










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Animal Science
On-line version ISSN 2221-4062
Print version ISSN 0375-1589
S. Afr. j. anim. sci. vol.39 n.1 Pretoria Jan. 2009
SHORT COMMUNICATION
Genetic parameter estimates for functional herd life for the South African Jersey breed using a multiple trait linear model
J. du ToitI, II; J.B. van WykII; A. MaiwasheIII
IAgricultural Research Council, Animal Production Institute, Private Bag X5013, Stellenbosch 7599, South Africa
IIDepartment of Animal, Wildlife and Grassland Sciences, University of the Free State, P.O. Box 339, Bloemfontein 9300, South Africa
IIIAgricultural Research Council, Animal Production Institute, Private Bag X2, Irene 0062, South Africa
ABSTRACT
Longevity reflects the ability of a cow to avoid being culled for low production, low fertility or illness. Longevity can be used in breeding programmes if genetic parameters are known. Various measures are used for longevity. In this study survival in each of the first three lactations was analysed. Survival was denoted by a 1 if a cow survived, and 0 otherwise. The primary objective of the current study was to estimate genetic parameters for functional herd life. The secondary objective was to compare estimates of genetic parameters from the linear sire and animal models. Data and pedigree records on purebred Jersey cows that participated in National Milk Recording and Improvement Scheme of South Africa were used to estimate genetic parameters. A total of 181 269 cow records from 636 herds recorded over 16 years were available for analysis. Estimates of genetic parameters for herd life were obtained using REML procedures fitting three-trait (first three lactations as separate traits) linear animal and sire models. Heritability estimates (0.02 to 0.03) from the animal and sire models were somewhat similar for all lactations. However, heritability estimates for lactations 2 and 3 were slightly higher with the sire model compared to the animal model. The genetic correlation between lactations 1 and 2 from both the sire and animal models was higher than that between lactations 2 and 3. Genetic correlations from the sire model ranged from 0.68 to 0.99, compared to 0.76 to 0.99 from the animal model. Results from the current study suggest that genetic variation exists for functional herd life to allow for genetic improvement through selection. The moderate positive genetic correlation between survival in the first and third lactation suggest that early selection for functional herd life is feasible.
Keywords: Functional herd life, linear model, genetic parameters
In South Africa, as in most other countries, dairy cattle breeding programmes were mainly focused towards yield traits. In the last decade interest has increased in broadening the breeding objective to include other traits such as fertility, health and longevity (Miglior, 2005; Sewalem et al., 2008).
Longevity or herd life in dairy cattle, a measure of success of the cow to delay the risk of involuntary culling, is a trait of considerable economic importance. Increased herd life is associated with decreased culling and therefore with decreased costs for raising or purchasing replacement heifers (Boettcher et al., 1999). In addition, increased herd life is associated with increases in mean production of the herd because a greater proportion of the culling decisions are based on production and less on diseases, resulting in a higher proportion of mature cows in the population.
Longevity can be measured in different ways and genetic evaluations are not standardized across countries. In most countries taking part in the international bull evaluation service (INTERBULL), survival analyses are being used to estimate direct longevity (Ducrocq & Sölkner, 1998). In Australia and Canada survival in each of the first three lactations is recorded as a binomial trait and evaluated with a multi-trait linear animal model in which survival in each lactation is considered a different trait (Madgwick & Goddard, 1989; Jairath et al., 1998). Despite these differences all models of evaluation aim to assist in selection of the same aggregate genotype of lifetime profitability. Many studies have incorporated adjustments for production in models for genetic analysis of herd life, and is termed functional herd life (Ducrocq et al., 1988; Dekkers et al., 1994). Culling decisions are made within herds and therefore the correction for production has to be on a within-herd basis (Vollema & Groen, 1996).
Currently there is no genetic evaluation for herd life of dairy cattle in South Africa. A prerequisite to the implementation of a genetic improvement programme is the estimate of genetic parameters for traits to be improved. The purpose of the current study was to estimate genetic parameters for functional herd life for the South African Jersey breed using a multiple trait linear approach.
Data and pedigree records on purebred South African Jersey cows that participated in the National Milk Recording and Improvement Scheme were obtained from the Integrated Registration and Genetic Information System (INTERGIS). The original data included test-day and lactation yields from 252 629 Jersey cows born between 1968 and 2005 from 2 028 herds sired by 5 394 sires. There were 4 189 393 test-day records in the original data file. Preliminary edits to ensure quality of records were conducted on the data. The editing criteria employed in the national evaluation for milk production traits for the Jersey breed were used since functional herd life is considered in the current study: For example, the following records were excluded from the analysis: (1) test-day milk yield < 1 kg or > 70 kg, fat yield < 2% or > 9%, and protein yield < 2% or > 6%, (2) first test exceeding 75 days, (3) at least one interval between test dates exceeding 100 days, and (4) records with more than one test date interval between 60 and 100 days.
The following further edits were employed: (1) first lactation records terminated before 01 January 1989 were excluded because records prior to this date comprised only completed lactations without test-day records, (2) only lactations less or equal to 305 days were considered, (3) records with incorrect herd code, yields equal to zero and records out of specified age range were excluded; the allowable age ranges were as follows: 17 to 40, 29 to 53, and 41 to 67 months for first, second, and third calving, respectively, (4) records with unknown registration status were excluded, and (5) a first parity record was required for all cows. Furthermore, records from cows with unknown sires were excluded. Cows born after 2004 were excluded due to limited number of records. A summary of the records excluded due to specific criterion is provided in Table 1. After editing, 181 269 cow records from 636 herds recorded over 16 years were available for analysis.
The first three lactation records on cows that calved from 1 January 1989 to 31 December 2004 were used for parameter estimation. Observations for longevity were created as follows: Survival to the next lactation was determined based on the presence or absence of a subsequent lactation. Survival in a given lactation was treated as a binary trait and coded 1 if the cow survived during that lactation and 0 if the cow was culled during that lactation or the number of days between the current calving and extraction date exceeded 581 days. Records from cows where the number of days between current calving date and data extraction date were less than 581 days were considered as records in progress and were excluded from the analysis. The value 581 was calculated as mean calving interval plus 3 standard deviations to ensure each cow had enough opportunity to calve if she was still in the herd. The criteria used to determine survival is further explained in Table 2. Survival rate per lactation is summarized in Table 2. Survival rate was 72.4, 71.6, and 67.4% for lactations 1, 2 and 3, respectively.
Two mixed linear models were used to estimate (co)variance components for survival: the three-trait sire and animal models. The three-trait sire model can be presented as follows:
Yijkl = hyil + rhsjl + (agek)l + (protk x rhsjl)jl + (protk)1 + (fatk)l + sirekl + eijkl [1]
where:
Yijkl = observation for survival (0 or 1) in lactation l (l = 1, 2 and 3) on cow k that calved in herd-year i; hyil = fixed effect of herd-year i for lactation l;
rhsjl = fixed effect of subclass j for registry status x herd size change x season of calving for lactation l;
(agek)l = linear and quadratic regressions of survival in lactation l on age at first calving (17 to 40 months);
(protkxrhsjl)jl = linear regression of survival in lactation l on protkl x rhsjl subclasses, where protk is the normalized deviation of 305-day protein yield within herd-year average in first lactation;
(protk)l + (fatk)l = linear, quadratic, and cubic regressions of survival in lactation l on normalized deviation of 305-day protein and fat yield within herd-year average in first lactation;
sirekl = random genetic effect of sire k;
eijkl = random residual.
The following distributional assumptions were made about the random effects: s~N(0,Aσ2s) and e~N(0,Iσ2e) where s is a vector of sire additive genetic effects and e is a vector of residual effects. The σ2s and σ2e are the sire and residual variances, respectively. The A is Wright's numerator relationship matrix and I is an identity matrix. The animal model was similar to equation [1] except that the sire genetic effect was replaced by the animal genetic effect. The animal genetic effects were assumed to be normally distributed with (co)variance structure Aσ2a with σ2a being the animal genetic variance.
A minimum of 10 cows sired by at least two sires were required per herd-year of calving to ensure connectedness in the data and to reduce the bias caused by preferential treatment of daughters of a particular sire in a particular herd or group of herds. Records of cows from herd-year subclass without variation were excluded from the analysis. Registration status was defined in two classes; namely, cows in herds with registered cows, and cows in herds with non-registered cows. Three classes for change in herd size were defined based on the percentage of change from one year to the next (decreasing = for a decrease in herd size of <-5%; nearly unchanged = no appreciable change >-5% to <10%; and increasing = for increasing in herd size of >10%).
The original data set including all the cows was used when creating the registration status and herd size change classes. Similar to Mostert et al. (2004), two seasons of calving were defined as Winter (season 1, from April to September) versus Summer (season 2, from October to March). The same procedure is used in the current national evaluation of the Jersey breed. The first lactation yield for protein and fat was used because the genetic correlation between first lactation yield and yield in later lactations is high. Furthermore, low yield in the terminal lactation could have been caused by a health problem, which is what functional herd life attempts to measure (Short & Lawlor, 1992).
(Co) variance components were estimated fitting a multiple-trait sire and animal model using the Variance Component Estimation (VCE 5) package of Groeneveld (2004). Heritability for the sire model was calculated using  . The standard errors of the heritability estimates were computed using the delta method of Searle et al. (1992).
. The standard errors of the heritability estimates were computed using the delta method of Searle et al. (1992).
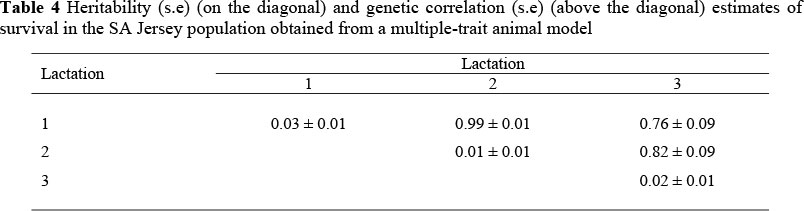
Estimates of heritability and genetic correlations for survival in the first three lactations from the multiple-trait sire and animal models are presented in Tables 3 and 4, respectively. Heritability estimates from both models were approximately similar for all lactations. When traits with low heritability such as longevity are analyzed with an animal model, most information comes from the sire component and the difference between sire and animal models is expected to be small (Vollema & Groen, 1996). Estimates of heritability were in the range of 0.02 to 0.03, which are comparable but on the lower end of previously published estimates of approximately 0.04 (Short & Lawlor, 1992; Jairath & Dekkers, 1994). Boettcher et al. (1999) reported heritabilities of approximately 0.07 for all lactations on the underlying scale with the threshold model and in the order of 0.08 with the survival analyses.
The genetic correlations between lactations 1 and 2 from both the sire and animal models were higher than that between lactations 2 and 3. The lowest correlation was between lactation 1 and 3 for both models. Boettcher et al. (1999) reported similar results using data on Canadian Holsteins. These results suggest that the genetic factors that determine survival are slightly different in the first and the third lactations. Genetic correlations from the sire model ranged from 0.68 to 0.99. This range is similar to results (0.84 to 0.91) reported by Boettcher et al. (1999) and Short & Lawlor (1992). Jairath & Dekkers (1994) observed genetic correlations that were moderately high (0.60 to 0.75), while Visscher & Goddard (1995) reported estimates between 0.91 and 0.97 for Holstein-Friesians and Jerseys.
In the current study a linear model was considered because all records prior to 2001 were officially terminated at 305 days and no opportunity for extended lactations existed. This practice caused a skewed distribution in lactation length. Secondly, a linear model is easy to implement and previous research has shown that the difference in heritability estimates expressed on the underlying scale is negligible when linear and non-linear models are used for analysis of binary traits (e.g. Boettcher et al., 1999).
Results from the current study suggest that genetic variation exist for functional herd life to allow for genetic improvement through selection, although the response to selection could be slow due to the low estimates of heritability. The moderate positive genetic correlation between survival in the first and third lactation suggest that early selection for functional herd life is feasible.
References
Boettcher, P.J., Jairath, L.K. & Dekkers, J.C.M., 1999. Comparison of methods for genetic evaluation of sires for survival of their daughters in the first three lactations. J. Dairy Sci. 82, 1034-1044. [ Links ]
Dekkers, J.C.M., Jairath, L.K. & Lawrence, B.H., 1994. Relationships between sire genetic evaluations for conformation and functional herd life of daughters. J. Dairy Sci. 77, 844-854. [ Links ]
Ducrocq, V. & Sölkner, J., 1998. "The Survival Kit"- V3.0, a package for large analyses of survival data. In: 6th World Cong. Genet. Appl. Livest. Prod., Volume 27, 447-448. Anim. Genetics and Breeding Unit, Univ. of New England, Armidale, Australia. [ Links ]
Ducrocq, V., Quaas, R.L., Pollak, E.J. & Casella, G., 1988. Length of productive life of dairy cows. I. Justification of a Weibull model. J. Dairy Sci. 71, 3061-3070. [ Links ]
Jairath, L.K. & Dekkers, J.C.M., 1994. Genetic parameters of functional and true lactational survival, and relationships with milk production and conformation traits in registered Canadian Holsteins. J. Dairy Sci. 77 (Suppl. 1), 146. (Abstract). [ Links ]
Jairath, L.K., Dekkers, J.C.M., Schaeffer, R.L., Liu, Z., Burnside, E.B. & Kolstad, B., 1998. Genetic evaluation for herd life in Canada. J. Dairy Sci. 81, 550-562. [ Links ]
Madgwick, P.A. & Goddard, M.E., 1989. Genetic and phenotypic parameters of longevity in Australian dairy cattle. J. Dairy Sci. 72, 2624-2632. [ Links ]
Miglior, F., 2005. Overview of different breeding objectives in various countries. Proc. 11th WHFF Meeting Session 4, 7-11. Guelph, Ontario Canada. [ Links ]
Mostert, B.E., Groeneveld, E. & Kanfer, F.H.J., 2004. Test day models for production traits in dairy cattle. S. Afr. J. Anim. Sci. 34, 35-37. [ Links ]
Sewalem, A., Miglior, F., Kistemaker, G.J., Sullivan, P. & Van Doormaal, B.J., 2008. Relationship between reproduction traits and functional longevity in Canadian dairy cattle. J. Dairy Sci. 91, 1660-1668. [ Links ]
Short, T.H. & Lawlor, T.J., 1992. Genetic parameters of conformation traits, milk yield and herd life in Holsteins. J. Dairy Sci. 75, 1987-1998. [ Links ]
Searle, S.R., Casella, G. & McCulloch, C.E., 1992. Variance Components. John Wiley & Sons, Inc., New York, N.Y., USA. [ Links ]
Tsuruta, S., Misztal, I. & Lawlor, T.J., 2005. Changing definition of productive life in US Holsteins: Effect on genetic correlations. J. Dairy Sci. 88, 1156-1165. [ Links ]
Visscher, P.M. & Goddard, M.E., 1995. Genetic parameters for milk yield, survival, workability and type traits for Australian dairy cattle. J. Dairy Sci. 78, 205-220. [ Links ]
Vollema, A.R. & Groen, A.F., 1996. Genetic parameters of longevity traits of an upgrading population of dairy cattle. J. Dairy Sci. 79, 2261-2267. [ Links ]
 Correspondence:
Correspondence:
Email: dtoitj@arc.agric.za


{kind=link}
{kind=link}
{kind=link}
{kind=link}