










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.1 Pretoria Jan. 2009
SHORT COMMUNICATION
Single nucleotide polymorphisms in the 5'-flanking region of the prolactin gene and the association with reproduction traits in geese
R.S. Jiang; L.L. Zhang; Z.Y. Geng; T. Yang; S.S. Zhang
College of Animal Science and Technology, Anhui Agricultural University, Hefei, 230036, P.R. China
ABSTRACT
Prolactin (PRL), a polypeptide hormone synthesized and secreted by the animal's anterior pituitary gland, plays an important role in the regulation of mammalian lactation and avian reproduction. Considering the significant association between single nucleotide polymorphisms (SNPs) in the 5'-flanking region of PRL and reproduction traits in the chicken, the objective of this study was to screen for SNP in the 5'-proximal region of PRL in geese and to evaluate the association between SNP and reproduction traits in geese. Chinese Wan-xi White geese (n = 200) and European Rhine geese (n = 80) were used for phenotyping. SNP was screened by comparing sequences of PCR products, and the single-stranded conformational polymorphism (SSCP) protocol was adopted for genotyping. Three SNPs (A-401G, G-268A and T-266A) in the 5'-proximal region of goose PRL were identified in both breeds. Statistical analysis suggested that the genotype AA characterized by A-401, G-268 and T-266 had a positive genetic effect on egg production. Therefore, these polymorphisms have the potential to be utilized in molecular breeding for egg production in geese.
Keywords: Goose, Wan-xi White, Rhine, PRL, egg production, SNP
Prolactin (PRL) is one of the polypeptide hormones synthesized and secreted by the animal's anterior pituitary gland. It has been demonstrated that PRL is an essential hormone regulating reproduction and lactation in mammals (Byrnes & Bridges, 2005; Bonomo et al., 2007). In domestic chickens and turkeys, PRL has proven to be one of the key hormones in the onset and maintenance of broodiness (Sharp et al., 1988). A sharp rise in plasma PRL level may induce incubation behaviour and thus terminate laying (Sockman et al., 2000), resulting in decreased egg production (Reddy et al., 2002). Shiue et al. (2006) showed that mRNA expression levels of PRL were significantly higher in high than in low egg production strains, indicating that egg production was related to a PRL mRNA expression in chickens. In waterfowl species such as ducks and geese, a sharp rise in plasma PRL levels occurred during the formation of the final 10 - 20% of the clutch, during which females markedly increased their nest-box occupancy (Hall, 1987; 1991; Fang et al., 2005). Plasma PRL levels of non-laying ducks were significantly lower than those of laying ducks (Bluhm et al., 1983). Therefore, PRL exhibits similar changing profiles in the reproduction cycle and exerts biological functions in the similar pattern in different domestic avian species. To elucidate the genetics of PRL, genomic structures of PRL in avians have been investigated extensively (Zhou et al., 2001; Kansaku et al., 2005). An abundance of SNP has been reported in the 5'-flanking region of chicken PRL (Liang et al., 2006), and the 24-bp Insert-deletion was significantly associated with broody behaviour and egg production (Jiang et al., 2005; Cui et al., 2006), indicating its usefulness as a molecular marker for egg production. However, whether SNPs exist and are associated with goose reproduction as exhibited in chickens, is still unknown. The purpose of the present study was to identify and characterize the SNP in the 5'-flanking region of PRL and evaluate the genetic effects of the SNP on reproduction traits in geese, thus providing a molecular reference in marker-assisted selection programmes.
The stocks used in the experiment were unrelated female Wan-xi White geese (n = 200), a local Chinese breed from the Anhui Province and unrelated female Rhine geese (n = 80) that have been imported to China previously. Up to 25 wk of age the geese were reared on a floor system in open-sided houses. At 25 wk of age the birds were transferred to pens (3 x 3 m) in groups of four per pen. Each pen contained on one side a 3 x 3 m outside ground area with a water bath. During the laying period the experimental geese were fed a commercial maize-soyabean-based diet containing 155 g CP/kg feed with a metabolisable energy content of 10.87 MJ/kg feed, and sufficient forage. The birds had free access to feed and water.
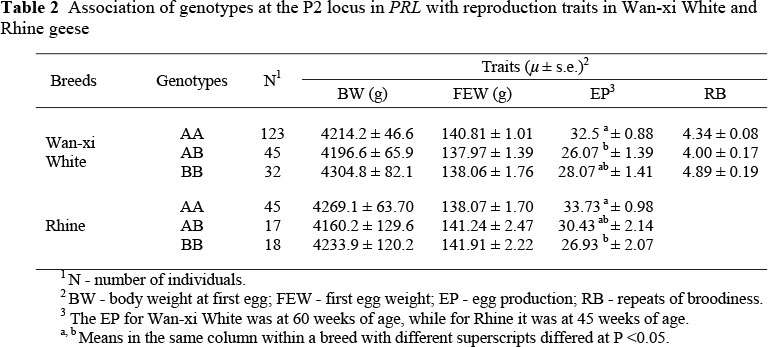
Reproductive traits recorded in the experiment included body weight at first egg (BW), the first egg weight (FEW) and total egg production (EP). The EP for Wan-xi White was measured up to 60 wks of age, i.e. the total first laying cycle, while for Rhine it was up to 45 wks of age, the early stage of the first laying cycle. To accurately record individual EP, each pen was equipped with four separated nest-boxes that prevented escaping. The four geese in a pen were each put into a nest-box for the night and were taken out in the morning after checking whether they had laid or not.
The repeats of broodiness (RB) which were defined as the number of broody cycles of the Wan-xi White, were recorded during the total first laying cycle. A goose was defined as broody when she persistently nested for more than three consecutive days, exhibited nesting defence, and reduced food intake and activities. The criterion for the end of broodiness was that the bird did not exhibit broody behaviour for more than one week.
Genomic DNA was isolated from geese blood samples by the phenol-chloroform method (Sambrook et al., 1989). PCR was performed to amplify the fragments of the 5'-proximal region of PRL. The primers (P1) were designed according to the sequence of goose PRL (GenBank accession No.: EF190486) using the online primer design procedure, Primer 3.0 (Primer 3.0, Whitehead Institute for Biomedical Research, www.cbr.nrc.ca/cgi-bin/primer3_www.cgi). The sequences of P1 were: Forward 5'- TGGACATGC ACATCTTTTACG -3', Reverse 5'- ATCCACAAGTGAATGGCTAA -3'. The 20 µL PCR volume included 50 ng of DNA template, 0.20 mM deoxynucleoside triphosphate, 2.5 mM MgCl2, 0.20 mM primer and 0.5 U of Taq DNA polymerase (Dingguo Biotechnology Company, Beijing, P. R. China). The PCR protocol was 94 °C for 5 min followed by 35 cycles of 94 °C for 1 min, 55 °C for 1 min, and 72 °C for 1 min and a final extension at 72 °C for 10 min. The PCR products were purified using a DNA Fragment Quick purification/Recover Kit (Dingguo Biotechnology Company, Beijing, P. R. China), ligated to the pMD 18-T vector (Dingguo Biotechnology Company, Beijing, P. R. China) and transformed into DH5-α Escherichia coli (Dingguo Biotechnology Company, Beijing, P. R. China) for cloning (Sambrook et al., 1989). Sequencing was performed on an ABI377 sequencer.
PCR products of 20 individuals were individually sequenced to screen the SNP. Because of variation, sites screened in this study only ranged from -402 to -255 bp. To facilitate genotyping, another PCR set of primers (P2) that produced a shorter PCR product (<300 bp) was re-designed to amplify the region covering the variation sites. The P2 was as following: Forward 5'-AAGCCCCCATTATCCCTCTC -3', Reverse 5'-TGGTTTGATGAAGAGAATGCC -3'. The PCR products of P2 for all individuals were genotyped by single-stranded conformational polymorphism protocol: 2 µL of PCR product of each individual were mixed with 5 µL of denaturing buffer (98% formamide, 0.09% xylene cyanole FF, and 0.09% bromophenol blue) and then denatured at 94 °C for 5 min followed by a rapid chill on ice for 10 min. The denatured PCR products were electrophoresed for 14 h at 8 V/cm on 12% acrylamide gels. The DNA bands on the gel were stained with 0.2% AgNO3 for 20 min, followed by 3% Na2CO3 for about 5 min (Qu et al., 2005). Genotypes were recorded according to banding patterns. The PCR products of P2 for each homozygote were sequenced to identify the SNP mutation type.
Body weight, FEW and EP were calculated for each genotype. Genotypic effects on BW, FEW, EP and RB were analyzed by one-way ANOVA using the GLM procedure of the SAS Institute (2001) with genotype (G) as the fixed effect, according to the following model:
Y = µ+ G + e
where Y = dependent variable, µ = population mean and e = random error. Significant differences among means of different genotypes or haplotypes were calculated using the Duncan's multiple-range test, and the significance was determined at P <0.05.
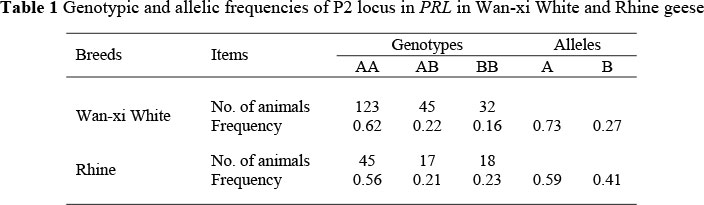
The sequence fragment, from -790 to -1, of 5'-proximal region of goose PRL was successfully amplified using the primer P1. By comparing the sequences of 20 individuals, three nucleotide variation sites, A-401G, G-268A and T-266A, were detected. By developing the single-stranded conformational polymorphism protocol, three genotypes were identified at the P2 locus both in Wan-xi White and Rhine geese and defined as AA, AB, and BB (Figure 1). Sequencing of the different genotypes showed that the homozygote individuals of genotype AA shared the genomic characteristics of -401A, -268G and -266T, and the BB of -401G, -268A and -266A, while the heterozygote AB combined the two.
Genotypic and allelic frequencies of the P2 locus were calculated within each breed (Table 1). In Wan-xi White and Rhine geese, the genotypic frequency for AA (0.62 and 0.56, respectively) was the greatest and, therefore, the allele A was predominant in the populations (0.73 and 0.59, respectively).
As shown in Table 2, no significant difference (P >0.05) was found between genotypes in BW and FEW in the two breeds, and the RB did not differ (P >0.05) between genotypes in Wan-xi White. However, in Wan-xi White geese, the mean EP in the first laying cycle of genotype AA (32.5) was greater (P <0.05) than that of genotype AB (26.07), and in Rhine geese, the mean EP at 45 wk of age of genotype AA (33.73) was greater (P <0.05) than that of genotype BB (26.93).
By screening the 5'-flanking region of goose PRL, three SNPs, A-401G, G-268A and T-266A, were identified both in Wan-xi White, a Chinese breed and in Rhine geese, a European breed, suggesting the abundance of nucleotide variations in the region as exhibited in the chicken. The fact that the three SNPs produced only three genotypes in the experimental populations indicated the linkage heredity of the SNPs. In the study the association of genotypes with egg production in Wan-xi White and Rhine geese shared a similar profile, the geese of genotype AA laid more eggs than genotypes AB or BB, indicating that allele A could be the favourite for egg production or linked with the QTLs of interest. The similar association observed in both breeds with different genetic backgrounds basically ensured the reliability of the result, suggesting that the enrichment of allele A in a population by molecular selection may be helpful for the enhancement of egg production in geese.
The plasma PRL protein expression level is of great importance in regulating avian reproduction ability including incubation and egg production. In avian species, changes in plasma PRL level are associated with the expression of PRL mRNA in the anterior pituitary (Wong et al., 1991). However, the expression of PRL is regulated by the 5'-flanking region sequence by binding with specific transcription factors (Jiang et al., 2005). Studies in mammals and birds have shown that Pit-1/GHF-1 (Frisch et al., 2000), oestrogen receptors (Maurer & Notides, 1987), the CCAAT-enhancer binding protein-α (Enwright et al., 2003) and other proteins are essential in regulating the expression of PRL via specific promoter binding sites. The sequence variation in the 5'-proximal region of PRL may lead to changes of transcription factor binding sites and alter the expression of PRL. In the experiment reported here, there were three single nucleotide variations at -401, -268 and -266 bp sites in the 5'-proximal region of goose PRL. To understand whether and how the SNPs change the transcription factor binding sites, binding modes for pre- and post-mutation were predicted using TFSEARCH ver.1.3 (http://www.cbrc.jp/research/DSB/tfsearch.html). According to the prediction, the binding sites for transcription factor GATA-2 and GATA-1 are lost when the G-268A and T-266A mutation occurs. Although the transcription factors GATA-1 and GATA-2 play key roles in the regulation of gene mRNA expression (Ohneda & Yamamoto, 2002), whether and how the nucleotide variations identified in the present study lead to the alteration of transcription factor binding mode and mRNA expression of PRL in geese are still to be tested in future studies.
In conclusion, three SNPs in the 5'-proximal region of goose PRL were identified. Statistical analysis suggested that the genotype AA characterized by A-401, G-268 and T-266 had positive genetic effects on egg production (P <0.05), and therefore the polymorphisms reported here have the potential for utilization in molecular marker-assisted selection programmes for egg production in geese.
Acknowledgments
The authors are grateful to the members of the poultry laboratory of the Anhui Agricultural University for phenotyping and data collecting. This work was supported by a grant from the National Natural Science Foundation of China (30600431) and the Bureau of Science and Technology of Anhui Province (06013039A).
References
Bluhm, C.K., Phillips, R.E. & Burke, W.H., 1983. Serum levels of luteinizing hormone (LH), prolactin, estradiol, and progesterone in laying and nonlaying canvasback ducks (Aythya valisineria). Gen. Comp. Endocrinol. 52, 1-16. [ Links ]
Bonomo, I.T., Lisboa, P.C., Pereira, A.R., Passos, M.C. & de Moura, E.G., 2007. Prolactin inhibition in dams during lactation programs for overweight and leptin resistance in adult offspring. J. Endocrinol. 192, 339-344. [ Links ]
Byrnes, E.M. & Bridges, R.S., 2005. Lactation reduces prolactin levels in reproductively experienced female rats. Horm.Behav. 48, 278-282. [ Links ]
Cui, J.X., Du, H.L., Liang, Y., Deng, X.M., Li, N. & Zhang, X.Q., 2006. Association of polymorphisms in the promoter region of chicken prolactin with egg production. Poult. Sci. 85, 26-31. [ Links ]
Enwright, J.F., Kawecki-Crook, M.A. & Voss, T.C., 2003. A PIT-1 homeodomain mutant blocks the intranuclear recruitment of the CCAAT/enhancer binding protein a required for prolactin gene transcription. Mol. Endocrinol. 17, 209-222. [ Links ]
Fang, D.A., Geng, Z.Y., Zhang, X.R. & Tao, Y., 2005. Studies on the role of hypothalamic dopamine and 5-hydroxy tryptamine in modulation broodiness in Wanxi goose. China Anim. Husb. Vet. Med. 32 (7), 35-37. [ Links ]
Frisch, H., Kim, C., Hausler, G. & Pfaffle, R., 2000. Combined pituitary hormone deficiency and pituitary hypoplasia due to a mutation of the Pit-1 gene. Clin. Endocrinol. 52, 661-665. [ Links ]
Hall, M.R., 1987. External stimuli affecting incubation behavior and prolactin secretion in the duck (Anas platyrhynchos). Horm. Behav. 21, 269-287. [ Links ]
Hall, M.R., 1991. Endocrinological and behavioral changes associated with the onset of incubation in the duck. Physiol. Behav. 50, 311-316. [ Links ]
Jiang, R.S., Xu, G.Y., Zhang, X.Q. & Yang, N., 2005. Association of polymorphisms for prolactin and prolactin receptor genes with broody traits in chickens. Poult. Sci. 84, 839-845. [ Links ]
Kansaku, N., Ohkubo, T., Okabayashi, H., Guémené, D., Kuhnlein, U., Zadworny, D. & Shimada, K., 2005. Cloning of duck PRL cDNA and genomic DNA. Gen. Comp. Endocrinol. 141, 39-47. [ Links ]
Liang, Y., Cui, J., Yang, G., Leung, F.C. & Zhang, X., 2006. Polymorphisms of 5'-flanking region of chicken prolactin gene. Domest. Anim. Endocrinol. 30, 1-16. [ Links ]
Maurer, R.A. & Notides, A.C., 1987. Identification of an estrogen responsive element from the 5'-flanking region of the rat prolactin gene. Mol. Cell. Biol. 7, 4247-4254. [ Links ]
Ohneda, K. & Yamamoto, M., 2002. Roles of hemotopoietic transcription factors GATA-1 and GATA-2 in the development of red blood cell lineage. Acta Haematol. 108, 237-245. [ Links ]
Qu, L.J., Li, X.Y., Wu, G.Q. & Yang, N., 2005. Efficient and sensitive method of DNA silver staining in polyacrylamide gel. Electrophoresis 26, 99-101. [ Links ]
Reddy, I.J., David, C.G., Sarma, P.V. & Singh, K., 2002. The possible role of prolactin in laying performance and steroid hormone secretion in domestic hen (Gallus domesticus). Gen. Comp. Endocrinol. 127, 249-255. [ Links ]
Sambrook, J., Fritsch, E.F. & Maniatis, T., 1989. Molecular Cloning: A Laboratory Manual. 2nd ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., USA. [ Links ]
SAS, 2001. Statistical Analysis Systems user's guide (Version 8.2.). SAS Institute Inc., Raleigh, North Carolina, USA. [ Links ]
Sharp, P.J., Macnamee. M.C., Sterling, R.J., Lea, R.W. & Pedersen, H.C., 1988. Relationship between prolactin, LH and broody behavior in bantam hens. J. Endocrinol. 118, 279-286. [ Links ]
Shiue, Y.L., Chen, L.R., Chen, C.F., Chen, Y.L., Ju, J.P., Chao, C.H., Lin, Y.P., Kuo, Y.M., Tang, P.C. & Lee, Y.P., 2006. Identification of transcripts related to high egg production in the chicken hypothalamus and pituitary gland. Theriogenology 66, 1274-1283. [ Links ]
Sockman, K.W., Schwabl, H. & Sharp, P.J., 2000. The role of prolactin in the regulation of clutch size and onset of incubation behavior in the American kestrel. Horm. Behav. 38, 168-176. [ Links ]
Wong, E.A., Ferrin, N.H. & Silsby, J.L., 1991. Cloning of a turkey prolactin cDNA: Expression of prolactin mRNA throughout the reproductive cycle of the domestic turkey (Meleagris gallopavo). Gen. Comp. Endocrinol. 83, 18-26. [ Links ]
Zhou, M., Zhang, X.Q., Shi, Z.D. & Cao, Y.C., 2001. Cloning and sequencing of prolactin gene cDNA in three chicken breeds. Yi Chuan Xue Bao. 28, 614-620. [ Links ]
 Correspondence:
Correspondence:
E-mail: jiangrunshen@ahau.edu.cn


{kind=link}
{kind=link}
{kind=link}