










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.2 Pretoria Feb. 2009
Long-term selection experiment with Afrikaner cattle 1. Environmental factors affecting calf growth traits
L.M. BeffaI, II, 1; J.B. van WykI, #; G.J. ErasmusI
IUniversity of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa
IIMatopos Research Station, P.Bag K5137, Bulawayo, Zimbabwe
ABSTRACT
A selection and line x environment interaction study with grade Afrikaner cattle was established in 1956 at Matopos Research Station, Zimbabwe. Two selection lines of 100 cows each were reared in different management environments. The non-supplemented (NS) line relied on the range throughout the year and was mated to calve with the onset of the rains (December to February). The supplemented (S) line was offered protein-rich supplements during the dry season and mated to calve prior to the onset of the rains (October to December). In 1976 lines were sub-divided into four lines with 75 cows each, where one sub-line remained within each environment as a control; the remaining sub-lines were interchanged between environments. Bulls were selected on weaning weight within control lines, while replacement heifers were selected on weight at mating within sub-line. Data collected over a 40-year period (1958 to 1997) were analyzed. There were no line differences, however, productivity in the S environment was superior compared with the NS environment. While there were no environment differences in birth weight, calves born early in the S environment were estimated to be 90 kg (33%) heavier at 18 months than calves born late in the NS environment. The performance of calf growth from birth to 18 months in this study has emphasized the importance of matching animal physiological status with seasonal changes in the sub-tropics. The effect of previous lactation status (PLS) increased from 1.2 ± 0.13 kg at birth to 7 ± 0.6 kg at 205 days of age, a difference that persisted to 18 months (5 ± 0.9 kg). Given the significant and consistent detrimental effects of PLS on calf growth, it is recommended that this effect be considered in routine evaluations.
Keywords: Sanga, beef cattle, growth traits, lactation status, sub-tropics
Introduction
Until the mid 1970's the Afrikaner was the dominant breed in southern Africa and was used in the formation of composite breeds (Oliver, 1983). The introduction of the Brahman into Zimbabwe in the mid 1950's led to the marginalization of the Afrikaner and currently there are no Afrikaner stud breeders. The Afrikaner remains an important breed in South Africa, being promoted as a dam line for crossbreeding purposes. Recent DNA studies showed that southern African Sanga breeds of cattle are largely Bos taurus, now classified as Bos taurus africanus (Mostert et al., 1998). The influence of non-genetic factors on the growth of range beef calves has been very well documented for Bos taurus and B. indicus cattle in temperate and tropical environments, however, there is little information for Sanga cattle in the sub-tropics (Moyo, 1990; Van der Westhuizen, 1997).
Mixed model methodologies (Henderson, 1984), incorporating the numerator relationship matrix (NRM), facilitate the use of genetic linkages across contemporary groups yielding more accurate measures of both non-genetic factors as well as improved genetic parameter estimates. The inclusion of the NRM also allows for the accounting of genetic trends (Blair & Pollak, 1984). However, these complex mathematical models are at best a crude attempt to model complicated biological events that are subject to numerous non-systematic interacting factors.
In addition to the standard factors typically influencing calf growth (such as herd-year-season, age of dam, sex and age of calf), the long calving season in this trial poses additional modelling challenges. In contrast to reports in the temperate regions, reports in the sub-tropics and tropics have noted the significant influence of the previous lactation of the cow on calf growth (Hetzel et al., 1989; Moyo, 1990). The objective of this study was to quantify the nature of non-genetic factors influencing calf growth in Afrikaner cattle in a sub-tropical environment from birth to 18 months of age.
Materials and Methods
Matopos Research Station, southwest Zimbabwe, is characterised by wide fluctuations in the quantity and distribution of rainfall within and across season (Ward et al., 1979). Rainfall normally occurs between November and March and is followed by a long dry season. For the 40-year period of the study (1957 to 1997) the mean annual rainfall was 566 mm. The vegetation on the predominantly red soils is dominated by Acacia species and the dominant grass species are Heteropogon contortus, Themeda triandra and Cymbopogon plurinodes (Rattray, 1957).
From a common gene pool, two lines of 100 grade Afrikaner cows each were established and subjected to different nutritional and management regimes, hereafter termed environments. The supplemented (S) line cows were offered supplements during the dry season (9 kg maize silage and 1 kg cottonseed meal/head/day) and were mated to calve early (October to December) relative to the expected onset of the rains. The non-supplemented (NS) line cows were mated to calve two months later than those in the supplemented line. While supplementation afforded a relatively 'early' mating and calving season, the effects of supplementation and calving season are totally confounded. As a result, routine practices such as castration, weaning, dosing and vaccinations were performed at fixed times in the year where calf age differed by approximately two months.
For the first eight years (1957 to 1964) six foundation sires were rotated among the lines, thereafter the lines have been closed. However, the rotation was not uniform: three bulls were used for three to six years in the S environment, and for only one year each in the NS environment; similarly two bulls were used for three to four years in the NS environment but only one year each in the S environment. Only one bull was used for more than one year in both environments. A fixed mating season of 90 days and four single-sired herds of 25 cows each were used within each line. Three bulls were replaced each year and one was retained as a repeat sire. Selection for weight within line to establish lines adapted to their respective environments was based on weight at weaning for bulls and weight prior to mating (three years) for replacement heifers.
From 1976 to 1978 the number of breeding females in each line was increased to 150 and two equal sub-lines were created. One sub-line remained within each environment as a control; the remaining sub-lines were interchanged between environments. Significantly, the supplementation regime in the supplemented environment was changed so that animals were offered 0.5 kg of protein-rich concentrates/head/day during the dry season. Bulls were selected within the control sub-lines and were used within lines across environments. Cows were mated in five single-sire herds of 15 cows each. Four bulls were replaced each year and one was retained as a repeat sire.
Bull selection took place two weeks before weaning and in the post-crossover phase, six animals were selected with a proviso that not more than two animals would be selected from a particular cowherd. Candidates had to have reasonably well developed testes and sound animals were selected on weight linearly adjusted for age. From 1988, selection criteria was based on weight adjusted for other known environmental factors, derived from a fixed effects model, which were line, environment, year of birth and age and previous lactation status of the dam (heifer, suckling or not suckling). Replacement heifers were generated within sub-lines and entered their respective mating groups at 27 months of age. Replacement rates were kept constant across sub-lines and were dictated by the sub-line with the least number of suitable (>265 kg) replacement heifers. Thus selection and culling intensities differed for the sub-lines. Cows were culled for poor production (infertility and low calf weaning weight). In the post crossover phase cows in excess of 12 years of age were generally not retained for breeding purposes. The allocation of sires and heifers to the mating herds was based on minimizing relationships as far as possible. It is important to note that after the crossover was implemented, only half the genetic material, i.e. sires, continued to be crossed over environments, while selection of heifer replacements catered for any within sub-line adaptation.
The experimental site was subdivided into 35 paddocks and as far as possible lines were offered similar grazing. As cows displayed signs of approaching parturition they were moved to a 'calving' paddock close to the homestead, which was checked daily and cow-calf pairs were rounded-up and taken to the handling facilities within 24 h of birth. Calves were identified by means of ear notching and weights were recorded. All cattle were dipped in an acaricide plunge-dip once a week in the wet season and once every fortnight at the height of the dry-season, June through to August. All animals were annually vaccinated against rabies, anthrax, botulism and young animals were vaccinated against quarter-evil and contagious abortion (heifers). It was also recommended that young animals be treated for round worms and fluke at the beginning and end of the wet season (in October and in May). As a standard procedure cattle were individually weighed on a monthly basis. All male calves not retained for breeding purposes were castrated approximately two weeks before physical weaning in mid-August where calf ages ranged from four to seven months for the NS environment and from seven to 10 months for the S environment.
Birth days (BD) were transformed to a standard calving date within year relative to a base date of 1st September. As complete sets of dates for the bulling period in the pre-crossover phase were not available, the calving distributions within environments were scrutinized for obvious 'out of season' calves. This was based on the assumption that the bulling seasons were approximately 90 days and ranged from mid-December to mid-March and mid-February to mid-May for the S and NS environments respectively, together with a gestation interval ranging from 280 to 300 days. The calving pattern in the S environment indicated that the mating season extended beyond mid-March; there were, however, no obvious breaks in the distribution to assist in distinguishing 'out of season' calvings. Fourteen out of a total of 3775 calvings were deemed out of season and discarded. Complete records detailing the start and end of the mating seasons were available for the post-crossover phase, and assuming a gestation interval of 280 to 300 days, 53 records out of a total of 4552 were deemed out of season and discarded. In all instances, a cow was credited with calving in the season in question for early calves and was credited for calving in the next season for late born calves.
Weights at birth (BW), weaning (WW, recorded in July), yearling (W12, recorded in October) and 18 months (W18, recorded in May) were analyzed. In addition, calf weight at 205 days of age (W205), derived from monthly weighings, was analyzed, bearing in mind that calves in the S and NS environments would obtain these ages at very different stages of the grazing season. Post-weaning, the weights in October and in May were extracted to define the start and end of the second grazing season. These weights approximated yearling and 18 month weights. Pre-weaning gain (ADG) was measured between birth and 205 days of age, while post-weaning gain (PWG) taken as the gain between YW and W18. Growth traits were also scrutinized within year and mating season for obvious outliers. Pre-weaning weights and weight gains for five calves in the late-mated group were discarded where gains were less than 3 kg/month and in most instances the calf died before weaning. The post-weaning growth patterns of six calves in the post-crossover phase were exceptionally poor resulting in the discarding of three yearling and three 18-month weights.
In preliminary analyses, year of birth was the most important factor accounting for the greatest proportion of total variation in the growth traits. However, significant two- and three-way interactions were expressed with most other fixed effects considered. Furthermore, in terms of the accounting for variation in the observed values, the interaction between environment and year of birth was the most important. As it was neither practical nor feasible to account for all interactions with year of birth, a concatenation of line-year-environment (LYE) classification was employed. For all the traits, only two classes of sex were considered. Pre-weaning, these were intact male versus female and post-weaning, these were steer versus heifer. Data of bull calves post-weaning were excluded from analyses.
In the pre-crossover phase, age of dam ranged from four to 17 years, apart for 1975 and 1976 matings in the S line where heifers were mated to calve as three-year-olds. However, the number of observations for cows older than 12 years was less than 4% for number of births. In the post-crossover phase, age of dam ranged from three to 14 years and less than 2% of the observations were of cows older than 12 years of age. While preliminary analyses revealed that age of dam (in years) included both as linear and quadratic covariates or as a class model accounted for a similar proportion of the total variance in the observed values, the covariate did not account for the confounding with cow lactation status of heifers in the previous season. Furthermore, previous lactation status (PLS) tended to interact with age of dam, the effect of PLS being greatest for young (<5 years) and old (>10 years) cows. In order to account for the interaction as well as confounding of heifer with PLS status, a class model concatenating three classes for PLS (0 for three year old heifers; 1 for cows not suckling in the previous season; and 2 for cows that had suckled a calf at least up to and until the calf had attained 90 days of age) with five age of dam classes (3, 4, 5 to 7, 8 to 10 and >10 years of age) was employed. Age of dam classes were derived after perusal of the least squares means for the various traits, where it was found that the performance of cows between five and seven and between eight and 10 years of age were similar.
With a long calving season extending 150 days across the two environments (October to February), it was not surprising to find important effects due to BD in preliminary analyses. Given the vagaries of the rainfall season and therefore grazing it was deemed inappropriate to account for month of birth as a cross-classified effect. A model where month of birth was nested within year would be partially confounded with environment. It was also not possible to treat BD as a simple covariate due to the partial confounding with the environment as well as significant interactions with year of birth. Within year and environment, the covariate of BD ranged over 100 days. The inclusion of BD as a covariate within environment and a split in calving season (early versus late), in attempt to avoid influence of outliers, did not indicate an improved model fit and notably complicated the estimation of environment effects. As the environment effect was accounted for by fitting a line-year of birth-environment (LYE) class model, the effects of BD were accounted for by its inclusion as a cross-classified covariate regressed from the year-environment subclass means, where it was assumed that BD did not interact with other fixed effects, in particular with year and environment. Calf age for weights taken at a fixed time in the year (WW, W12 and W18) and birth day was totally confounded. However, for W205, age deviated from the target date, was included in the analyses.
The following fixed effects model was employed for the analyses of fixed effects of growth traits:
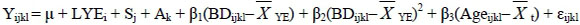
where Yijkl is the vector of observations; µ is the overall mean; LYEi is the effect of ith line-year-environment class (i = 1 to 122) for all traits except W18 and PWG (i = 1 to 116); Sj is the effect of jth sex class (j = 1 to 2); Ak is the effect of kth age-of-dam - previous lactation status class (k = 1 to 9); β1, β2 and β3 are the regression coefficients of the covariable on the observation; (BDijkl - 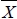 YE) and (BDijkl -
YE) and (BDijkl - 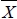 YE)2 are the linear and quadratic effect of the birth day deviated from the year-environment subclass mean; (Ageijkl -
YE)2 are the linear and quadratic effect of the birth day deviated from the year-environment subclass mean; (Ageijkl - 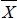 t) is the linear effect of calf age deviated from target date for W205 only and εijkl is the error term. Mixed model procedures (MTDFREML; Boldman et al., 1995), allowing for the inclusion of direct and maternal genetic random effects, as well as their correlation, and permanent environmental random effects due to repeated measures on the dam, were used to derive best linear unbiased estimates of fixed effects, regression coefficients and specific contrasts.
t) is the linear effect of calf age deviated from target date for W205 only and εijkl is the error term. Mixed model procedures (MTDFREML; Boldman et al., 1995), allowing for the inclusion of direct and maternal genetic random effects, as well as their correlation, and permanent environmental random effects due to repeated measures on the dam, were used to derive best linear unbiased estimates of fixed effects, regression coefficients and specific contrasts.
Results and Discussion
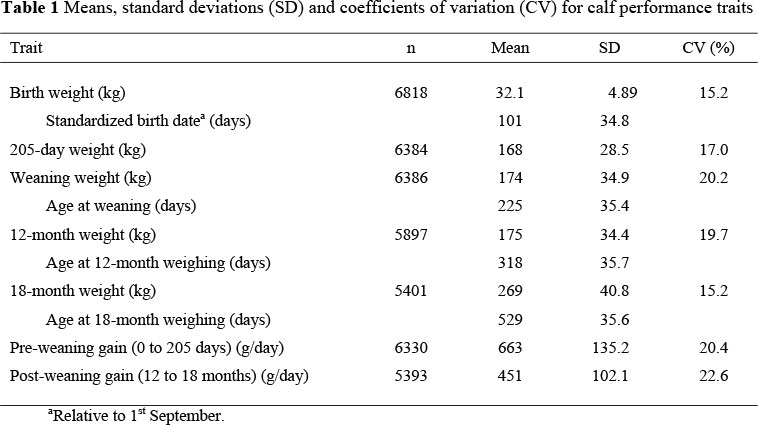
The number of observations, means, standard deviations and coefficients of variation (CV) of the weights and weight gains after editing are presented in Table 1. The CV for BW and W205 were between 15 and 17%, but increased to 20% for WW and W12, presumably reflecting the age differences between environments. Despite these differences, the CV for W18 had reverted back to 15%.
The similarity in production performance of the Afrikaner in this experiment and across a wide range of environments is remarkable (Botswana: Trial et al., 1977; Zimbabwe: Moyo, 1990; South Africa: Mostert et al., 1998), suggesting that the Afrikaner could be referred to as a stable genotype, as oppossed to a plastic genotype where the degree of phenotypic expression varies under different environmental conditions (Bryant et al., 2005).
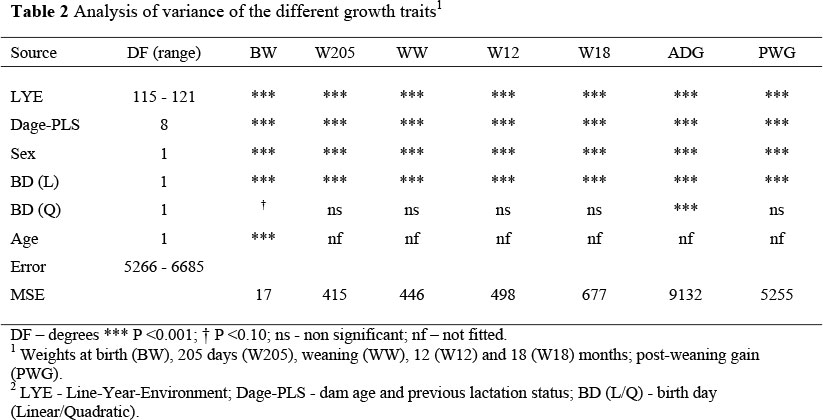
The total variation in the observed values accounted for by the fixed effect models for the growth traits (Table 2) tended to increase with age and varied from 30 to 64%. The effect of LYE was the most important effect accounting for 14 to 44% of the total variation and between 38 and 86% of variation accounted for by the models. The effect is illustrated in the yearly trends for WW (Fig. 1) showing the tremendous year-to-year variations as well as the large environmental effect. The large year-to-year variations are consistent with findings from other studies in the region (Rege & Moyo, 1993; Van der Westhuizen, 1997). Calves from the S line had small and consistently greater (P >0.05) weights and weight gains than their NS counterparts.
There was no apparent environmental effect for BW; however, calves born in the S environment had a distinct growth advantage to calves born in the NS environment that increased with age. The effect was marked for weight gains between 150 and 205 days where weight gains of calves in the S environment were 212 g/day greater (P <0.05) than those of their NS contemporaries (data not shown). On average, calves in the NS environment would attain 150 days of age in June, which is well into the dry season. At this time, grazing quality and quantity would be limiting and would presumably have a marked impact on the dam's ability to nurse its young calf. Perusal of the line means reveal markedly different response of calves to environment and is illustrated for WW in Figure 1. At weaning, calves from the S environment were 34 kg heavier (Figure 1), a difference that persisted to 18 months. Part of this difference is also due to the difference in age between environments of 53 days.
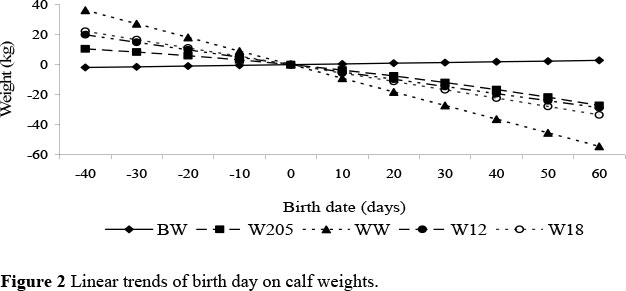
With a calving season extending over 100 days within environment it was not surprising to note the dramatic influence of birth day on growth traits. The effects of BD (linear for all traits and quadratic covariates for W205 and ADG only) accounted for 5% of the total variation for BW, which increased to 10% for WW and persisted to 18 months accounting for 7% of the total variation for W18. Similarly, BD accounted for 7% for ADG, before declining to 1% for PWG. Late born calves were on average 4 kg heavier at birth than early born calves (Fig. 2). The increase in birth weight as the season progresses has been attributed to the improved grazing conditions with the onset of the rains (Carvalheira et al., 1995). The effect of birth day on weaning weight was dramatic with early born calves being up to 90 kg heavier than their late born contemporaries. In a study of Mashona cattle, Buvanendran (1990) also found that later born calves were significantly lighter (1 to 9%) than early born calves. Similarly, Niemann & Heydenrych (1965) found that weight at weaning decreased by 2.86 kg/week for calves born after 1st October. Birth day did not appear to influence PWG, but the effect of BD persisted up to 18 months and was of the order of 56 kg (Figure 2). Rege & Moyo (1993) noted that the effect of delayed BD of calf was more apparent at older ages and was attributed to a declining maternal influence. The analysis within line indicated that the covariate of BD interacted with environment for BW and PWG, but the effect of the covariate was remarkably constant across environment for all the other growth traits.
The combined effects of environment and BD imply that performance of calves born late in the NS environment are at a great disadvantage compared with calves born early in the S environment; a disadvantage that persists at least until the end of the second grazing season and is estimated to be in the order of 90 kg for W18. The persistent effect of late calving on growth post-weaning has been found in numerous other studies (Buvanendran, 1990). However, among Nguni calves in the lowveld, Tawonezvi et al. (1986) reported that late born calves grew faster and maintained that advantage to 18 months of age. This differential response was ascribed to the different production environments.
The effect of sex was fairly consistent and accounted for 3 to 6% of the total variation. Males (intact pre-weaning and steers post-weaning) were consistently 7% heavier (P <0.01) from birth through to 18 months. Similar effects due to sex of calf have been reported with Sanga cattle (Tawonezvi, 1989; Buvanendren, 1990; Carvalheira et al., 1995; Van der Westhuizen, 1997), in contrast to larger effects (10 to 13%) shown for Bos taurus cattle breeds in temperate regions (Nelsen & Kress, 1981).
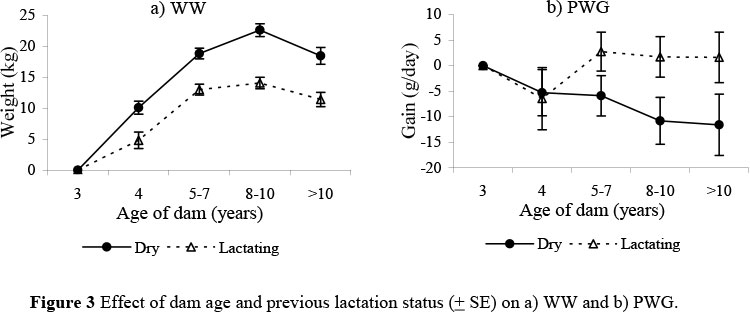
The effect of age of dam and previous lactation status tended to decrease as calves grew older and accounted for 5% of total variation in pre-weaning traits, but only 0.2% for PWG. The effect of PLS increased from 1.2 ± 0.13 kg at birth to 7 ± 0.6 kg at 205 days of age, a difference that persisted to 18 months (5 ± 0.9 kg). The effect of PLS was important for ADG, but was small and reversed for PWG. The consistent effect of age of dam and previous lactation status on weights from birth to 18 months is well illustrated in Figure 3a. Weights increased with age of dam and peaked for 8 to 10 year-old dams before decreasing for older dams, while the effect of PLS tended to be least for calves from 5 to 7 year-old dams. However, compensatory gains were indicated for PWG (Figure 3b).
Analysis within environment did not reveal evidence to support the assumption that cows in the NS environment were under greater stress to provide milk for their calves (data not shown). Similar effects due to previous lactation have been reported where the larger effect found in young cows was attributed to the extra nutrient requirements for growth (Thorpe et al., 1980; Tawonezvi et al., 1986; Hetzel et al., 1989; Tawonezvi, 1989). Hetzel et al. (1990) found that the effects of PLS were highest in Hereford-Shorthorn, intermediate in Afrikaner cross and lowest in Brahman cross, with the authors concluding that the magnitude of maternal dam age and PLS effects were inversely proportional to the level of tropical adaptation of the genotypes. The persistent effects of both age and previous lactation status of dam post-weaning concurs with some studies (Tawonezvi et al., 1986; MacKinnon et al., 1990; Moyo, 1990; Rege & Moyo, 1993), others show compensatory gains (Tawonezvi, 1989) and Hetzel et al. (1990) found no carry-over effects. In contrast, in an experiment with Afrikaner calves in Mozambique, Carvalheira et al. (1995) only found an effect of PLS at birth.
Conclusions
The average reproduction and growth performance of the Afrikaner across a wide range of production environments show remarkable similarities which is probably related to many generations of selection under harsh environmental conditions. The effect of month of calving on growth was dramatic with late born (February) calves displaying markedly poorer growth compared with their early born (October) contemporaries. It is therefore essential to appreciate the dramatic differences in production environment both within and across seasons in the sub-tropics, determined by the quantity and distribution of rainfall, and emphasizes the importance of matching animal physiological status with seasonal changes in the sub-tropics (De Waal, 1990). The effects of PLS need to be considered to avoid penalizing calves from more productive cows.
Acknowledgements
This study formed part of the first author's doctorate thesis who benefited from funding from the John Wakeford Scholarship Trust, the Afrikaner Cattle Society of South Africa, the International Livestock Research Institute the South African National Research Foundation and the Livestock Identification Trust. This work was funded by and carried out at Matopos Research Station, Ministry of Agriculture, Zimbabwe. Important contributions by numerous individuals, both named and unmentioned, influenced the conduct of this long term experiment. They include T.S. Clark, B.P. Kinghorn, A.S. Pretorius, J.D.G. Steenkamp, T.H. Vorster and H.K. Ward. The dedicated care of animals and meticulous data collection over the years by staff of Matopos Research Station and in particular of Lucydale Section is gratefully acknowledged. The first author also acknowledges with gratitude a three-month study period under the guidance of L.R. Baker in Nairobi, Kenya (ILRI).
References
Blair, H.T. & Pollak, E.J., 1984. Estimation of genetic trend in a selected population with and without the use of a control population. J. Anim. Sci. 58, 878-886. [ Links ]
Boldman, K.G., Kriese, L.A., Van Vleck, L.D., Van Tassell, C.P. & Kachman, S.D., 1995. A Manual for Use of MTDFREML. A Set of Programs to Obtain Estimates of Variances and Covariances [DRAFT]. [ Links ] U.S. Department of Agriculture, Agriculture Research Service.
Bryant, J., López-Villalobos, N., Holmes, C. & Pryce, J., 2005. Simulation modeling of dairy cattle performance based on knowledge of genotype, environment and genotype by environment interactions: current status. Agric. Syst. 86, 121-143. [ Links ]
Buvanendran, V., 1990. Adjustment factors for weaning weights of Mashona cattle in Zimbabwe. J. Agric. Sci., Camb. 114, 35-40. [ Links ]
Carvalheira, J.G.V., Blake, R.W., Pollak, E.J. & Van Soest, P.J., 1995. Comparison of Landim and Africander cattle in southern Mozambique: I. Body weights and growth. J. Anim. Sci. 73, 3519-3526. [ Links ]
De Waal, H.O., 1990. Animal production from native pasture (veld) in the Free State Region - A perspective of the grazing ruminant. (Review). S. Afr. J. Anim. Sci. 20, 1-9. [ Links ]
Henderson, C.R., 1984. Applications of Linear Models in Animal Breeding. University of Guelph, Ontario, Canada. [ Links ]
Hetzel, D.J.S., Mackinnon, M.J., Dixon, R. & Entwistle, K.W., 1989. Fertility in a tropical beef herd divergently selected for pregnancy rate. Anim. Prod. 49, 73-81. [ Links ]
Hetzel, D.J.S., Quaas, R.L., Seifert, G.W., Bean, K.G., Burrow, H.M. & Aspden, W.J., 1990. Genetic parameters for growth of tropical beef cattle. Proc. Aust. Assoc. Anim. Breed. Genet. 8, 517-520. [ Links ]
Mackinnon, M.J., Hetzel, D.J.S., Corbet, N.J., Bryan, R.P. & Dixon, R., 1990. Correlated responses to selection for cow fertility in a tropical beef herd. Anim. Prod. 50, 417-424. [ Links ]
Mostert, B.E., Groeneveld, E., Rust, T. & Van der Westhuizen, J., 1998. Multitrait variance component estimation of South African beef breeds for growth traits. Proc. 6th World Congr. Genet. Appl. Livest. Prod. 23, 145-148. [ Links ]
Moyo, S., 1990. Evaluation of the productivity of indigenous cattle and some exotic beef breeds and their crosses in Zimbabwe. International Livestock Centre for Africa, Addis Ababa, Ethiopia. [ Links ]
Nelsen, T.C. & Kress, D.D., 1981. Additive and multiplicative correction factors for sex and age of dam in beef cattle weaning weight. J. Anim. Sci. 52, 1217-1224. [ Links ]
Niemann, P.J. & Heydenrych, H.J., 1965. Some environmental and physiological factors affecting birth and weaning weights of beef calves. Tech. Comm. No. 41. Dept. of Agricultural Technical Services. Government Printer, Pretoria. (Cited by De Waal, 1990). [ Links ]
Oliver, J., 1983. Beef cattle in Zimbabwe, 1890-1981. (Review). Zimbabwe J. Agric. Res. 21, 1-17. [ Links ]
Rattray, J.M., 1957. The grasses and grass associations of southern Rhodesia. Rhod. Agric. J. 54, 197-234. [ Links ]
Rege, J.E.O. & Moyo, S., 1993. Effect of birth date on growth performance of calves of seasonally-bred range cattle. J. Anim. Breed. Genet. 110, 209-227. [ Links ]
Tawonezvi, H.P.R., 1989. Growth of Mashona cattle on range in Zimbabwe. I. Environmental influences on liveweight and weight gain. Trop. Anim. Hlth. Prod. 21, 37-42. [ Links ]
Tawonezvi, H.P.R., Brownlee, J.W.I. & Ward, H.K., 1986. Studies on growth of Nkone cattle. 1. Environmental influences on body mass. Zimbabwe J. Agric. Res. 24, 17-29. [ Links ]
Thorpe, W., Cruickshank, D.K.R. & Thompson, R., 1980. Genetic and environmental influences on beef cattle production in Zambia. 1. Factors affecting weaner production from Angoni, Barotse and Boran dams. Anim. Prod. 30, 217-234. [ Links ]
Trail, J.C.M., Buck, N.G., Light, D., Rennie, T.W., Rutherford, A., Miller, M., Pratchett, D. & Capper, B.S., 1977. Productivity of Africander, Tswana, Tuli and crossbred beef cattle in Botswana. Anim. Prod. 24, 57-62. [ Links ]
Van der Westhuizen, J., 1997. Performance factors influencing total herd efficiency in beef cattle. Ph.D. dissertation, University of the Orange Free State, Bloemfontein, South Africa. [ Links ]
Ward, H.K., Richardson, F.D., Denny, R.P. & Dye, P.T., 1979. Matopos Research Station: A perspective. Rhod. Agric. J. 76, 5-18. [ Links ]
# Corresponding author. E-mail: vanwykjb.sci@ufs.ac.za
1Current address. Livestock Identification Trust, P O Box BE 209, Belvedere, Harare, Zimbabwe


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}