Lampe, J., Thorn, B., Rothschild, M.F., Johnson, A., Karriker, L. & Serenius, T., 2008. Heritabilities and genetic correlations of body composition and structural soundness traits in commercial gilts. Iowa State University Industry Report 2008. A.S. Leaflet R2349, available from http://www.ans.iastate.edu/report/air/ [ Links ]
SAS, 2006. Statistical Analysis Systems user's guide, Release 9.1. SAS Institute, Inc., Cary, North Carolina, USA. [ Links ]
Stern, S., Lundeheim, N., Johansson, K. & Andersson K., 1995. Osteochondrosis and leg weakness in pigs selected for lean tissue growth rate. Livest. Prod. Sci. 44, 45-52. [ Links ]
Webb, A.J., Russell, W.S. & Sales, D.I., 1983. Genetics of leg weakness in performance-tested boars. Anim. Prod. 36, 117-130. [ Links ]
# Corresponding author. E-mail: bfan@iastate.edu


{kind=link}
{kind=link}